
基于个体的西北太平洋柔鱼冬春生群生活史早期生态模型构建
2021-09-09 09:34:56李曰嵩白松麟余为张瑜陈新军
海洋学报 2021年9期
李曰嵩,白松麟,余为,张瑜,陈新军*
( 1.上海海洋大学 海洋科学学院,上海 201306;2.上海海洋大学 海洋生态与环境学院,上海 201306;3.国家远洋渔业工程技术研究中心,上海 201306;4.大洋渔业资源可持续开发省部共建教育部重点实验室,上海 201306;5.远洋渔业协同创新中心,上海 201306)
1 引言
西北太平洋柔鱼(Ommastrephes bartramii)包括秋生群和冬春生群[1],其中的冬春生群是我国主要捕捞群体[2],生活在1 500 m水深以内[3],主要分布在温带和亚热带[1],寿命为1 a左右[4],属于短生命周期种类[5]。
前人在研究柔鱼时,主要通过柔鱼的捕捞产量和捕捞位置来研究生长趋势、资源量变动和渔场时空变化[6]以及分析群体与环境的关系、资源量或胴体各部位[7]等,然而鲜有对柔鱼在生活史早期的生长、死亡和输运的研究报道,但实际上生活史早期是柔鱼生命周期中最脆弱的时期,所以加强此方面研究十分重要。
柔鱼在生活史早期是随海流漂移的,自身没有游泳能力,一般把它们当成质点,然后在流场驱动其运动过程中增加跟周围物理环境相关的生长、死亡等生物过程,使其转化成有生命的“活”个体。采用的物理模型为FVCOM-Global,选取北太平洋作为本文的计算区域,获得北太平洋的物理环境信息。前人在研究柔鱼时一般是通过对采集的样本进行分析,没有考虑到每个个体都有自己的生长情况,为此本研究使用的生物模型为基于个体模型。基于个体模型是以个体而不是群体(种群)为研究对象,考虑到了个体之间存在的差异以及环境变化对个体发育的影响,更接近生物在自然环境中的生存状态,主要通过参数化描述一些过程,被学者认为是研究鱼类早期生活史最合理的方法[8],且个体模型在国外应用广泛,研究鳕鱼[9]和鲱鱼[10]的较多,国内也有少量研究,例如鳀鱼[11]和鲐鱼[12]等,但未见柔鱼研究相关报道。
本文将物理模型和生物模型耦合形成一个基于个体的西北太平洋柔鱼冬春生群生活史早期的生态模型来模拟冬春生群随物理环境的变化,了解冬春生群的生长、死亡、输运路径和丰度分布等情况,能为深入研究影响鱼类资源变动的动力学和生态动力学因素奠定基础。
2 研究方法
2.1 物理模型
采用的物理模型为FVCOM-Global,其是美国麻省大学开发的新一代非结构全球海洋模式[13],其外界强迫有潮汐、风、热通量、辐射、大气压、蒸发和降水等[13-14],能很好地应对物理环境复杂的问题[12]且近海区域精度较高[13],网格的水平分辨率在大洋内部最低约为50 km,而在近岸可达2 km[1],在垂直方向采用混合坐标,分45层。
用该模型模拟1997-2010年全球海域的三维物理场[13],并按每日进行存储。图1为本文的计算区域,范围为 11°~56°N,123°E~116°W。将该区域用三角网格进行划分,共有59 646个三角网格,三角网格点的数量为30 532个,以便在生物模型中设置亲体的位置。从图上可看出,计算区域覆盖了北太平洋的大部分海域,能够满足用于柔鱼生态模型模拟的广度和精度。
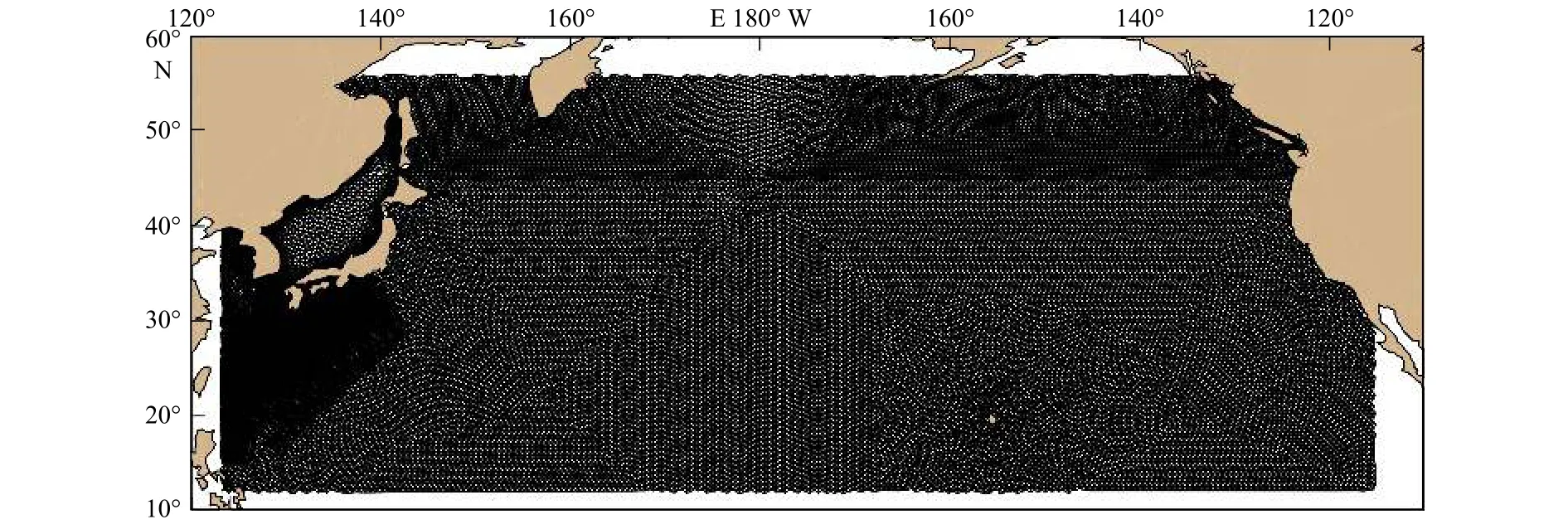
