
太湖流域稻田管理方式对蜘蛛群落特征的影响*
2021-09-10 08:18满吉勇陈宝雄王子睿刘云慧
中国生态农业学报(中英文) 2021年9期
满吉勇, 袁 凯, 陈宝雄, 王子睿, 刘云慧
(1.中国农业大学资源与环境学院 北京 100193; 2.中国农业大学生物多样性与有机农业北京市重点实验室 北京 100193;3.中国农业大学有机循环研究院(苏州) 苏州 215000; 4.农业农村部农业生态与资源保护总站 北京 100125)
太湖流域自然条件优越, 经济基础雄厚, 人口密集, 自古以来便是我国社会发展最迅速的地区之一[1]。但长期以来, 因受高强度人为干扰活动的影响, 生态环境整体脆弱[2-3], 尤其以农业面源污染为代表的生态问题突出[4-5], 改善当地生态环境已迫在眉睫[4]。水稻(Oryza sativa)作为当地重要的种植作物, 改进稻田农业生产方式已成为当地生态环境改善的必由之路。生物多样性是生态安全和生态系统健康的重要表征, 不同的生产管理模式因施肥、除虫、除草等措施的不同, 可能会对稻田生物多样性产生不同的影响[6-7]。依赖化肥和农药投入来保障粮食产量的集约化稻田生产方式, 会导致农田适宜栖息地的丧失,造成稻田生物多样性下降, 甚至引起某些天敌物种的消失[8], 无法有效维持稻田害虫生物控制能力[9-10]。而有机、绿色等生态友好型的生产方式对稻田天敌的干扰相对较少, 往往可以为天敌提供更多的栖息地[11-13], 从而有利于天敌多样性的维持。此外, 不同种植季及不同稻区水稻田间主要害虫与天敌种类和消长规律存在差异[14], 这也可能会影响稻田生态系统对于管理方式的响应。
蜘蛛类群多样、对环境变化敏感, 常被作为环境变化、人类活动影响以及生物多样性和生态系统服务功能评估的重要指示生物[6]。同时, 蜘蛛无论在数量上还是功能方面均是稻田重要的天敌类群[15-16],在农田生态系统中提供了重要的害虫控制服务[17], 其多样性的提升不仅标志良好的农田生态环境质量,也有利于农田的可持续生产。同时, 除了多样性的变化, 群落功能特征及功能群变化也是反映土地利用影响和生态系统服务变化的重要指标[18-19]。目前,大部分研究集中在有机和常规生产对蜘蛛多样性影响的比较上[20-22], 却很少有人研究亚洲地区特有的绿色生产方式对蜘蛛多样性的影响[23], 且只考虑管理措施对蜘蛛多度和种类影响的研究较多[20-22], 而考虑其是如何影响蜘蛛关键种数量及其行为特征的研究较少。此外, 不同水稻生育期稻田害虫的数量不同, 可能改变食物资源的数量及质量, 进而自下而上调控蜘蛛群落结构对稻田管理的响应模式, 不同水稻生育期蜘蛛丰富度、多度以及功能性状的变化仍需要进一步的研究[24-25]。
因此, 本研究通过选取有机、绿色和常规3种不同管理模式的水稻农场开展蜘蛛多样性调查, 通过分析不同生产模式下蜘蛛群落的生物多样性及其物种组成和功能特征的差异, 评估不同生产管理方式下稻田蜘蛛多样性的状况, 为该地区制定推动生态环境质量改善、生态系统服务的生产管理策略和促进可持续发展提供科学依据。研究主要回答以下问题: 有机、绿色和常规3种稻田生产管理模式下稻田蜘蛛群落多样性、功能组成及群落结构是否存在差异? 哪种生产管理方式有利于蜘蛛多样性维持和害虫控制功能发挥?
