
塔里木河上游荒漠河岸胡杨林树洞型空心树发生过程与形成机制
2021-09-02 01:28:14周洪华朱成刚方功焕
生态学报 2021年14期
周洪华,朱成刚,方功焕
中国科学院新疆生态与地理研究所,荒漠与绿洲生态国家重点实验室, 乌鲁木齐 830011
空心树在森林中非常常见[1- 2]:亚马孙热带雨林中的树木有37%的空心率;澳大利亚稀疏草原上66%—89%的物种会发生空心,甚至有的树干空心直径达到整棵树木直径的50%以上[3]。空心树是森林生态系统的重要组成部分,它在维持森林的动物或微生物物种多样性方面起着重要作用[1,4]。因此,空心树生态学研究已在世界多国和地区广泛开展,成为了森林生态系统管理以及生物多样性保护的重要研究内容之一[2-3]。当前对空心树的研究主要集中在各种森林生态系统中空心树的数量和组成[5- 7]、空心树调查方法[8-9]、空心树与森林管理[10-11]、树洞依赖动物对空心树的选择利用[12]、空心树对生态系统的生态意义[2,3,13]、空心树的成因探讨[1,14-16]等各方面。纵观国内外空心树研究,澳大利亚的研究最为详尽,但研究物种较为单一,大多以桉树为研究对象[1,13,15,17-18],其次为北美地区[2,19]、欧洲[11,20-22]等地。然而,我国空心树研究几乎一片空白,仅在西双版纳热带森林、哀牢山亚热带森林、吉林次生林和额济纳旗荒漠河岸林等地有关于空心树的零星研究[23-27]。
荒漠河岸林是干旱荒漠区森林生态系统的主体,是干旱荒漠区最具代表性的非地带性植被类型和珍贵的森林资源,也是干旱区生态系统重要的组成部分和内陆河流域生态系统的主要生产者,在生态平衡维持、荒漠化防治、生物多样性保护及绿洲农牧业的天然保障等方面有着重要的生态意义[28-30]。胡杨是我国西北干旱区荒漠河岸林的唯一乔木建群种,它作为世界林木中古老而珍贵、濒临灭绝的物种,因兼具耐旱、耐盐、耐风沙的特性,广泛分布在从摩洛哥西部到北非、西亚、南亚、再到中亚以及蒙古,在印度、巴基斯坦、哈萨克斯坦、蒙古、伊朗和中国的干旱荒漠区都能见到它们傲立的身影[31-32]。胡杨也是荒漠森林系统中树干空心的典型树种[33-35],然而,目前胡杨空心树的数量特征和分布格局并不清楚。
位于西北干旱区的塔里木河流域是世界胡杨林存林面积最多、林相保存最完好的区域,占世界胡杨林面积约60%,占我国胡杨林面积约90%,堪称世界上最大的胡杨林基因库[36],是研究胡杨空心树生态学的理想场所。因此,本研究以塔里木河流域为研究靶区,调查研究胡杨空心树的数量特征与分布格局,并试图初探胡杨空心树的发生与形成过程,以补充和完善空心树生态学的理论基础,并为干旱区荒漠河岸林的生态系统维系提供科学支撑。
1 材料与方法
1.1 研究区概况
塔里木河是我国最大的内陆河,全长1321 km,其水系由“九源一干”144条河流构成。目前,构成塔里木河的9源流中,与塔里木河干流保持有自然地表水力联系的仅有阿克苏河、和田河以及叶尔羌河:阿克苏河与塔里木河干流保持常年的地表水力联系,是塔里木河的主要补给水源;和田河和叶尔羌河仅在丰水期短暂补给干流,其余时间均为断流状态[37]。阿克苏河、和田河和叶尔羌河三河在阿克苏地区阿拉尔市的肖夹克汇合,形成塔里木河干流的始端。塔里木河干流分为上中下游三段,其中从阿拉尔市的肖夹克至轮台县的英巴扎为上游,从英巴扎到尉犁县的恰拉为中游,从恰拉到若羌县的台特玛湖为下游。塔里木河流域属大陆性暖温带荒漠干旱气候,年均太阳总辐射 5692—6360MJ/m2,年均日照时数 2780—2980 h,≥10 ℃ 年积温4040—4300 ℃,年均降水量 17.4—42.0 mm,年均潜在蒸发量2500—3000 mm,是降水量的 40 倍;土壤类型主要为含盐量较高的沙土;天然植被主要包括胡杨(Populuseuphratica)、柽柳(Tamarixspp.) 