图1 由三角网格构成的模型计算区域Fig.1 The model computational domain is made up of triangular mesh
2.2 生物模型
本文采用的生物模型基于个体模型[15],简称IBM,该模型在研究生物的运动、生殖、集群、遗传与进化、早期生活史以及对环境变化的响应等方面都有应用实例[16],本文主要是利用该模型研究西北太平洋柔鱼冬春生群的早期生活史,描述和设置其产卵、生长和死亡等生物过程。将方舟[17]的研究与本文结合,把冬春生群的生活史分为6个阶段,即鱼卵、仔鱼(20~30 d)、仔稚鱼(30~50 d)、稚鱼(50~70 d)、亚成鱼(110~140 d)和成鱼(140~180 d)。因柔鱼后期游泳能力较强,故模型中只模拟到稚鱼阶段,并假设整个模拟过程中没有游泳能力且都是被动漂流。
2.2.1 产卵场的设置
北太平洋柔鱼冬春生群的实际产卵场为20°~30°N,130°~170°E 和 20°~30°N,130°~170°W[1](图2)。由于本文主要研究西北太平洋柔鱼冬春生群,因此需要在位于西北太平洋的实际产卵场内选取部分海域作为本研究的产卵场,但是有学者提出在西北太平洋产卵场内柔鱼主要位于 140°~150°E[18],因此,如图2蓝色圆形区域所示,在模型中将产卵场的位置设置为 20°~30°N,140°~150°E。
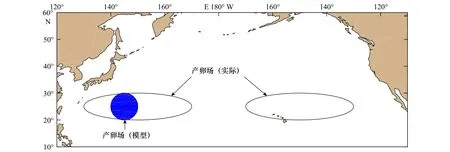
图2 北太平洋柔鱼冬春生群的产卵场示意图Fig.2 Schematic diagram of spawning ground of winter-spring cohort of neon flying squid Ommastrephes bartramii in the North Pacific Ocean
北太平洋柔鱼冬春生群的产卵时间主要为每年的1-5月[1],王易帆和陈新军[19]研究发现,西北太平洋柔鱼的产卵时间主要在1-3月。综上所述,模型中将开始产卵时间设为每年的2月1日,模拟到4月11日,共70 d左右。
2.2.2 繁殖力和产卵策略的设置
柔鱼的卵是包于卵袋里分批产出,每个个体的产卵量跨度范围较大,结合短生命周期中上层鱼类的生殖力以及对在大西洋捕获的柔鱼繁殖力的估算认为,其产卵量范围为 2×104~4.9×106个[17,20],在这个范围内选择105个作为每个个体的产卵量。
从陈新军等[21-22]、马金等[3,23]、魏广恩和陈新军[24]、Murata和 Nakamura[25]及 Nakamura[26]的研究发现,柔鱼晚上主要位于0~100 m水深处,白天主要位于100~700 m水深处,在水深1 500 m以内都有分布。此外,有学者在水深100 m内采集过仔鱼[5],因此推测柔鱼产卵位置可能位于上层,但对于精确的产卵水深至今未知。结合柔鱼的分布和采集到仔鱼的位置,本文将基于个体模型中的产卵水深设置为100 m。
模型中设定产卵场内的每个三角网格(面积约为30 km2)里有104个产卵亲体,每个产卵亲体的产卵量为105个,同时在100 m水深处开始产卵。由于卵的数量庞大,难以模拟每个个体真实的生长和死亡过程,因此模型中采用超级个体方法[15]进行研究。当每个三角网格里的产卵量超过一定数量(本文设置为108个)时形成一个超级个体,每个超级个体里个体的生长和死亡情况相同,用超级个体位置的变化来代替个体位置的变化。