1 研究区概况与研究方法
1.1 研究区概况
1.2 试验设计
2019年6−10月, 在该镇选取有机生产、绿色生产和常规生产的水稻农场进行蜘蛛多样性监测, 3个农场分别位于江苏省苏州市临湖镇中国农业大学水稻生态农场临湖基地(31.12°N, 120.45°E), 某农发集团绿色生产水稻农场(31.09°N, 120.28°E), 某农户常规生产水稻农场(31.08°N, 120.26°E), 每种生产方式选取5块重复样地(考虑到取样地的独立性, 样地间距离>500 m, 且选取样地距离稻田边缘>50 m, 并被其他稻田所包围), 共计15个样地, 每个样地为20 m×30 m的固定样方, 在每个样方内, 取中线划分成4个10 m×15 m的方块, 在样方和4个小方块的中央位置设置取样点(为避免边缘效应, 取样点距离样方边界>2 m), 每样点6丛, 固定每样地30丛稻株开展蜘蛛调查。不同农场基本管理方式如表1所示。
1.3 取样方法
考虑到节肢动物活动性较强, 采用吸虫器法取样, 利用吸虫器在稻株上中下部位抽吸, 吸虫器由意大利欧码BV300手提式吹风机改造而成, 功率为1 kW, 风量为720 m3∙h−1, 风速为70 m∙s−1, 在吸虫器的吸口端反套上一个长25 cm、宽15 cm的40目尼龙网袋, 用于蜘蛛虫样的收集, 尽可能吸完能看到的物种为止。为了解稻田蜘蛛多样性随水稻发育时期的变化, 并捕获尽可能多的蜘蛛样本, 于水稻分蘖早期(6月27日)、分蘖后期(7月17日)、拔节期(8月8日)、抽穗期(8月30日)、扬花期(9月14日)、灌浆期(9月26日)、黄熟期(10月15日)取样, 每次取样为期1 d, 并将蜘蛛放置于75%的乙醇溶液中保存。蜘蛛标本统计个数, 成虫鉴定至物种水平。
1.4 数据处理
多样性指数主要进行丰富度、多度、Simpson指数的计算[26], 蜘蛛群落的物种丰富度是同一样地5个样点捕获蜘蛛种类数量, 多度为同一样地5个样点捕获蜘蛛的总个体数。Simpson指数(D)的计算公式如下:
式中:ni为第i个物种的个体数,N为所有物种个体数,S为物种数。
为分析不同生产管理处理蜘蛛的多样性差异,首先对蜘蛛的物种丰富度及多度进行了正态性及方差齐性检验[27-28], 当数据分析满足正态性及方差齐性要求时, 采用单因子方差分析, 检验不同类型稻田的生物多样性指数的差异显著性[29]; 当数据不满足其中任意一个前置要求时, 采用非参数K-W检验方法检验不同类型稻田的生物多样性指数的差异显著性[30]。如检验发现不同组别存在显著差异, 则分别采用TukeyHSD法[31]与基于Bonferroni校正的多重比较方法[32]进行多重比较。
为分析不同农田处理蜘蛛物种组成的差异, 采用基于CNESS (normalized expected species shared)相似系数的非度量多维尺度法(non-metric multi-dimensional scaling, NMDS)比较分析不同生产模式稻田蜘蛛群落结构的相似性, NMDS中取样大小参数m可以变化, 但必须大于最小的取样值, 且小于最大的取样值,m值取值较大时, 强调稀有物种, 反之强调的则是优势物种[33-34]; 采用胁强系数(stress)检验拟合结果, 当模型stress值小于0.2时, 则差异性在可接受范围内, 说明此方法可以较好地反映群落组成的相对关系[35]。为进一步检验物种组成差异的显著性, 本研究采用非参数多元方差分析(PERMANOVA)分析物种组成是否存在显著差异, 当P<0.05时, 认为不同组别蜘蛛群落物种组成存在显著差异[36]。