、黑果枸杞(Lyciumruthenicum) 、骆驼刺(Alhagiparsifolia) 、花花柴(Kareliniacaspia) 、 芦苇(Phragmitescommunis) 、罗布麻(Poacynumhendersonii) 和河西菊(Hexiniapolydichotoma) 等[34]。
由于河道南北摆动、迁徙无定,塔里木河作为“一匹无僵的野马”奔腾咆哮着穿行在荒漠之上,孕育着面积广阔的胡杨林。沿塔里木河干流河道,生长着一条带状、宽数公里的荒漠河岸林,牢牢地挡在塔克拉玛干沙漠与库姆塔格沙漠中间,成为阻挡沙漠入侵绿洲的 “排头兵”、守护南疆家园的“绿色走廊”,对维系生态系统的稳定起着极其重要的作用。因其兼具抗旱、抗盐碱和抗风沙等特点,胡杨是这一绿色走廊中最具特色的建群乔木物种。根据塔里木河分段,以胡杨为优势建群种的荒漠河岸林也分为上、中、下游三段。由于塔里木河上中游地表水被大量开发,用于工农业发展,塔里木河下游荒漠河岸林因河道断流、地下水位下降等原因大面积衰退[34-35],上中游荒漠河岸林保存相对完整,尤其是上游。本研究选择林相完整、存活完好的上游荒漠河岸林胡杨为研究对象,调查胡杨空心树数量组成与分布格局,具体研究区位于塔里木河三河汇合口(肖夹克)的塔河源林区。
1.2 胡杨空心树调查方法
在肖夹克河岸林区,胡杨幼林主要分布在垂直河道0—1.0 km范围内、成熟林和过熟林主要分布在垂直河道1—1.5 km范围内。根据胡杨生长状态,分别在幼林区和成熟林区随机设置1个30 m(水平于河道)×200 m(垂直于河道)的调查样地,将各样地内的所有存活的胡杨进行编号、挂牌,并将每个样地内胸径大于5 cm 的活体胡杨均作为调查对象;幼林样区内合计胡杨892株,成熟林样区合计胡杨104株。参考国内外空心树的调查方法,采用目视+望远镜的地面观察法界定树洞型空心树。本研究中把至少含有一个明显可见的树洞且洞口直径(洞口垂直高度和水平宽度中取最小者)≥5cm 的都定义为树洞型空心树。对于在地面可以测量到树洞的直接测量树洞的大小,而对于分布在树干或树枝高处无法直接测量到的树洞则采用望远镜(10×25)从各个方位对整株乔木的结构进行观察,确定树洞所处的位置并估测树洞的大小[8,10];同时记录树洞距离地面高度、在树干中的位置、树洞朝向以及样方内树木数量和每棵树的胸径。
2 结果与讨论
2.1 空心树数量特征与分布格局分析
肖夹克河源林区幼林的树木密度较高,平均密度为0.15棵/m2;平均胸径为10.74cm。幼林中胡杨空心树的出现频率较低,仅占样地全部树木数量的10.34%,有89.66%的胡杨为非空心树;幼林树洞较小,平均为5.80cm(水平宽度)×6.40cm(垂直高度)。成熟林的树木密度很低,平均密度为0.02棵/m2;平均胸径为36.07cm。成熟林中胡杨空心树的出现频率极高,占样地全部树木数量的83.78%,仅有16.22%的胡杨为非空心树;树洞大小平均为13.10cm(水平宽度)×16.00cm(垂直高度)。由此可看出,空心树出现的几率随树龄增加而增加。
根据实地调查结果,可按树洞所处位置将树洞类型分为两大类:主干树洞型空心树和分枝树洞型空心树。依据胡杨树洞的发生位置,主干树洞型空心树可细分为主干基部树洞、主干中部树洞、主干劈裂树洞、主干分叉树洞、主干小枝断口树洞;分枝树洞型空心树可细分为分枝断口树洞、分枝中部树洞(图1)。