2.2.3 生长和死亡的设置
柔鱼刚孵化时胴长在1.4 mm左右[5]。从Bigelow和Landgraf[27]的研究可知,柔鱼的日龄为35 d时胴长为12.1 mm,35 d以后柔鱼的生长率为1.1~2.5 mm/d;假设35 d以后柔鱼的生长率为1.8 mm/d,那么日龄为70 d时柔鱼胴长约为75 mm。
有很多学者研究过最适合柔鱼孵化的温度,王易帆和陈新军[19]的研究结论是22.5~26℃,余为等[5]、魏广恩等[24]和陈峰等[28]通过总结前人研究得到的结果是21~25℃。结合前人研究,本文在模型中将最适水温设置为23℃。
胴长计算公式[29]为
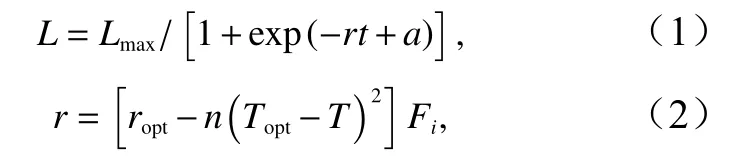
式中,L是胴长(单位:mm);Lmax是最大胴长(75 mm);t是日龄(0~70 d);a是常数(当t为 0 时,代入L的值为1.4 mm、Lmax的值为75 mm,求出a的值为3.962 2);r是中间变量;ropt是常数(0.12);n是常数(0.000 8);Topt是最适水温(23 ℃);T是真实水温(单位:℃),Fi是食物指数(0.5)。
生长率计算公式[29]为
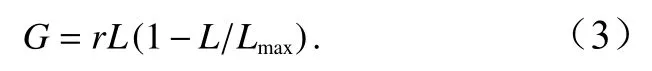
死亡率计算公式[29]为

通过调整上述各参数的值,观察结果对整体的影响而最终确定其中的参数和系数。
2.3 模型的耦合与设定
柔鱼在生活史早期没有游泳能力[1],而是被海流带到不同地方[12]。由于海流的方向和强弱不断变化,所以柔鱼在每一个时刻都会出现在不同位置,然后根据输运位置周围环境进行生长或死亡[12],如图3所示。柔鱼的位置变动属于物理层面,生长和死亡属于生物层面。因此,对于西北太平洋柔鱼冬春生群生活史早期生长、死亡和输运等过程的模拟研究,只利用生物模型单方面进行模拟不可能实现,必须将物理模型和生物模型两者耦合成一个生态模型才能实现[12]。
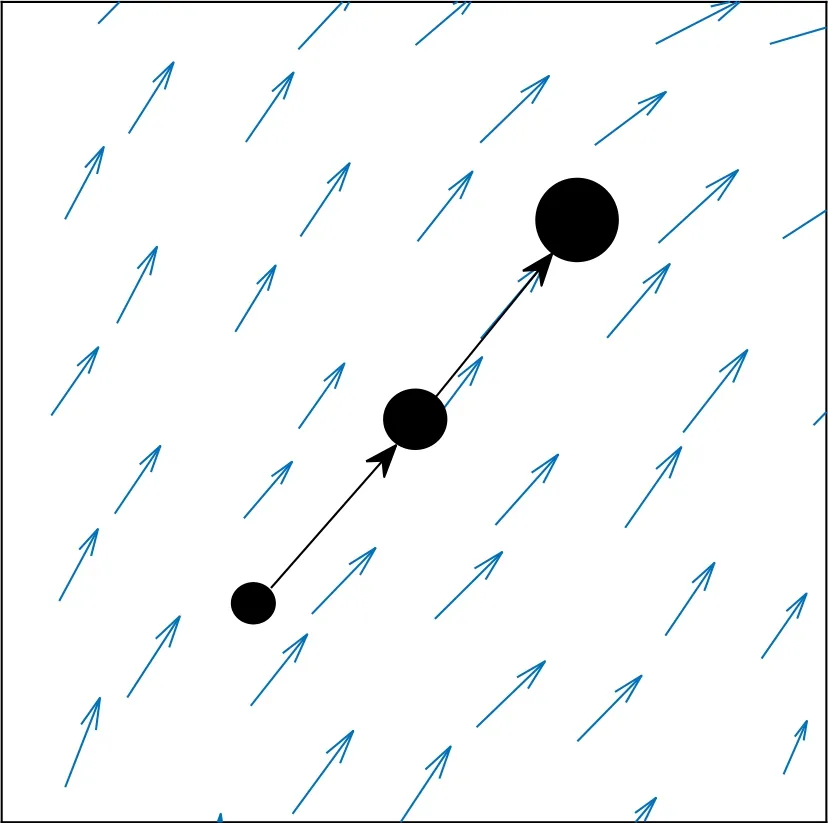
图3 柔鱼的输运和生长示意图Fig.3 Schematic diagram of transport and growth of neon flying squid Ommastrephes bartramii
将物理模型和生物模型耦合[12]的具体操作方法为(图4):在生物模型(IBM)中设置产卵场、水深、亲体量、最适温度、生长和死亡等初始条件,按照时间顺序读取从物理模型(FVCOM-Global)中得到的物理场数据(流场、水温等),将物理模型与生物模型通过拉格朗日质子追踪的方法进行耦合;用物理模型提供的物理场驱动个体进行位置变动,同时,个体被赋予生物模型中设置的生长特性,从各自的初始位置开始在物理场变动的条件下被输运到不同地点,接触到不同的物理环境,受周围环境影响进行生长或死亡,达到模拟个体在真实环境中生存状态的目的。为提高计算精度,在模型中设置每2 min计算一次,每12 h输出一次结果。每一次更新物理场并运行后都会产生相应的数据,包括超级个体的数量、每个超级个体所处的位置、总产卵数、每个超级个体里剩余个体的数量、日龄、水深、胴长和温度等生物信息。再通过对结果的整理与分析就能了解冬春生群在生活史早期的胴长变化、补充量、输运和分布等情况。