为进一步分析不同管理方式稻田中蜘蛛在捕食及活动等功能行为特征和害虫控制能力上是否存在显著差异, 研究根据蜘蛛体长、捕猎方式和是否具备飞航能力将蜘蛛分为不同的功能类群(表2), 其中蜘蛛体长及捕猎方式通过检索世界蜘蛛名录网站(https://wsc.nmbe.ch/)和各地动物志[37-39]的方式获取,蜘蛛飞航能力通过相关文献获取[40-44]。蜘蛛的体长体现蜘蛛的资源利用效率与蜘蛛群落内部的捕食情况[45]; 捕猎方式体现蜘蛛的捕食策略及其捕捉的猎物种类[46]; 飞航能力体现蜘蛛在碎片化的景观中进行长距离迁移的能力[40]。计算群落功能加权平均值(CWM)来体现不同样地群落的功能特征[47]。个体总数大于10%的设定为优势科、优势种[48]。CWM计算公式如下:

表2 蜘蛛功能特征的类型及标准Table 2 Types and standards of spider functional traits
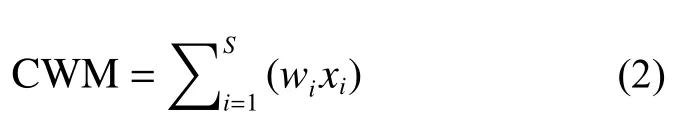
式中:S为总体的物种数,wi为第i个物种的相对多度,xi为第i个物种的功能特征值。
采用排序的方法检验蜘蛛群落功能特征与生产方式类型的关系。首先对不同样地的功能特征CWM值进行非约束性排序[49]; 其次将试验条件纳入环境因子进行约束性排序, 采用主成分分析(PCA)进行排序[50]。采用R3.6.1进行正态性检验、方差齐性检验和单因子方差分析, 采用PAST3.2进行多样性指数计算; 采用FD包[51]进行CWM值计算; 采用R3.6.1作图; 采用R3.6.1进行PCA分析和NMDS分析。
2 结果与分析
2.1 蜘蛛种类和数量
本研究共计捕捉了来自于13个科、25个属、35个种的共计2200头蜘蛛。其中, 物种数的优势科为肖蛸科(Tetragnathidae)、球蛛科(Theridiidae)、皿蛛科(Linyphiidae)和狼蛛科(Lycosidae), 分别占蜘蛛物种数的17.14%、14.29%、11.43%和11.43%, 个体数的优势科为狼蛛科、皿蛛科、跳蛛科(Salticidae)和球蛛科, 分别占捕获蜘蛛总数的22.41%、21.23%、14.95%和13.77%, 优势种分别为拟环纹豹蛛(Pardosa pseudoannulata) (14.86%)和驼背额角蛛(Gnathonarium gibberum) (12.41%), 以上2个种共占总捕获数量的27.27%。
2.2 不同稻田生产方式下蜘蛛α多样性
单因子方差分析(One-way ANOVA)以及K-W检验的结果显示, 不同类型稻田的蜘蛛物种丰富度(P=0.001)、多度(P=0.008)与辛普森多样性指数(P=0.007)均差异显著。