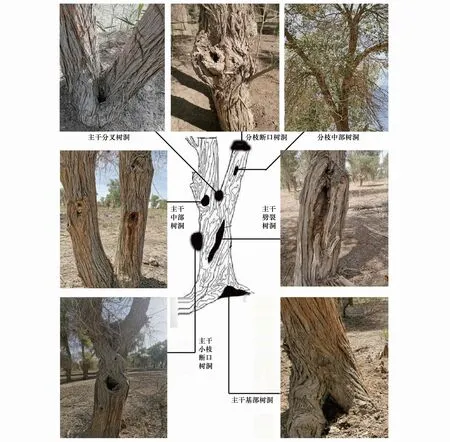
图1 塔里木河上游胡杨林树洞型空心树分布类型Fig.1 Types of hollow-bearing trees of Populus euphratica in the upper Tarim River
塔里木河上游肖夹克胡杨林各类型空心树所占比例显示(表1),主干树洞型空心树占空心树类型的74.20%,分枝树洞型空心树占空心树类型的25.80%,其中因主干小枝或大分枝折断后从断口处形成树洞的空心树是胡杨空心树形成的主要类型,占空心树的61.29%。

表1 塔里木河上游胡杨林树洞型空心树类型占比
塔里木河上游肖夹克胡杨林空心树的树洞朝向并不固定,东南西北向均有树洞出现。其中,朝向为北-东的树洞占树洞比例为43.24%、朝东-南的树洞为27.03%、朝南-西的树洞为18.92%、朝西-北的树洞为10.81%。因此,肖夹克林区胡杨空心树树洞朝向集中在树干的东北面、东南面和西南面,其中东北面是树洞出现的最主要朝向位置,此朝向树洞约占所有树洞朝向的约50%。
2.2 空心树树洞发生与形成过程初探
随着树龄的增加,空心树发生几率增加,“树大心空”似乎成了“约定俗成”的自然现象,可树木为何会空心呢?早在1976 年,生态学家Janzen认为空心现象是树木对生长环境采取的一种适应性策略,提出树木空心是为了进行更有效地循环利用营养,即受外界侵扰后,树木内心慢慢腐烂,无用的营养物质逐步流到植物根部,被循环利用,从而使树干形成空心[16]。但Janzen的这一“无用之用的能量循环理论”一直未得到证实。近年来,英国科学家Ruxton否定了营养循环利用理论,认为树木空心是为了选择性地舍“内”求“外”,通过牺牲一部分化学防御来节省能量,即树心一旦形成后,它既不储存能量也不传输能量,使得化学防御物质无法得到更新,因此抵抗能力降低,从而更易受到外界环境的侵蚀,进而造成树木空心[14]。但同样,这个理论也备受争议,未得到证实。因而,树木空心的发生过程和形成机制一直在困扰着生物学家们,目前仍是一个悬而未决的科学问题。
本研究调查分析表明,肖夹克林区胡杨空心树树洞类型主要为主干小枝或者分枝断裂后形成的,这可能是因为小枝或分枝断裂形成“伤口”后,“伤口”周边的韧皮部里的营养物和周边木质部里的水分慢慢从“伤口”中分泌出来,这些分泌物在微生物和蚂蚁等昆虫的作用下逐渐“消融和腐蚀”木质部,使得“伤口”不断横向和纵向发展,从而形成了树洞。野外调查也证实,不同形成时期的树洞外观不同,小的、新鲜的树洞在洞口和洞内一般都有湿润的分泌物析出,且在树洞中有较为活跃的微生物和昆虫,大的、陈旧的树洞一般洞口和洞内都比较干燥,不再有明显的湿润分泌物析出。因此,大多数胡杨树洞形成都符合“外伤-分泌物外泄-木质部被腐蚀、消融-伤口扩大-形成树洞”这一过程(图2)。由此可见,胡杨树洞的形成是始于外伤,即外界侵扰造成伤口,然后使得营养物和水分等分泌物从伤口流出,在微生物等作用下木质部逐渐被腐烂,腐烂的物质逐步通过伤口流到树木根部供植物再次吸收利用。因此,胡杨空心树树洞的形成过程基本符合生态学家Janzen关于空心树的“无用之用的能量循环理论”形成机制。但是值得注意的是,由于塔里木河流域降水稀少,相对湿度较低,干燥的环境下,部分腐烂物质从伤口流出后,在流经树干的过程中,逐渐失去水分,并停止流动,仅在树洞周边和下方留下腐烂物流经的痕迹,这可能就是为什么一些离地较高或陈旧的树洞比较干燥的原因。

图2 塔里木河上游胡杨林树洞形成过程Fig.2 Formation process of hollow-bearing trees of Populus euphratica in the upper Tarim River
