
基于细胞色素B(Cyt b)基因对翘嘴鲌不同群体遗传多样性的分析
2021-09-03 02:48:36张桂宁方弟安薛向平张敏莹冯晓婷杨雪军
中国农学通报 2021年23期
张桂宁,方弟安,,薛向平,张敏莹,冯晓婷,杨雪军
(1上海海洋大学水产科学国家级实验教学示范中心,上海 201306;2中国水产科学研究院淡水渔业研究中心,农业农村部长江下游渔业资源环境科学观测实验站,江苏无锡 214081)
0 引言
翘嘴鲌(Culter alburnus Basilewsky)隶属于鲤科(Cyprinidae)、鲌亚科(Cultrinae)、鲌属(Culter),俗称白鱼、翘嘴白等,是鲌亚科中最大的一种,是太湖“三白”之一,与松江鲈(Trachidermus fasciatus)、黄河鲤(Cyprinus carpio)、松花江鲑(Siniperca chuatsi)被誉为中国四大名鱼[1]。翘嘴鲌在中国分布广泛,几乎各个江流湖泊中都有存在。翘嘴鲌多2~3龄性成熟,生殖季节在5—7月,5月中旬为盛产期,具有溯河产卵特性,卵黏性,受精卵黏附于水生植物或砂石上发育[1]。翘嘴鲌生长迅速,耐低氧,抗病力强,适合高密度养殖,是长江三角洲地区的主要名特淡水养殖品种[2]。鉴于翘嘴鲌的重要经济价值和生态地位,受到众多学者的广泛关注与研究,目前主要集中在生物学特征研究[3]、资源现状[4]、育种[5]胚胎发育[6]、繁殖[7]、病害[8]和遗传多样性[9-11]等方面。
研究遗传多样性对生物多样性保护和生物资源可持续利用有重要意义,由于线粒体DNA(Mitochondrial DNA,mtDNA)具有母系遗传、进化速率快等特性,因此,常用作研究亲缘物种遗传多样性的理想分子标记[12]。在mtDNA的13个蛋白编码基因中,细胞色素b基因(Cytochrome b,Cyt b)的结构和功能较为清楚,碱基含量的富集反映出Cyt b基因在密码子使用上的偏倚性[13-14]。Cyt b基因进化速率适中,包含着从种内到种间的较强进化信号,故被广泛地用来进行物种多样性和系统进化等方面的研究[15]。王利华等[16]探讨基于CO I和Cyt b DNA条形码鉴定6种鲌属鱼类(达氏鲌、海南鲌、尖头鲌、蒙古鲌、拟尖头鲌和翘嘴鲌)的可行性,通过计算其碱基含量、群体间的平均遗传距离、种内遗传距离和构建系统进化树,研究显示,线粒体CO I和Cyt b基因可以作为DNA条形码鉴定鲌属鱼类。王伟[17]应用扩增片段长度多态性(AFLP)、内部简单序列重复(ISSR)和DNA序列(COII和D-LOOP)分析技术,对中国不同地理的7个群体翘嘴鲌遗传结构和多样性进行了分析,并对各种标记的结果进行了横向比较,研究显示AFLP标记要略优于ISSR标记技术,两种序列均能提供一定的遗传信息,但D-LOOP序列进化速度更快,另外中国野生翘嘴鲌遗传多样性比养殖群体丰富,遗传结构合理。黄小彧[11]针对长江水系不同江段的14个翘嘴鲌群体,应用线粒体控制区研究其遗传多样性,发现不同江段的翘嘴鲌群体遗传多样丰富度不同,表现出高单倍型多样性和低核苷酸多样性共存的现象,遗传多样性较低[11]。