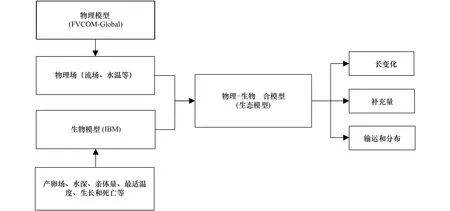
图4 生态模型的构建过程Fig.4 The construction process of ecological model
3 结果与分析
3.1 生长
从Bigelow和Landgraf[27]、Yatsu和Mori[30]的研究可知,柔鱼自孵化开始到至少35 d之内呈指数型生长,且在35 d时,胴长约为12 mm,以后逐渐变成线性生长。由于生物模型主要与温度相关,为了验证生长方程参数化的正确性,模拟画出了西北太平洋柔鱼冬春生群在不同水温条件下的胴长变化(图5),以23℃为最适水温,温度增高或降低都会使冬春生群胴长的生长速率减慢,这符合柔鱼生长规律。
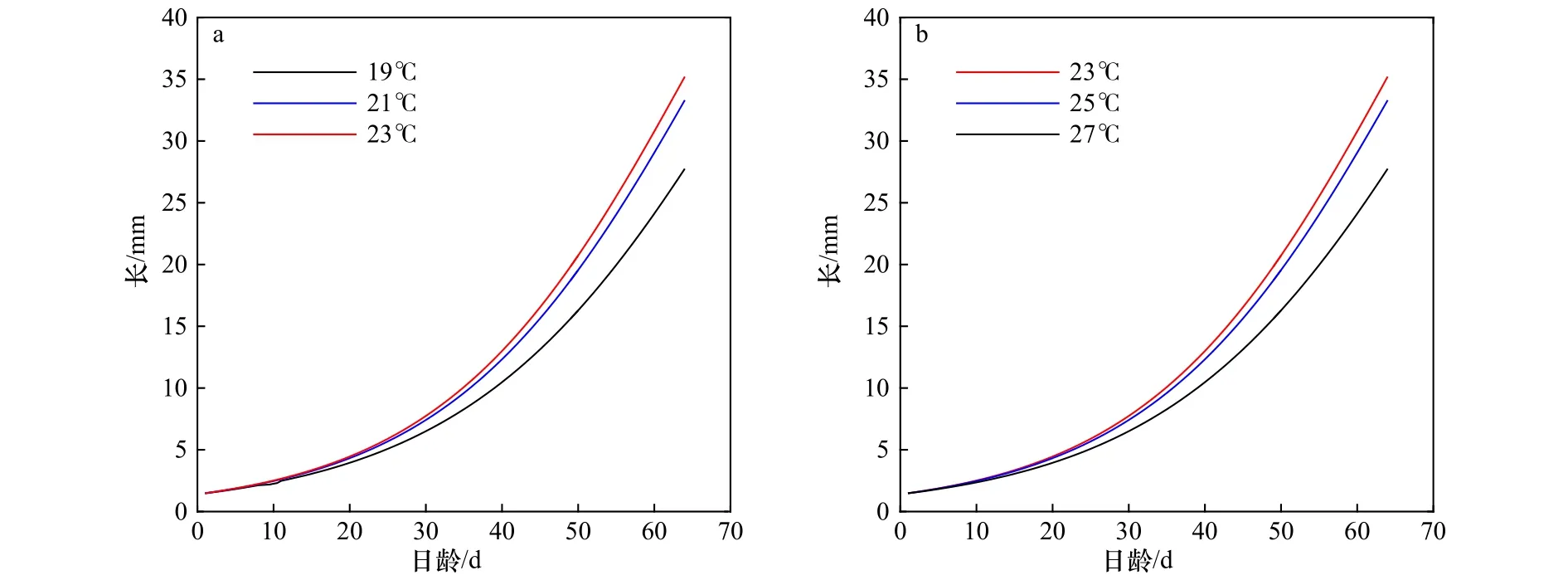
图5 西北太平洋柔鱼冬春生群在不同水温条件下的胴长变化Fig.5 Variation of mantle length of winter-spring cohort of neon flying squid Ommastrephes bartramii in the Northwest Pacific Ocean under different water temperature conditions
当温度为23℃时,第35天的胴长为10.08 mm,在第38天时可以达到11.76 mm;当温度为21℃和25℃时,胴长变化曲线与23℃时的曲线相差不大,第35天时的胴长为9.60 mm,胴长达到12.00 mm所需要的日龄为39~40 d;当温度为19℃和27℃时,第35天时的胴长为8.28 mm,第43天时的胴长为12.01 mm。第3条线与前两条线相差较大,生长速率明显减慢,但是这几条线整体变化趋势基本一致,即从孵化后到40 d前后呈指数型生长,之后逐渐变为线性生长。综上所述,生长变化曲线的模拟结果与前人的研究结果基本相似,说明在生物模型中对冬春生群设置的与生长有关的参数是合理的。
3.2 补充量
模型中设置的初始产卵时间为每年的2月1日,随着时间流逝,产生的卵越来越多,当达到一定数量时形成一个超级个体。产生第1批超级个体的时间为2月初,到2月中旬达到最大值(7 029个),卵的总数是7.2×1011个,为产卵高峰期,符合分批产卵[17]的特点。
1997-2010 年冬春生群模拟补充量的变化如图6所示,1997-1999年模拟补充量逐年升高,1999-2002年逐年下降,2002-2010年又回到整体升高的趋势。由于物理环境的不同,模拟补充量存在年际波动,并且在一些年份差异较大。其中模拟补充量最低的年份是2002年,其次是2003年,1999年最高,2005年仅次于1999年。
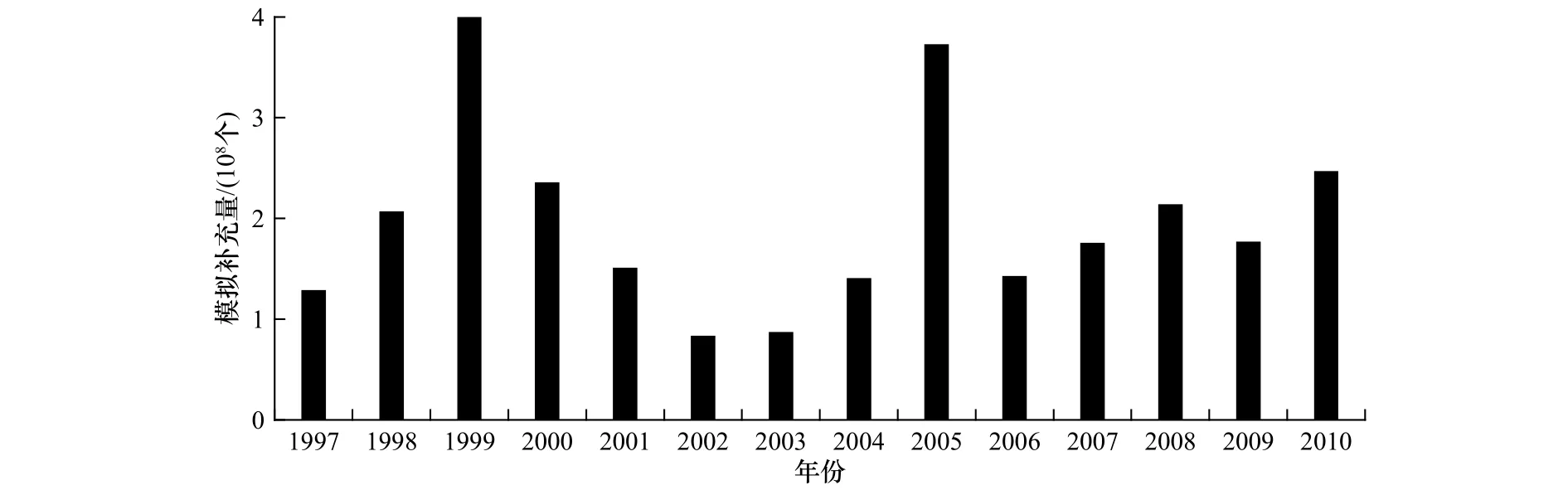
图6 模拟补充量Fig.6 Simulative recruitment
模型中设置的亲体量每年都是相同的,得到的结果为亲体量相同时不同年份的补充量情况(模拟补充量),但是实际上研究区域内的亲体量每年都不一样。为了了解亲体量不同时的补充量情况,列出余为[1]总结的中国在1997-2010年西北太平洋柔鱼渔获量(表1),取14 a内渔获量的平均值,用每一年的渔获量除以平均渔获量得出各自对应的系数,再乘以模拟补充量,对年际补充量进行调整,就能得到亲体量不等时的补充量,即实际补充量。