有机生产稻田平均可捕获22.8种蜘蛛, 显著高于常规生产(18.2)与绿色生产稻田(15.6)(图1a); 有机生产稻田蜘蛛多度与辛普森多样性指数显著高于绿色和常规生产稻田(图1b, 1c)。有机生产稻田则平均每样地可捕获592.4头蜘蛛, 显著高于常规生产稻田(168.8)和绿色生产稻田(118.8)。同样, 有机生产稻田辛普森多样性指数平均为0.728, 而绿色生产与常规生产稻田则分别为0.721与0.711, 显著低于有机生产稻田(图1c)。
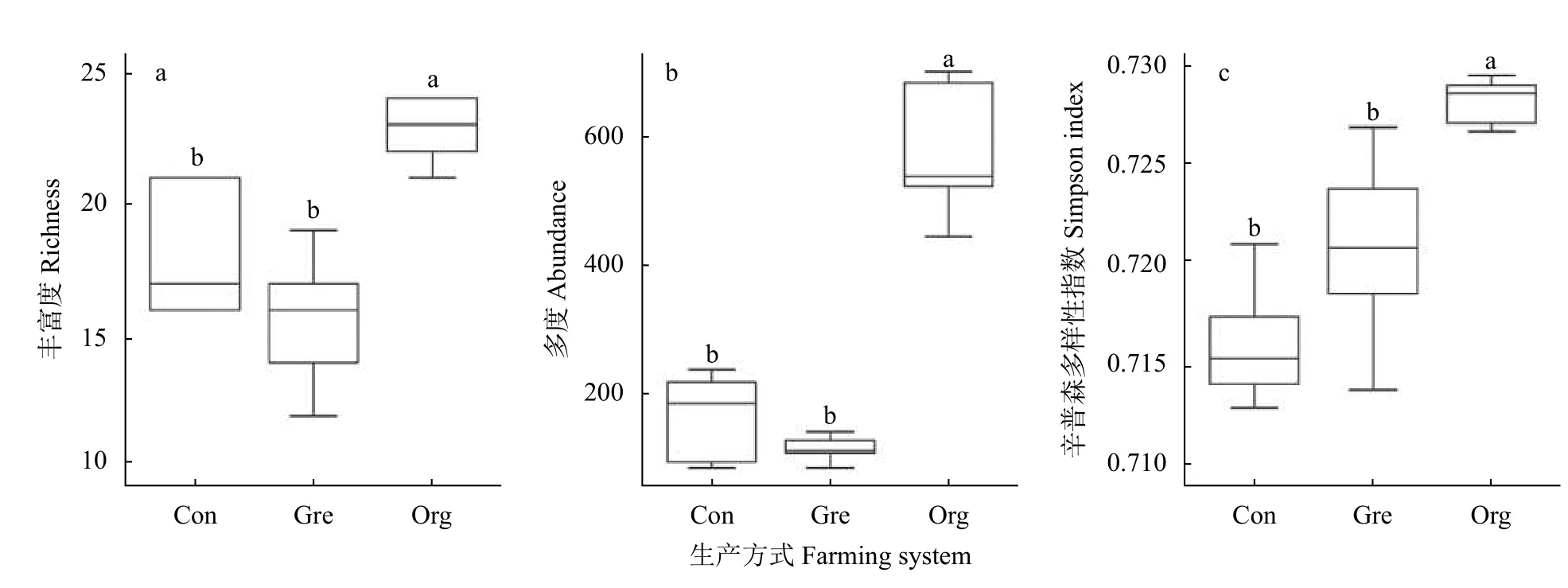
图1 不同生产模式稻田蜘蛛的物种多样性Fig.1 Diversity of spiders in rice fields under different farming systems
2.3 不同稻田生产方式下蜘蛛功能群多样性及功能特征组成
单因子方差分析结果显示, 仅有捕猎型蜘蛛占比在不同处理稻田差异显著(P<0.05)(图2b), 蜘蛛体长 (图2a)与蜘蛛飞航能力均无显著差异(图2c)。多重比较结果显示, 常规稻田中捕猎型蜘蛛比例达66.78%, 显著高于绿色生产稻田(51.91%)与有机稻田(52.65%)。
主成分分析(PCA)结果显示(图2d), 第1轴共计解释了55.84%的环境变量, 而第2轴则共计解释了39.37%的环境变量, 两轴共计解释了95.21%的环境变量, 较好地反映了不同功能特征之间的相互关系。其中, 代表捕猎方式的箭头和代表飞航能力的箭头夹角接近90°, 前者在后者箭头方向投影长度的绝对值接近于0, 这表明捕猎方式与蜘蛛的飞航能力无显著关联; 代表体长的箭头和代表捕猎方式、飞航能力的两个箭头方向相同, 且前者在后两个箭头的轴上拥有较长的投影长度, 这表明蜘蛛的体长与捕猎方式及飞航能力均有明显的正相关关系; 常规生产稻田主要分布在第1排序轴左侧, 与蜘蛛捕猎方式显著正相关; 绿色生产稻田主要分布在第2和第3象限; 有机生产稻田主要分布在第4象限。
2.4 不同稻田生产方式对蜘蛛群落组成的影响
对不同生产模式稻田蜘蛛群落结构分析显示,在强调优势物种(m=1)时, 除有机生产稻田样点相对集中, 与其他两种生产方式样点相分离, 说明有机生产下稻田蜘蛛优势物种群落结构相似, 但不同于其他两种管理模式(图3a); 在强调稀有物种(图3b)(m=5)时, 有机稻田蜘蛛群落相对聚集, 并与绿色和常规管理稻田相分离, 说明有机生产稻田稀有物种群落结构也相似, 且不同于其他管理模式。无论是强调优势物种(图3a), 还是强调稀有物种(图3b)时,绿色生产与常规生产稻田样点均存在一定程度的重叠, 说明绿色生产稻田群落和常规生产稻间相较于有机生产稻田具有更高的相似性。
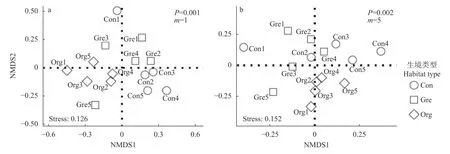
图3 基于非度量多维尺度分析(NMDS)与群落相似度分析的不同生产模式稻田蜘蛛群落组成Fig.3 Spider community groups of rice fields under different farming systems based on non-metric multidimensional scale analysis (NMDS) and community similarity analysis