长江是中国第一大河,对其渔业资源遗传多样性的研究极其重要,太湖、淀山湖和长荡湖均是位于长江下游的湖泊,是翘嘴鲌的良好栖息场所,近几年,这些湖泊都有不同程度的富营养化,翘嘴鲌资源有所衰退[18]。翘嘴鲌是“太湖三白”之一,在淀山湖和长荡湖中亦为渔业资源的优势物种。近年来,基于线粒体DNA对长江下游水域翘嘴鲌遗传多样性的研究较少。因此,本研究以长江、长荡湖、太湖和淀山湖4个水域的翘嘴鲌为研究对象,通过mtDNA Cyt b基因核苷酸序列的比较分析,探讨长江下游4个水域翘嘴鲌群体的遗传多样性、遗传结构和群体间的进化关系,以深入了解其遗传背景和种质资源现状,为翘嘴鲌的野生资源保护提供基础数据。
1 材料和方法
1.1 实验材料
2019年5—8月,使用定制丝网和流刺网分别于淀山湖(DSH)、太湖(TH)、长荡湖(CD)和长江(CJ)水域的固定采样位点随机捕获收集翘嘴鲌尾鳍样本,浸泡于无水乙醇中保存,其中淀山湖47尾,太湖40尾,长荡湖40尾,长江84尾,共计211尾。
1.2 DNA提取
将鳍条样品剪取约30 mg的组织样品于离心管中,使用试剂盒法提取DNA(海洋动物组织基因组DNA提取试剂盒,天根生化有限公司),并用浓度1%的琼脂糖凝胶电泳检测DNA的浓度和纯度,符合后续扩增要求的DNA样品置于-20℃保存备用。
1.3 PCR和DNA测序
PCR扩增Cyt b基因所需引物序列为Cytb-F:5-GACTTGAAAAACCACCGTTG-3 Cytb-R:5-CTCCG ATCTCCGGATTACAAGA-3。
PCR反应体系:总体积25 μL,包括dNTP Mixture 12 μL,上、下游引物各1.25 μL(50 μmol/L),DNA 2 μL,ddH2O 8.5 μL。PCR扩增程序:94℃预变形5 min;94℃变性10 s,55℃退火10 s,72℃延伸30 s,35个循环;72℃延伸5 min;4℃结束。2%琼脂糖凝胶电泳检测PCR产物后,将条带清晰扩增有效的PCR产物送亦欣生物科技(上海)有限公司进行双向测序,测序引物与扩增引物一致。
1.4 数据分析