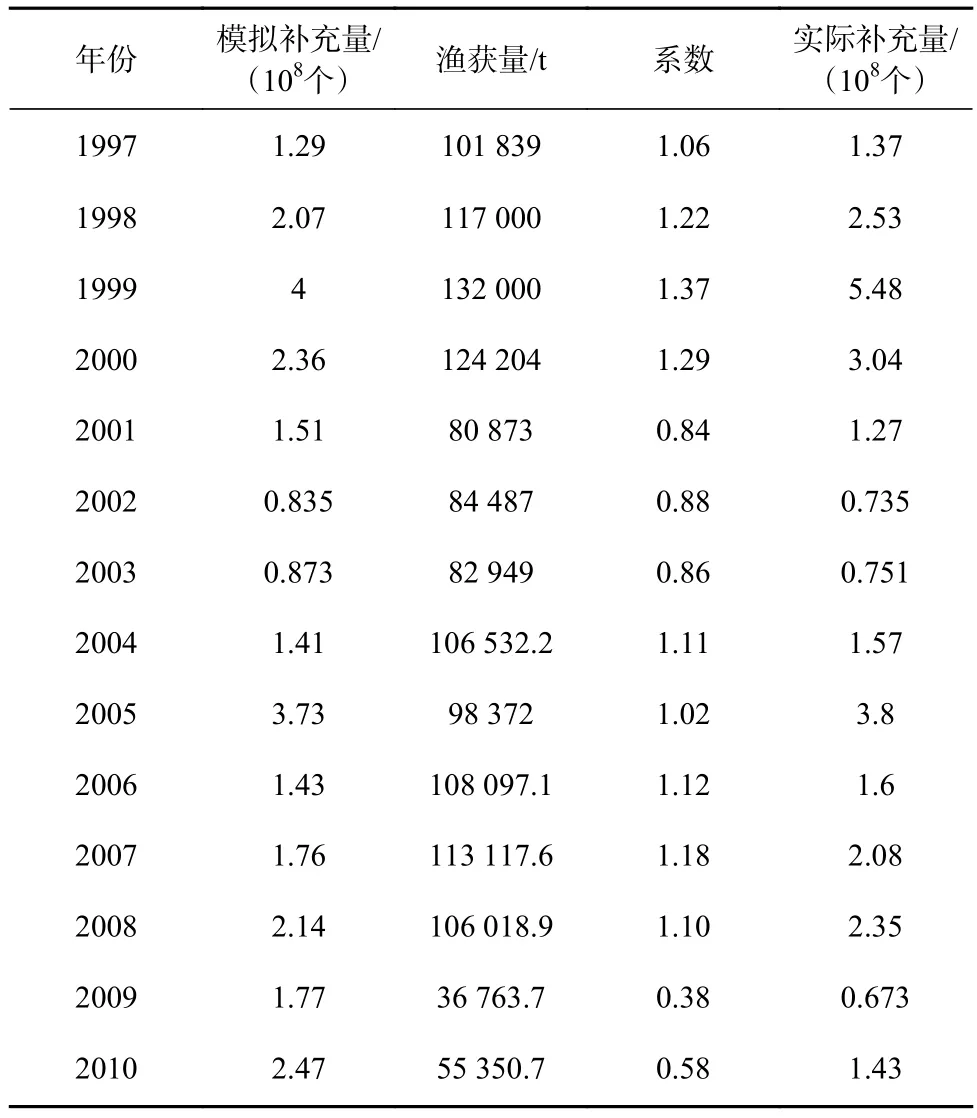
表1 补充量和渔获量Table 1 Recruitment and catch
将实际补充量与模拟补充量进行对比(图7),年际变化趋势基本相似,变化较大的是1999年、2009年和2010年。1999年的渔获量最高,2009年和2010年的渔获量最低,因此,对这几年模拟补充量的调整较明显,从而对补充量整体变化有一定影响,使数据更接近实际情况。
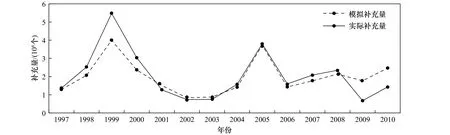
图7 模拟补充量与实际补充量对比Fig.7 Comparison of simulative recruitment and actual recruitment
将实际补充量与渔获量进行对比(图8):从最高点和最低点来看,实际补充量和渔获量最高的年份都是1999年,最低的年份都是2009年;从变化趋势来看,1997-1998年补充量和渔获量都增加,到1999年达到最大值,之后到2001年呈下降趋势,2001-2003年变化不大,比较平稳,之后到2005年整体呈上升趋势,2006-2008年波动不大,2008-2010年整体呈下降 趋势,其中的2009年达到了最小值。1997-2010年的实际补充量与渔获量的整个变化趋势基本一致,证明模型中设置的与死亡情况有关的公式是合理的,可以用来计算每年的补充量情况。
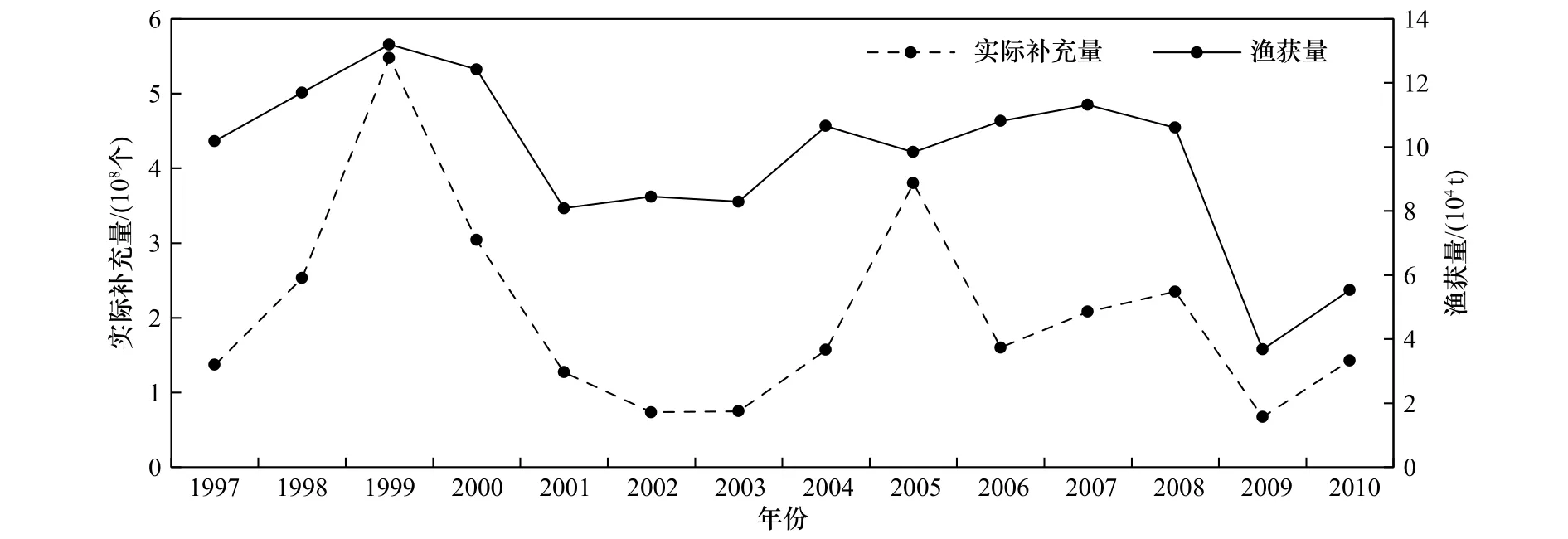
图8 实际补充量和渔获量对比Fig.8 Comparison of actual recruitment and catch
3.3 输运和分布
3.3.1 水平方向
模型中是通过改变物理场来达到冬春生群位置变化的效果,从而实现模拟冬春生群在生活史早期年际输运和分布情况的目的。从1997-2010年水平输运分布结果来看,与产卵场西北部很明显的强流相比,产卵场内的海域属于弱流区,大部分柔鱼分布于产卵场内或产卵场周围,说明在生活史早期,冬春生群主要栖息于产卵场,但是在整体上有向北或东北方向输运的趋势,模拟结果与Nishikawa等[31]的研究相似。然而由于物理环境不同,其在水平方向上的输运和分布依然存在差异。
从1997-2010年模拟阶段内输运到四周最远的柔鱼相对于产卵场的距离(表2)可以了解到,这14 a内,在南北方向上,有8 a向北输运得更远,在东西方向上,有11 a向西输运的距离大于向东输运的距离,从这个角度对比柔鱼在东西方向上的输运更明显。
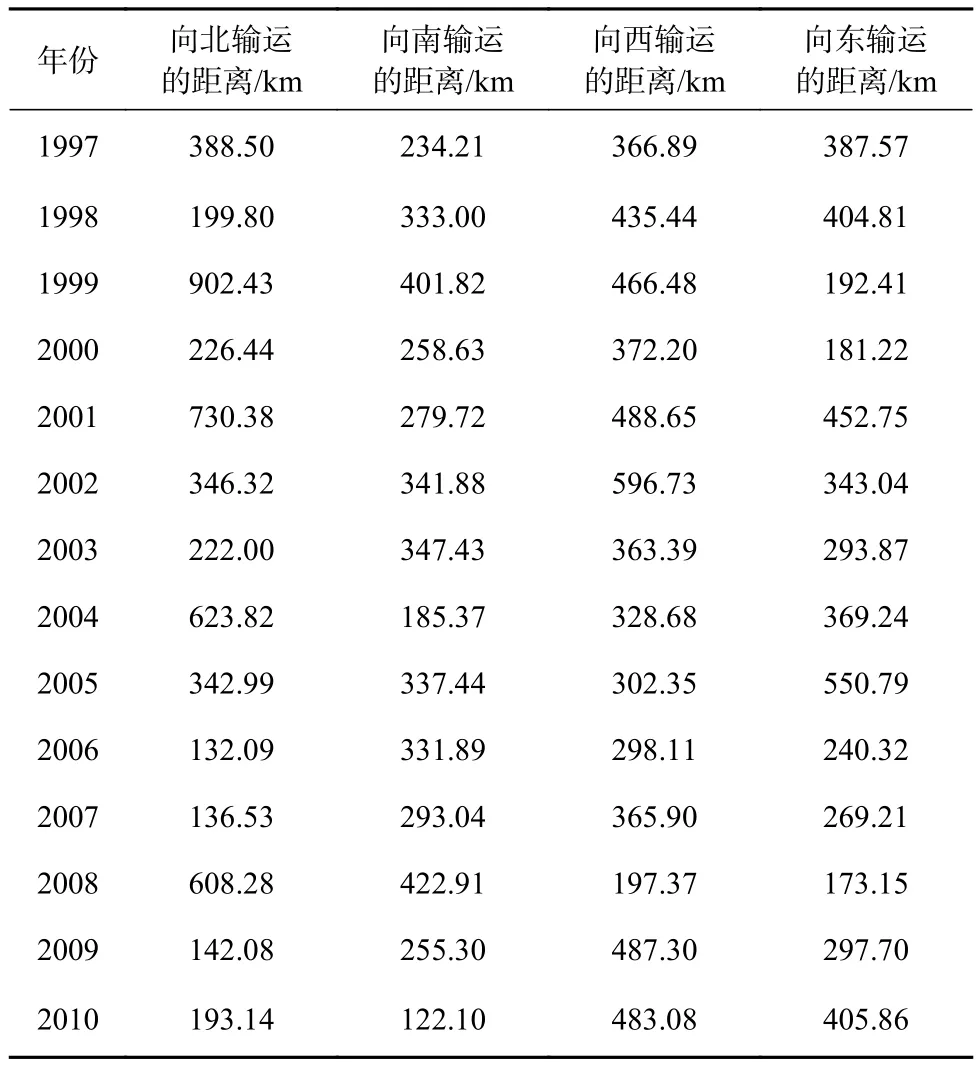
表2 输运到四周最远的柔鱼相对于产卵场的距离Table 2 The distance to spawning ground of the farthest neon flying squid Ommastrephes bartramii transported to the surrounding waters