塔里木河沿线天然胡杨林生长在沿河荒漠土壤,其生长受人为扰动相对较少,主要受自然环境和水资源影响。荒漠区动物种类不多,但风沙较为频繁,树木小枝或分枝易受风沙侵扰,容易折断,尤其是在秋、冬季树枝相对干枯的时节。肖夹克所属的阿拉尔市气象局2005—2018年的14年间日最大风速风向显示(图3),阿拉尔市风向多年均值中北/北东/东北向风占比最高,为所有风向的43.45%;其次为西/西北/北西向风,占所有风向的24.38%;再次为南/西南/南西向风,占比21.82%;东/东南/南东向风占比最小,为10.35%。多年季节风向变化上,春秋季北/北东/东北向风和西/西北/北西向风频率较高,分别为49.34%和31.91%;秋冬季北/北东/东北向风和南/西南/南西向风出现频率较高,分别为38.04%和31.44%。这些高频率风沙风向与树洞在树干中所处的主要朝向较为一致。因此,可以推断,胡杨树木的外伤可能主要来源于长期的风沙侵扰。风沙侵扰造成树木小枝或分枝折断,从而造成树木出现不同程度的“伤口”。
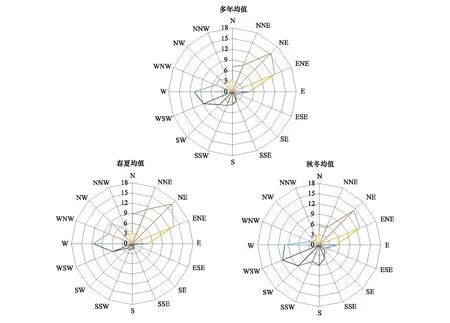
图3 塔里木河上游阿拉尔2005—2018年风向解析Fig.3 Wind direction in 2005—2018 in Alaer of Tarim River Basin
3 结论与展望
塔里木河上游肖夹克荒漠林区胡杨树洞型空心树发生频率与树龄有关,随树龄的增加而增加,幼林空心树发生频率较低,成熟林空心树发生率极高,达到了林区树木的80%以上。空心树树洞可分为主干型和分枝型两大类,主干型树洞占绝对优势;主干小枝断口造成的树洞和大分枝断口造成的树洞是树洞形成的两种主要表现方式。树洞主要始于长期风沙侵扰造成树干小枝或分枝断裂形成的外伤,“伤口”周围韧皮部、木质部内水分和营养物质等外泄,这些分泌物在化学过程和微生物等作用下使得韧皮部和木质部不断被腐蚀、消融和分解,最终形成了树洞,其发生过程和形成机制基本符合“无用之用的能量循环理论”。
当然,树木空心的发生过程和形成机制非常复杂,涉及一系列的自然环境、微生物、昆虫、动物以及化学作用等作用过程。本研究仅对林区树洞现状进行了调查,并对其可能的形成过程进行了初步剖析和推测,其更深入的发生过程和形成机制还有待于更多的生态学家们共同努力,进一步详细探究。同时,我们在野外调查过程中也相继采集了树洞内分泌物样品、主干基部树洞内外土壤样品以及空心树根际土壤水和地下水等样品,未来准备进一步分析树洞分泌物中微生物特征及胡杨木质部在分泌物、根际土壤和地下水作用下的分解特征,以期深入探讨和揭示胡杨树洞型空心树的发生过程与形成机制,进一步完善胡杨空心树生态学理论基础。
猜你喜欢
公民与法治(2022年10期)2022-10-12 07:46:54
小哥白尼(军事科学)(2022年5期)2022-08-17 08:14:10
小哥白尼(军事科学)(2022年2期)2022-05-25 13:19:28
Chinese Physics B(2021年2期)2021-03-11 08:31:50
少儿美术(快乐历史地理)(2019年3期)2019-07-23 01:21:24
意林绘阅读(2019年2期)2019-03-05 19:56:34
下一代英才(2018年12期)2018-02-20 14:28:06
Coco薇(2015年12期)2015-12-10 02:46:23
天文爱好者(2015年12期)2015-08-23 11:54:32
吐鲁番(2014年2期)2014-02-28 16:54:39