采用MEGA 7.0.26软件中的Align by clustalW软件进行Cyt b基因原始序列的比对、剪切。用DNASP v5[19]软件计算群体单倍型,DNASP v5软件进行Tajima's D和Fu’s Fs的中性测试,以衡量群体是否发生了扩张。采用Arlequin 3.5软件[20]进行遗传多样性、群体的分子方差分析(Analysis of molecular variance,AMOVA)、遗传距离和基因流(Nm)值的计算。在MEGA 7.0.26软件中以Kimura双参数法(Kimura-2-parameter)为替代模型,采用NJ法(Neighbour-joining,NJ)构建翘嘴鲌不同群体单倍型的分子聚类树,从分支关系分析各群体之间的亲缘远近关系。
2 结果与分析
2.1 序列特征
PCR产物基因测序之后经MEGA7.0.26软件中的Align by Clustal W进行原始序列的比对剪切,得到1088 bp的序列,与NCBI数据库的翘嘴鲌Cyt b序列比对,数据同源性达到99%以上。MEGA7.0.26软件分析211个个体序列的碱基组成平均含量为T(27.1%),C(23.1%),A(27.9%),G(21.9%),A+T含量为55%,G+C为45%。表1中211个个体共有46个单倍型,单倍型多样性在(0.849±0.004)~(0.946±0.015),4个群体中太湖群体的最高,其次是长荡湖群体,再次是长江群体,淀山湖水域的翘嘴鲌单倍型最低。核苷酸多样性在0.0083~0.174,整体而言核苷酸多样性较高,其中淀山湖群体>长荡湖群体>长江群体>太湖群体。MEGA7.0.26软件中分析得到单态突变位点46个,简约信息点1011个,变异位点1057,变异率为97.15%,保守位点31个。DNASP软件分析得到258个多态性位点,约占核苷酸总数的23.71%。转换与颠换比值(si/sv)为0.7。
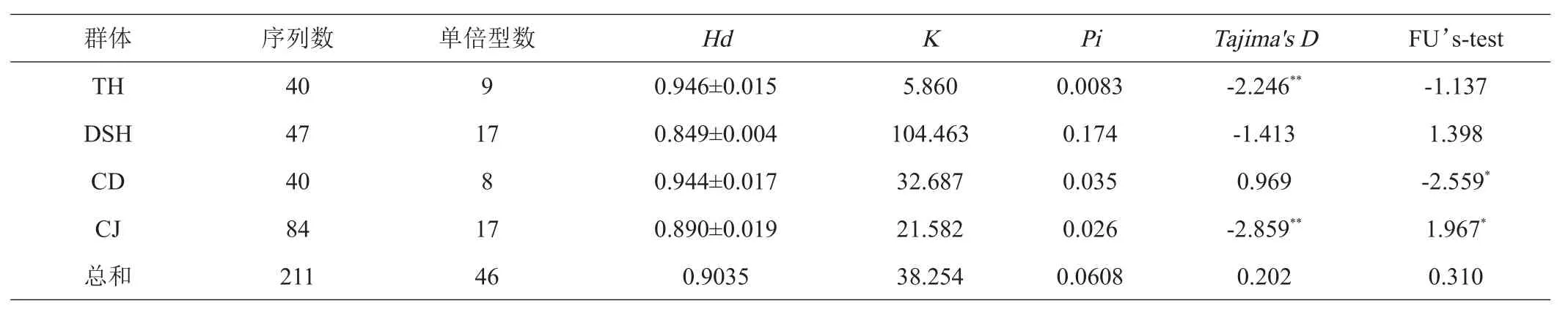
表1 不同群体的单倍型及遗传多样性参数
2.2 单倍型分布
在211个翘嘴鲌个体共有46个单倍型,其中Hap1、Hap2、Hap3、Hap4和Hap8在长江和长荡湖群体中共享,其余41个单倍型分别在4个地理群体中独有,占单倍型的89.13%,其中Hap5、Hap6和Hap10-Hap19在长江群体中特有,Hap20-Hap34、Hap45及Hap46在淀山湖群体中独有,Hap7、Hap9和Hap40在长荡湖中特有,Hap35-Hap39和Hap41-Hap44在太湖群体中独有。
通过Network软件绘制单倍型网络图,如图1所示,每个圆代表一个单倍型,其大小与总频率成正比,圆圈大小表示该单倍型出现的次数。每个单倍型按地理分布用不同的颜色表示,4个地理群体的单倍型构成一个网状结构图,只有长江群体的长荡湖群体有共享单倍型,长江群体和淀山湖群体的单倍型散布在各个位置,其余群体的单倍型分布较均匀。图中圆的面积代表出现次数,由图可知,Hap2出现次数最多为54次,包含长江群体和长荡湖群体,其次是Hap41,出现23次,属于太湖群体。Hap14出现22次,属于长江群体。