再比较一下柔鱼被输运到不同海域的比例。产卵场的中心点是 25°N,145°E,将 25°N 和 145°E 作为分界线,把柔鱼的分布区域分为4部分,通过计算模拟结束后每个区域内柔鱼的占比(图9a)加深对其输运情况的认识。其中,被输运到西北方向的柔鱼最多,西南仅次于西北,然后是东北方向,最少的是东南方向。再将柔鱼分布区域在经度上分为145°E以东和 145°E 以西,在纬度上分为 25°N 以南和 25°N 以北,统计被输运到这4个海域的柔鱼占柔鱼总数的比例(图9b),能看出被输运到产卵场西部的柔鱼最多,其次是北部,然后是南部,最少的是东部,即位于西部和北部的柔鱼数量基本分别超过了位于东部和南部的柔鱼数量。将图9b与图9a相比能较明显地看出柔鱼的输运方向,结合两图来看初步认为柔鱼主要向西部和北部输运。

图9 不同海域内柔鱼的占比Fig.9 The proportion of neon flying squid Ommastrephes bartramii in different sea areas
然而,只从柔鱼相对于产卵场的距离以及在各个海域内的占比两方面还无法对其输运进行更全面的了解,需要结合柔鱼在模拟阶段的分布和流场图(图10)来分析其输运的规律和影响输运的原因。

图10 1997-2010年西北太平洋柔鱼冬春生群的分布和流场Fig.10 Distribution and flow field of winter-spring cohort of neon flying squid Ommastrephes bartramii in the Northwest Pacific Ocean from 1997 to 2010

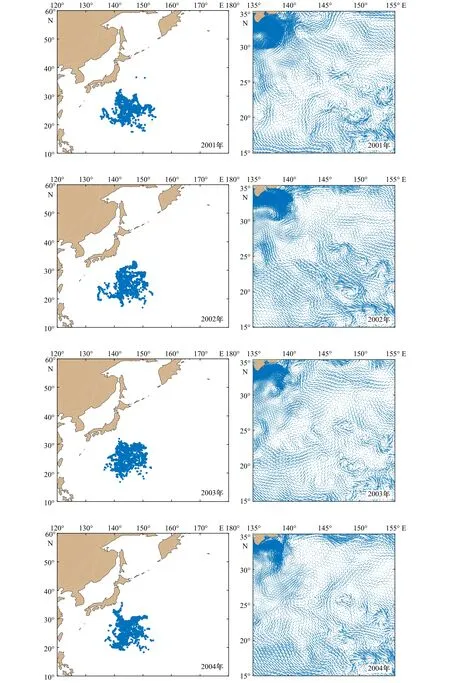
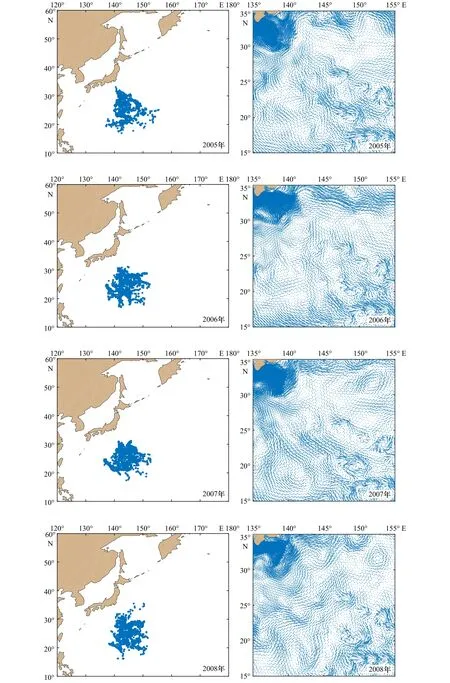
从图10可以看出,在模拟阶段大部分柔鱼还是位于产卵场内,尤其是1998年、2000年、2003年、2006年和2007年,其他年份的小部分柔鱼在海流的作用下被输运到各个方向。
向北输运较明显的有1997年、1999年、2004年和2008年,但向北输运又分为几种情况,主要是由于28°N以北海域柔鱼分布的差异。1997年一部分柔鱼向北输运,一部分向东北输运;1999年向西北、北和东北输运都较明显;2004年的输运情况与1997年类似,不同的是向北输运得更明显;2008年向东北输运得更明显。这是因为在 28°~35°N,135°~145°E 存在强流海域,使28°N以北的柔鱼受到强流的影响较大,即使是原本向西北方向输运的柔鱼在经过这里时也会以较快的速度向北或东北输运,表现出明显向北或东北方向输运的趋势。此外,还有由于向多个方向输运较平均而没有表现出明显输运方向的年份,如2001年、2005年和2010年。
在产卵场的东南方向存在着很多小范围且不规则涡旋的海域,就是这些涡旋的存在使得产卵场东南部柔鱼的输运无规则。除此之外,产卵场附近还分布着黑潮逆流[1],而且在模拟阶段主要位于35°N以南,因此,冬春生群除了会受到海洋中各种变化和气候等因素的影响,还会被黑潮逆流影响,部分向西部和南部扩散的柔鱼就符合在黑潮逆流作用下的结果。但是绝大多数向西部输运的柔鱼主要是由于受到25°N以南余流方向的影响,表现最明显的是2002年和2009年,这使得有一部分位于西南部和东南部的柔鱼向西部输运,该海域内余流强度较大,使得在模拟阶段柔鱼向西输运的距离较远,同时导致西北部与西南部柔鱼的占比较高,又由于25°N以北余流的方向逐渐由西北转向北,尤其是在28°N以北表现得更明显且有向东北转向的趋势,所以部分向西部和西北部输运的柔鱼最终还是会向北或东北部输运,同时位于东北方向的柔鱼也比位于东南方向的柔鱼占比大,且东南方向的柔鱼占比最小。3.3.2 垂直方向