进一步的指示种分析(Indval)结果显示(表3),绿色生产稻田与常规处理稻田缺乏显著(P<0.05)的指示种, 这一结果表示, 绿色生产稻田与常规处理稻田在群落的总体分布上以广布种与共有种为主, 缺乏特有种。而有机稻田则具有大量的指示种, 包括裂菱头蛛(Bianor incitatus)、八斑丽蛛、褶管巢蛛(Clubiona corrugate)和驼背额角蛛(Gnathonarium gibberum)等。这些蜘蛛在绿色生产稻田与常规处理稻田中很少分布, 仅在有机稻田中广泛分布, 且指示值达0.56~0.95, 说明这些物种在有机农田中稳定、广泛大量分布, 但在其他生境中很少出现。

表3 不同生产方式稻田蜘蛛指示种及其功能特征Table 3 Indicator species and their functional traits in rice fields under different farming systems
2.5 不同水稻发育时期蜘蛛多样性及功能性状变化
单因子方差分析结果显示, 有机生产稻田蜘蛛丰富度(图4a)和多度(图4b), 除水稻分蘖早期和拔节期外, 其余各时期均显著高于常规和绿色生产稻田; 辛普森多样性指数(图4c)于抽穗期和扬花期显著高于常规和绿色生产稻田; 常规生产稻田捕猎型蜘蛛占比(图4d)在水稻灌浆期显著高于有机生产稻田, 在拔节期和黄熟期显著高于绿色生产稻田。
蜘蛛丰富度随时间的变化趋势如图4a所示, 3种生产模式稻田蜘蛛丰富度均由分蘖早期的最低值(2.4、1.6、1.4)至抽穗期达最高值(14.0、7.8、7.6)。绿色和常规生产稻田蜘蛛多度(图4b)均由分蘖早期的最低值(2.6、3.6), 至抽穗期达最高值(16.4、27.2);有机生产稻田蜘蛛多度(图4b), 由分蘖早期的最低值(4.4)至灌浆期达最高值(99.0)。3种生产模式稻田蜘蛛辛普森多样性指数(图4c)均由分蘖早期的最低值(0.44、0.13、0.13), 至抽穗期达峰值(0.89、0.78、0.74), 随后趋于稳定。有机、常规生产稻田蜘蛛捕猎型占比由分蘖后期的最低值(0.33、0.17), 至抽穗期达到最高值(0.68、0.77); 绿色生产稻田蜘蛛捕猎型占比由拔节期的最低值(0.1), 至扬花期达到最高值(0.77)。整体上来看, 有机生产稻田蜘蛛丰富度、多度在水稻生长中后期仍保持较高水平; 3种生产模式稻田蜘蛛丰富度、辛普森多样性指数、捕猎型占比随时间变化趋势基本一致。

图4 不同生产方式稻田不同水稻发育时期蜘蛛多样性及功能特征的变化Fig.4 Changes of spider diversity and functional characteristics at different growing stages of rice under different farming systems
3 讨论
3.1 不同生产方式对稻田蜘蛛多样性的影响