图1 4个地理群体翘嘴鲌的单倍型分布网络图
2.3 遗传分化分析
由MEGA7.0.26软件中的Data-distance计算出的4个地理群体的遗传距离如表2所示,在表中可看出种内遗传距离的范围是0.01~0.04,其中太湖群体的种内遗传距离是0.01,长荡湖群体的种内遗传距离最大,达到0.04。4个群体的种间遗传距离0.052~0.152,太湖群体和淀山湖群体的种间遗传距离最远是0.152。
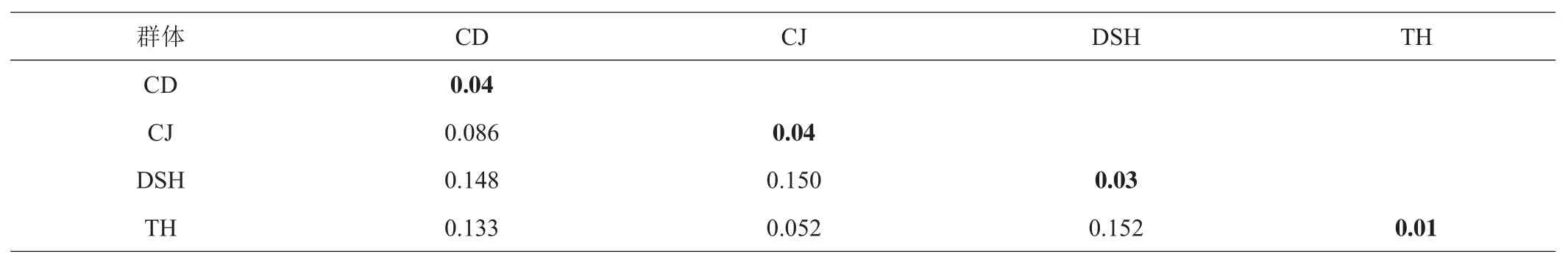
表2 不同翘嘴鲌群体遗传距离
4个地理的翘嘴鲌群体基因流和遗传分化指数如表3和4所示,从表中显示出两两群体间的遗传分化指数范围在0.197~0.646。基因流值的范围为0.152~1.010,其中长江和淀山湖群体的基因流值最小为0.152,长江与长荡湖群体的基因流值最大为1.019。

表3 不同翘嘴鲌群体基因流(Nm)及遗传分化指数(Fst)

表4 4个翘嘴鲌地理群体间遗传分化参数
2.4 聚类分析
应用MEGA7.0.26软件对211个翘嘴鲌个体的Cyt b的序列进行聚类分析,得出4个翘嘴鲌群体46个单倍型的系统进化树如图2所示。不同地理来源的单倍型总共分成7个大分支,Hap25、Hap28、Hap31和Hap33独自各成一支,属于太湖群体。Hap30和Hap34组成一支,亦属于太湖群体,Hap45独自为一支,属于淀山湖群体,剩余一支包含很多单倍型,其中不同地理群体的单倍型散乱分布。

图2 4个地理群体翘嘴鲌的单倍型NJ系统发育树
2.5 4个翘嘴鲌群体错配分布图
用DNASPv5软件分析得到如图3所示的错配分布图,由图3可知,当把4个水域的翘嘴鲌群体作为一个整体分析时,其歧点分布图大体呈现为单峰。
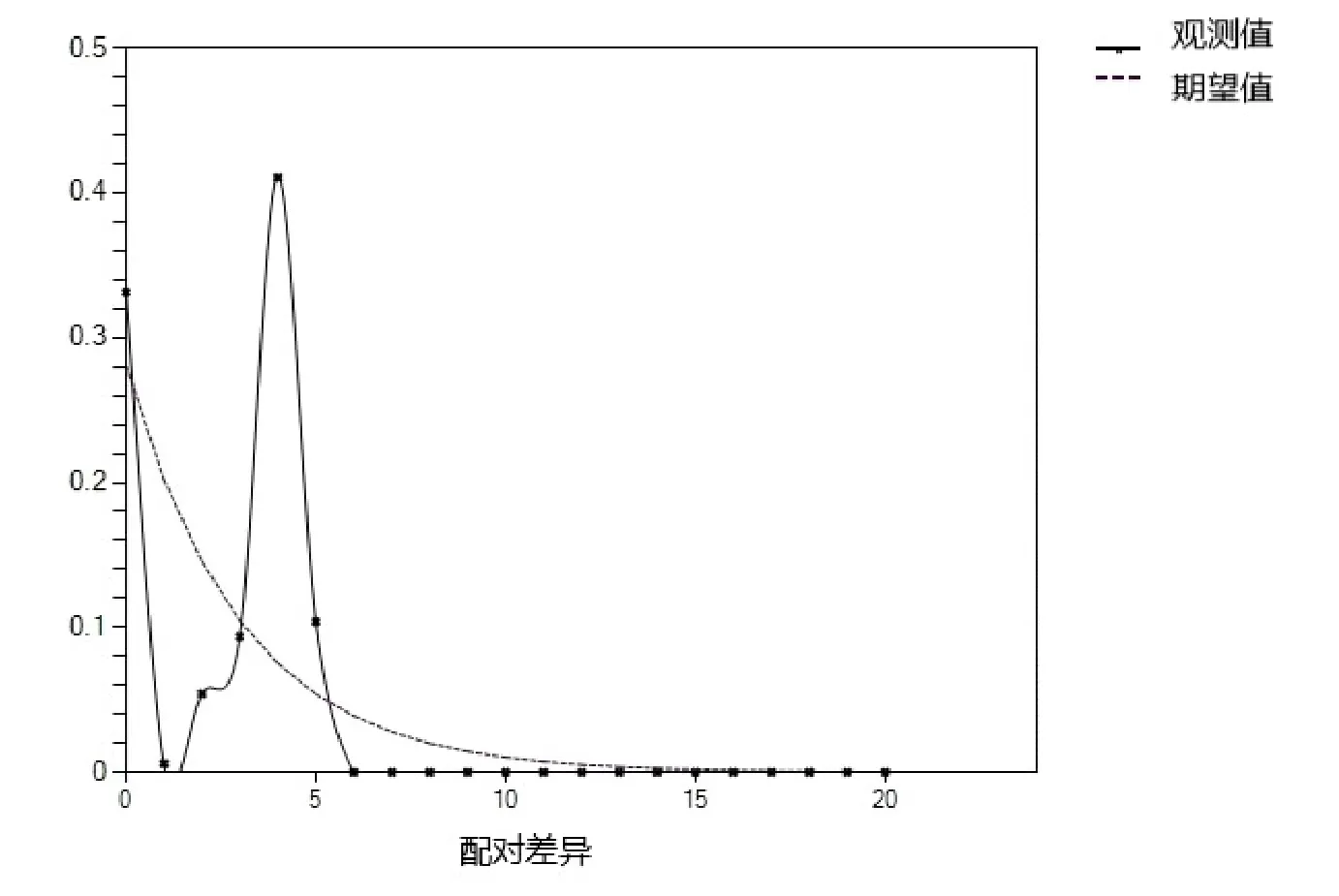
图3 错配分布曲线
3 讨论
3.1 序列组成特点
本研究中211个个体序列的碱基组成平均含量为T(27.1%),C(23.1%),A(27.9%)和 G(21.9%),A+T 的含量占55%,G+C占45%,表现出A+T的含量高于G+C的含量。与张久盘[21]文中A+T含量为56.8%,G+C含量为43.2%;冯晓宇[22]文中的A+T含量为56.6%,G+C含量为43.4%,研究结果相一致,均表现出碱基偏倚性,符合大多数硬骨鱼的特征[22]。
3.2 翘嘴鲌地理群体遗传多样性