为了观察本文设定的初始条件是否会使仔鱼出现在100 m水深内,将冬春生群在模拟时间内鱼卵、仔鱼、仔稚鱼和稚鱼这4个阶段分别处于各个水深范围内的比例进行统计,如图11所示,100 m水深内仔鱼的占比将近总数的一半,说明在水深100 m内能采集到仔鱼。此外,Bower[32]在夏威夷群岛采集仔鱼时发现,在22~22.5℃时柔鱼仔鱼丰度最大,本文通过计算得到在21.10~22.66℃时仔鱼的丰度最大,与Bower的研究基本相似。
根据图11可知,从鱼卵、仔鱼、仔稚鱼到稚鱼,位于0~200 m水深内的柔鱼占比逐渐减小,在200~300 m水深内仔鱼和仔稚鱼的占比较大,300~400 m范围内仔稚鱼和稚鱼的占比较大,从400 m开始水越深稚鱼的占比越大。这说明在生活史早期,随着时间的流逝,一部分柔鱼会被输运到更深的水层。
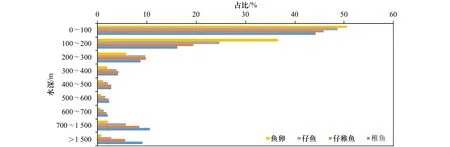
图11 冬春生群在不同生长阶段处于各个水深范围内的比例Fig.11 The proportion of winter-spring cohort in different water depth ranges at different growth stages
为了了解冬春生群在整个模拟阶段内垂直方向上的分布,将其在不同水深处的比例列出来,如表3所示。在模拟的14 a里,位于水深700 m以内的柔鱼平均占比为90.00%,位于水深1 500 m以内的柔鱼平均占比为95.98%。与柔鱼主要位于0~700 m以内[3,21-26]以及在水深1 500 m以内都有分布[3]的结论基本一致。虽然模拟结果显示1 500 m以深还有少量柔鱼存在,但这里的环境不适合其生长,因此它们基本不能存活,会在一定时间内死亡。

表3 整个模拟阶段内冬春生群分布在不同水深处的比例(%)Table 3 The proportion of winter-spring cohort distributed in different water depths during the whole simulation stage(%)
4 结论
本文从物理模型、生物模型以及模型耦合与设定等几方面构建了基于个体的西北太平洋柔鱼冬春生群生活史早期生态模型,初步模拟了冬春生群在生活史早期的生长、补充量、输运和分布等过程,并与前人的研究成果进行对比验证了该模型的准确性。
在生活史早期,冬春生群极易受周围环境影响,随着胴长的增加和身体各部分的发育,应对环境的能力不断增强。为使冬春生群一直保持被动输运的状态,本文将模拟时间设置为0~70 d。若时间范围设置太短,会使冬春生群输运的距离较短,从而增加分析其输运规律的难度;若时间范围设置太长,冬春生群生长到一定程度就会拥有游泳能力,使模拟过程出现偏差。因此,可以通过微调时间范围尽量减小误差。