农田管理措施会对蜘蛛群落组成、多度和丰度产生显著的影响[52-53]。稻田是蜘蛛的重要栖息地, 农药的密集使用和农业集约化往往会导致稻田蜘蛛多样性下降[54]。欧洲大量的研究证明, 禁止化学品的投入会对蜘蛛多样性产生积极的影响[55-56]; 国内的研究也表明, 大部分有机稻田的蜘蛛多样性高于常规稻田[57-58]。本研究中也得到了类似的结论, 相对于常规生产, 有机生产可以显著提高稻田蜘蛛的丰富度、多度和辛普森多样性指数。绿色生产是常规生产向有机生产过渡的一种生产方式, 相较于有机生产不使用农药、激素、化肥等化学投入, 绿色生产虽然在生产过程中允许使用农药和化肥, 但对用量和残留量有着严格的规定, 且不能使用国家禁止的高毒、高残留农药[59-60]。在本试验中, 绿色生产的稻田施用的是有机-无机复合肥, 并未过量施用氮肥, 这在蜘蛛多样性的保护上可以起到一定的促进作用[61], 但其在水稻生长期施用了3次杀虫剂, 而有机生产稻田病虫害防治主要依靠性引诱剂、竹醋、天然除虫菊素、苦参碱等生物农药。尽管绿色生产使用的都是低毒、低残留的化学杀虫剂, 但本研究结果显示, 相较于常规生产稻田, 绿色生产稻田对于稻田蜘蛛多样性的提高并没有明显地促进作用。这说明, 与肥料的施用相比, 太湖流域稻田蜘蛛可能对杀虫剂的施用更加敏感, 一旦施用化学源的杀虫剂, 即使是低毒、低残留的杀虫剂, 也会对其产生杀死或者驱逐的作用。此外, 即便采用部分生物源农药替代化学农药的方式, 也不能最大程度地发挥对蜘蛛多样性的保护, 且即使是低残留的化学农药, 分解也至少需要10年以上的时间[55]。另一方面, 也可能是有机稻田中较高的稻飞虱数量促进天敌多样性的提高。Martin等[62]指出, 景观中害虫群落的增殖速度往往较天敌更早、更迅速, 从而为天敌群落迅速增殖提供高密度的食物。因此, 在太湖流域稻田的生产经营中, 不能仅仅考虑对于稻田害虫的灭杀效果, 还应考虑天敌生物对于稻田害虫的取食, 综合考虑天敌和害虫的发生。
3.2 不同生产方式对蜘蛛功能组成的影响
在相同的生境类型中, 蜘蛛功能特征往往体现出相近的趋势, 这是不同蜘蛛对相同环境的趋同效应及生态位共同决定的[63], 但当其受到不同管理措施的干预后, 会发生趋异效益, 致使蜘蛛功能特征向着不同的方向发展。有机、绿色稻田因外界干扰相对较少, 灰飞虱、稻纵卷叶螟等害虫的数量相对较多,食物充足[56], 适应小体长结网型蜘蛛生存。而常规生产因外界干预相对较多, 食物相对较少[64], 只有活动能力和飞航能力较强、体长较大的捕猎型蜘蛛才能更好地适应此类环境。当生产方式由常规生产转化为绿色生产和有机生产后, 蜘蛛功能群更为丰富, 捕猎害虫的方式更加均衡, 总体虫害控制能力可能会相对提高。因此, 转换成有机生产模式后稻田害虫生物控制服务可能更高。
3.3 不同生产方式对蜘蛛群落结构的影响