遗传多样性是生物进化和适应环境的基础,黄小彧[11]等对线粒体DNA序列遗传变异的研究表明,若单倍型多样度(Hd)高于0.5且核苷酸多样度(Pi)低于0.005,是受瓶颈效应后群体快速扩张导致,表明该群体的遗传多样性水平低;如果单倍型多样度(Hd)高于0.5且核苷酸多样度(Pi)高于0.005,则该群体遗传多样性水平较高[23]。本研究中整个翘嘴鲌群体的遗传多样性水平较高,遗传多样性丰富,与徐丹丹[24](Hd:0.948,Pi:0.017),徐宇[25](Hd:0.969,Pi:0.2008)和张久盘[21](Hd:0.998,Pi:0.039)的研究结果一致。本研究中长江群体表现出高的单倍型多样性和高的核苷酸多样性,与黄小彧等[11]的结果(Hd:0.866,Pi:0.00330)存在差异,可能是黄小彧等研究中使用的是线粒体控制区,控制区是线粒体DNA的高突变区;还可能是近几年长江的水域鱼类生境优化和资源恢复措施导致长江翘嘴鲌的遗传多样性较高。
遗传多样性的高低通常由遗传距离的大小来反映,种内个体间遗传距离小,彼此间亲缘关系近,说明遗传多样性低,反之,遗传距离大则遗传多样性高[17]。本研究中4个地理群体的种内遗传距离的范围是0.01~0.04,其中长江群体和长荡湖群体的种内遗传距离相同且最大,遗传多样性较高,太湖群体的种内遗传距离最小,遗传多样性也较低,与王伟等[17]的研究结果一致。通常情况下,地理距离越远,群体间遗传距离也就越大,而徐丹丹等[24]在基于微卫星标记和线粒体Cyt b基因序列针对鲇(Silurus asotus)遗传多样性的研究一文中,发现不同鲇群体遗传距离与其地理距离之间没有相关性;沙航等[26]基于COI序列分析长江中上游6个鲢(Hypophthalmichthys molitrix)地理群体遗传多样性也斧正了遗传距离的大小与群体地理距离没有明显相关性。本研究中淀山湖距离长荡湖比距离太湖远,但淀山湖群体与长荡湖群体的遗传距离小于淀山湖群体与太湖群体的遗传距离,这3个湖泊的翘嘴鲌群体之间遗传距离与其分布的地理距离亦没有相关性,这一结果与上述研究结果类似。
群体遗传学中通常以Fst值的大小作为衡量群体间遗传分化程度的常见指标,Fst<0.05表示群体间是低度分化;0.05<Fst<0.15 表示群体间是中度分化;0.15<Fst<0.25表示群体间是高度分化;0.25<Fst<1表示群体间有极大的遗传分化[27]。本研究中长江翘嘴鲌群体和长荡湖群体的Fst相对较小,为0.197,属于高度分化,长江群体、淀山湖群体和太湖群体间的Fst值较大,属于极大分化,这有可能是研究水域存在地理隔离,翘嘴鲌群体间不存在基因交流。群体间平均净遗传距离(Da)是衡量群体间分化程度的重要指标,值越大,群体间的遗传分化程度越大[24,28]。长荡湖和太湖群体的平均净遗传距离相对其它群体之间较大(0.48),遗传分化程度高,与其遗传分化指数较高的研究结果相一致。本研究中4个群体之间的基因流值都小于4,研究结果进一步说明了4个翘嘴鲌群体之间基因交流较少,存在一定程度的遗传分化现象,这与陈会娟等[29]的结果一致。