从生长情况来看,在水温范围为19~27 ℃时,越靠近最适温度23℃,冬春生群生长越快。不同水温的胴长生长曲线均先为指数型生长,再转变为线性生长;达到12 mm的日龄至少需要38 d。该结论与前人[27,30]得到的柔鱼在35 d时胴长约为12 mm以及至少在35 d之内的生长曲线呈指数型的结论基本一致。
在该模型中,模拟的第1周内产生了第1批超级个体,20 d内超级个体的数量达到了最大值,符合柔鱼分批产卵的特点。从模拟结果来看,在1997-2010年范围内,若每年冬春生群的亲体量相等,补充量最高和最低的年份分别是1999年和2002年;在考虑到实际情况是每年亲体量不等的情况下,根据实际渔获量对模拟补充量进行了调整,得到的实际补充量与余为[1]的研究相符,最高和最低的年份分别是1999年和2009年,变化趋势也基本相似。
通过对1997-2010年柔鱼冬春生群输运分布和流场情况进行分析发现,在生活史早期,产卵场所在海域是弱流海域,有相当一部分柔鱼仍然位于产卵场内,但是结合向四周输运最远的柔鱼相对于产卵场的距离和柔鱼分布于各个海域的比例可发现,在25°N以南柔鱼主要向西和西北方向输运,在25°~28°N主要向西北、北和东北方向输运,这也使得位于西北部的柔鱼占比最多且向西输运距离较远,但由于在28°~35°N主要向北和东北方向输运且由于这里有强流的存在而使其整体有明显向北或东北方向输运的趋势。垂直方向上,在仔鱼阶段,有将近一半的仔鱼分布在0~100 m水深范围内;在整个模拟阶段,一部分柔鱼会随时间的流逝逐渐被输运到更高的水层,但主要还是位于0~1 500 m且在该水深内的占比为95.98%,少量被输运到1 500 m以外的柔鱼会由于周围环境不适合生存而在短时间内死亡。西北太平洋物理环境较复杂,水深几千米并且有多种生物分布,同一地点的不同时刻环境因子都不尽相同。海水在水平和垂直方向上的流速、温度和其余的各种环境因子都会影响冬春生群的输运、分布和生存等情况。大量的变量使得对西北太平洋海域的认识达不到全面了解,更无法一模一样模拟该海域的物理环境,只有不断完善和改进模型使之能接近实际情况。
在模型里设置冬春生群生长与死亡情况的公式来模拟真实的生长和死亡过程,这难免存在偏差,在以后的研究中可通过调整初始参数观察最终结果,使其不断接近真实情况,还可研究是否能把更多变量加入到模型中,如海水盐度、叶绿素浓度和其他生物等信息。最后,通过改变最适温度、产卵时间、产卵水深、产卵场的位置、刚孵化时的胴长与最大胴长等初始条件来探讨各方面对整个群体的影响,从而为将来学者们对渔业资源的管理和可持续利用等研究提供参考。
猜你喜欢
水生生物学报(2023年3期)2023-03-10 03:35:24
——以泉吉河为例
生态学报(2022年22期)2022-12-16 04:50:20
长江科学院院报(2021年8期)2021-04-03 00:19:52
农家之友(2020年9期)2020-04-21 02:19:04
劳动保护(2019年7期)2019-08-27 00:41:02
中学历史教学(2016年4期)2016-11-11 07:11:35
长江师范学院学报(2016年1期)2016-08-23 11:40:08
中国社会历史评论(2016年2期)2016-06-27 07:11:56
学习月刊(2015年22期)2015-07-09 03:40:48
中学科技(2015年1期)2015-04-28 05:06:12