本研究表明, 有机稻田蜘蛛的物种库相对于常规生产稻田有一定的差异, 出现了较多特有种, 而绿色生产和常规生产稻田蜘蛛群落缺乏显著的指示种,这进一步凸显了有机生产对于蜘蛛多样性保护的重要性。魏巍等[65]的研究表明, 于有机稻田中播种紫云英(Astragalus sinicus)作为绿肥, 可以在整个冬季中为蜘蛛提供栖息地和充足的食物来源, 并确保来年蜘蛛的多样性。本试验中的有机稻田在上一季水稻收割后, 也曾播种紫云英种子, 而绿色稻田播种油菜(Brassica napus)作为冬季覆盖作物、常规稻田没有种植冬季覆盖作物。这种管理措施上的差异, 可能是导致有机稻田蜘蛛多度和丰度较高的原因[66]。本研究中, 与常规稻田相比, 有机稻田中食虫沟瘤蛛(Ummeliata insecticeps)、驼背额角蛛等在冬播作物中越冬的蜘蛛明显增加, 验证了前文的猜测, 有机生产确实对越冬蜘蛛具有更好的保护作用。而绿色稻田中食虫沟瘤蛛、驼背额角蛛的数量没有发生显著增加, 可能是因为残留的化学除虫剂和杀虫剂会影响蜘蛛体内消化酶的活性, 进而影响蜘蛛的摄食量[67-69], 从而导致食虫沟瘤蛛、驼背额角蛛远离存在化学投入的绿色生产稻田, 迁徙到别的生境寻找食物。
水稻的生长季节通常是5月底到10月初, 这段时间大部分蜘蛛都能在稻田里生活, 但在其他时期蜘蛛必须寻找额外的栖息地, 田垄上的杂草往往被认为是主要的次生栖息地[70-71]。与绿色生产、常规生产稻田相比, 有机生产稻田田垄上拥有丰富的杂草, 可以为四斑粗螯蛛(Pachygnatha quadrimaculata)等结网型蜘蛛提供结网的物理结构, 且为拟水狼蛛(Pirata subpiraticus)、八斑丽蛛(C.octomaculata)等在水稻移栽前喜欢在杂草栖息的蜘蛛类群提供栖息地, 当水稻移栽后, 八斑丽蛛、拟水狼蛛会迅速从杂草中转移到有机稻田生境中[72-73]。同时, 在江苏地区水稻植株上部大部分为肖蛸科蜘蛛结网捕食[72], 一旦稻田中施用化学杀虫剂和除草剂, 肖蛸科蜘蛛受其影响较大, 这可能是有机稻田中肖蛸科的四斑粗螯蛛和狼蛛科的拟水狼蛛数量高于绿色和常规生产稻田的原因之一。由此可见, 拥有冬季覆盖物和杂草栖息地的有机稻田可能是提供研究区域农业景观中蜘蛛天敌多样性的重要物种库, 并为蜘蛛提供越冬场所。
3.4 不同水稻时期蜘蛛多样性及功能组成变化
在本试验中, 除分蘖早期、拔节期外, 其余各时期有机生产稻田蜘蛛丰富度和多度均显著高于绿色和常规生产稻田, 这表明有机生产方式在大部分水稻生育期中都对稻田蜘蛛丰富度、多度有显著的提升效果。分蘖早期, 3种生产模式稻田蜘蛛丰富度、多度无显著差异, 可能因为此时稻田内稻飞虱等蜘蛛食物资源不足[73-74], 无法满足蜘蛛数量和种类的快速提升; 拔节期, 3种生产模式稻田蜘蛛丰富度和多度发生下降, 因为该时期3种生产模式稻田均进行了病虫害集体防治, 此时草间钻头蛛(Hylyphantes graminicola)等蜘蛛以幼蛛为主, 耐受性相对较低[14],因而农药的施用对蜘蛛丰富度和多度产生了一定的影响, 同时, 该时期绿色生产稻田捕猎型蜘蛛比例发生明显下降, 可能是因化学农药的施用导致绿色生产稻田拟环纹豹蛛、拟水狼蛛等优势度较高的捕猎型蜘蛛逃离稻田生境, 前往田埂或者杂草根隙内以减轻化学农药对其产生的影响[75]。不过, 有机生产稻田蜘蛛多度从拔节期后一直增长, 而绿色生产和常规生产稻田在水稻扬花期蜘蛛多度再次发生下降,此时期有机生产稻田蜘蛛以成蛛为主[76], 耐受性增强, 加上生物农药对稻田蜘蛛影响相对化学农药较小, 从而蜘蛛多度可以继续保证增长, 而绿色生产稻田虽然较常规生产稻田减少了化学农药的施用量,但因蜘蛛对化学农药敏感, 即便低施用量也对其蜘蛛多度产生显著的影响, 导致蜘蛛多度发生下降, 并与常规生产稻田多度无显著差异。