3.3 翘嘴鲌群体历史动态
一般要判断某一群体有没有发生过群体扩张,有两种方法:中性检测和歧点分布分析[11,30-31]。本研究用这两种方法来分析了翘嘴鲌群体的种群历史动态情况。当检测数值为负值,且具有显著意义时,则认为该群体曾发生过群体扩张现象,如果群体发生过扩张,则歧点分布图呈现出单峰;如果群体未经历过扩张,呈现稳定状态,则其歧点分布图呈现为多峰[11]。对211个翘嘴鲌个体的Cyt b中性检验得出Tajima's D值,除了长荡湖群体,其余群体都为负值,且长江和太湖群体的Tajima's D表现出极显著性差异P<0.01,表明群体在历史上曾发生过群体扩张。淀山湖群体的Tajima's D为负值,但不具有显著性差异。除此之外,歧点分布图显示,4个翘嘴鲌地理群体的错配分布曲线中,将所有翘嘴鲌群体作为一个整体分析,错配曲线呈现单峰,说明该翘嘴鲌群体发生过群体扩张,这与张力文等[23]的结果相一致。
4 结论
本研究对4个水域的翘嘴鲌进行遗传多样性和遗传结构的分析,采样点覆盖长江下游典型水域,样本较多。结果显示4个地理水域的翘嘴鲌群体,表现出高的单倍型和高的核苷酸多样性,遗传多样性较为丰富,群体间都存在遗传分化,分化程度不同。研究表明,线粒体DNA Cyt b可以作为评估翘嘴鲌的遗传多样性和种质资源现状的有效分子标记,研究结果为翘嘴鲌种质资源评估和保护提供了科学的数据支撑。后续研究将继续增加野生翘嘴鲌群体的样品数量和研究水域,采用多种分子标记方法,全面而系统的评估长江下游各个水域翘嘴鲌的种质资源状况和分析其遗传结构。
猜你喜欢
美食(2017年9期)2017-09-15 08:52:47
玩具世界(2017年4期)2017-07-21 13:27:24
宝藏(2017年2期)2017-03-20 13:16:42
中亚信息(2016年3期)2016-12-01 06:08:24
中国诗歌(2016年6期)2016-11-25 23:54:04
金融经济(2015年16期)2015-08-21 11:23:58
现代农业科技(2014年13期)2014-09-18 04:00:38
现代农业科技(2014年7期)2014-07-07 12:36:00
城市建设理论研究(2014年11期)2014-04-21 00:36:00
华东师范大学学报(自然科学版)(2014年6期)2014-02-27 13:40:57