在水稻抽穗−扬花期, 3种生产模式稻田捕猎型蜘蛛占比均维持较高水平, 这可能与此时期4代纵卷叶虫进入产卵盛期,孵化出的幼虫为蜘蛛提供了充足的食物来源, 从而吸引稻田边缘生境的捕猎型蜘蛛来稻田内取食[77]。同时, 过高的农药施用量会导致稻飞虱等稻田害虫数量过低, 无法为蜘蛛等天敌提供充足的食物供应,破坏其食物链平衡[76]。因此, 在太湖流域生态敏感区域, 稻田生物和化学农药的最优种类、最佳施用量,还需要进一步研究。
3.5 有机操作时长对蜘蛛多样性的影响
本研究中, 有机生产开始时间仅为1年, 但显示了明显的生态效益, 有效提高了稻田蜘蛛多样性, 说明常规农田转换为有机生产可以在短期内有效提高蜘蛛多样性及其生态服务, 具有较好的生态效应。但He等[78]的研究结果表明, 有机生产5年、10年、15年稻田与常规稻田香农-威纳指数和蜘蛛群落组成无显著差异。这有可能是因多年有机生产后, 稻田生境中蜘蛛群落已达到稳定且饱和的状态, 食物无法供应所有的蜘蛛生存, 从而发生溢出效应, 某些原本倾向于在有机稻田中生存的特有种开始向周边常规稻田扩散, 也可能因为受稻田周围较高的半自然生境比例影响, 使得蜘蛛群落多样性发生显著的提高、群落结构发生明显的分化。因此, 转换生产方式对蜘蛛多样性的影响可能还受时间及周围景观结构的影响, 需要进一步的深入分析。
4 结论
稻田蜘蛛多样性和群落组成受生产管理措施影响, 有机生产稻田较绿色和常规生产稻田维持更高的蜘蛛丰富度和多度, 拥有更多的特有种, 捕猎型和结网型蜘蛛数量组成更加均衡, 害虫控制的作用可能更强。绿色生产方式虽然可以一定程度上提升蜘蛛多样性, 但效果并不显著。此外, 3种生产模式稻田蜘蛛体长和飞航能力无显著差异。因此, 在太湖流域, 采取有机生产方式种植水稻对保护稻田蜘蛛多样性、提升害虫生物控制服务和生态环境保护具有重要促进作用, 可以作为农业转型和改善生态环境的重要措施。此外, 建议在稻田病虫害集体防治时, 适当降低农药施用量以达到对稻田幼蛛的保护;绿色和常规生产稻田采取部分生物农药替代化学农药, 从而提高稻田蜘蛛的存活率; 水稻抽穗期和扬花期注意稻田周边半自然生境的维护, 尽可能提高非农生境的比例, 从而吸引更多的捕猎型蜘蛛来稻田内捕食害虫, 进而更好地发挥天敌蜘蛛的生物防治作用, 推动水稻绿色种植和农业的可持续发展。
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
学校教育研究(2020年7期)2020-04-09
文苑(2019年20期)2019-11-16
作文大王·低年级(2019年2期)2019-01-23
学苑创造·A版(2018年9期)2018-10-22
小朋友·快乐手工(2018年3期)2018-04-22
创新作文(1-2年级)(2018年7期)2018-01-22
小学阅读指南·低年级版(2017年6期)2017-06-12
小朋友·快乐手工(2015年1期)2015-03-13
