
链格孢霉菌侵染番茄产毒机制
2021-08-31 03:30曲劲尧林煜程卢国柱袁智鹏张瑶瑶尤艳莉李彦伸
食品科学 2021年15期
曲劲尧,林煜程,毛 馨,卢国柱,袁智鹏,张瑶瑶,尤艳莉,李彦伸*
(烟台大学生命科学学院,山东 烟台 264005)
链格孢霉属(Alternariaspp.)是一种广泛分布于自然界的真菌,属于暗梗孢科砖隔孢亚科[1-2],包括腐生菌和内生菌,是多种谷物、水果和蔬菜的致病菌[3]。由于其在低温下也能生存,因此普通的低温冷链技术运输和贮存方式仍然避免不了由其引起的农产品的腐烂,造成巨大的经济损失[4]。除此之外,链格孢霉可产生约70 种有毒代谢产物,如已经在多种食品中检测到的链格孢酚(alternariol,AOH)、链格孢甲基醚(alternariol monomethyl ether,AME)、细链格孢菌酮酸(tenuazonic acid,TeA)、链格孢霉烯(altenuene,ALT)、腾毒素(tentoxin,TEN)[5-6]。据报道,许多链格孢霉毒素具有细胞毒性、基因毒性、致突变性[7-10]。此外,大量研究发现,在一定条件下,真菌毒素分子可与糖、氨基酸或硫酸盐等极性较强的物质结合,生成结合型真菌毒素[11]。这些毒素在代谢过程中一经水解就会释放毒素本体,成为危害人类健康的潜在风险之一[12]。
液相色谱-质谱联用已成为多种真菌毒素准确分析的首选技术[13],但由于商业化标准品的缺乏,隐蔽型真菌毒素在常规检测中往往被忽视,再者百万级的质谱数据无疑增加了数据分析和挖掘有用信息的艰难程度[14]。利用高分辨质谱和后续数据分析软件进行非靶向代谢组学分析为隐蔽型真菌毒素的检测和天然化学产物的研究提供了新的解决方案[15]。
首先,开源性软件MZmine的数据处理模块利用嵌入式可视化工具,允许直接预览分析结果,经峰值检测、去同位素等处理后,可去除大量无用的峰,从而得到更有价值的分析结果,适合大批量的数据分析[16]。其次,Xcalibur软件可对全扫描、二级质谱扫描、同位素峰、公式预测、保留时间进行精准分析。最后,全球天然产物社会分子网络(global natural products social molecular networking,GNPS)是一个基于网络的质谱生态系统,旨在成为一个开放获取的知识库,供社区组织共享、加工和鉴定串联质谱原始数据[17-18]。基于特征的分子网络(feature based molecular networking,FBMN)可以对质谱碎片数据和评估结构相似性之间的代谢物进行可视化分析,对于化合物的快速鉴定及新型化合物发现具有重要的作用[14,19-21]。Quinn等[22]介绍了分子网络的应用范围,并认为其可作为创新工具用于药物发现、临床应用和精准医疗。Xu Shanshan等[23]使用基于高分辨质谱的 代谢组学和GNPS-FBMN评估了在湿热或干冷环境中存储1~9 年的普洱茶,结果表明,普洱茶的化学特征在早期相似,但在湿热环境中的第5年和干冷环境中的第7年,其化学特征开始相互区分。
中国是世界上主要的番茄生产国、消费国、贸易国[24]。由于番茄果皮很薄,所以容易受到真菌的感染,而链格孢霉是发霉番茄上最常见的真菌[25]。本研究以番茄为实验材料,模拟室温(25 ℃)和低温(4 ℃)贮藏过程中,番茄被链格孢霉感染的过程,对不同感染时期链格孢霉毒素及隐蔽型真菌毒素进行定性和代谢通路分析,为番茄链格孢霉污染防控和链格孢霉毒素代谢分析提供理论依据和数据支撑。
1 材料与方法
1.1 材料、菌株与试剂
成熟度相同的番茄(每个番茄质量约200 g,无机械损伤和病害、颜色均匀)购自山东省烟台市莱山区烟大市场;链格孢霉M1-2为本实验室保存菌株。
链格孢霉毒素标准品(AOH、AME、TeA、TEN、ALT) 青岛普瑞邦生物工程有限公司;马铃薯葡萄糖琼脂培养基 北京路桥技术有限责任公司;NaCl(分析纯)、乙腈(分析纯)、乙醇(分析纯) 国药集团化学试剂有限公司。
1.2 仪器与设备
SHH-250L生化培养箱 重庆市永生实验仪器厂;SW-CJ-2DX型双人单面超净工作台 苏州净化设备有限公司;D-1-70型自动控制压力蒸汽灭菌锅 北京发恩科贸有限公司;3k15高速台式冷冻离心机 德国SIGMA 公司;MP2002电子天平 上海市舜宇恒平科学仪器;血球计数板 上海市求精生化试剂仪器有限公司;M150显微镜 麦克奥迪实业集团有限公司;Q Exactive plus组合型四极杆Orbitrap质谱仪 赛默飞世尔科技 (中国)有限公司。
1.3 方法
1.3.1 分生孢子悬液的制备
将链格孢霉供试菌株接种于马铃薯葡萄糖琼脂培养基,在25 ℃下光暗交替各12 h培养7 d。吸取2 000 μL无菌生理盐水于培养真菌的平板上,用接种环轻轻刮取平板 表面的孢子,吸取平板上含有分生孢子的生理盐水并转移至50 mL离心管内。继续向平板上加入1 000 μL无菌生理盐水,将平板表面残留的孢子洗下,合并含有孢子的生理盐水于50 mL离心管中,并用无菌生理盐水定容至10 mL,充分漩涡后用4 层灭菌擦镜纸过滤,以除去菌丝。使用血球计数板在显微镜下计数后,用无菌生理盐水将分生孢子悬液浓度调整至1×105个/mL,置于4 ℃下保存备用。
1.3.2 番茄样品处理及毒素提取
选取成熟度相同的番茄果实36 个,浸泡在75%(体积分数,后同)乙醇溶液中表面消毒2 min。在超净台中用无菌去离子水冲洗3 次后晾干。在无菌环境下进行接种:用无菌牙签在果实赤道处刺1 个约5 mm深度的伤口,吸取5 μL孢子悬液接种于刺好的伤口处;接种相同体积的无菌生理盐水作为对照组。接种后的番茄果实放于塑料筐(已用75%乙醇溶液消毒)中,用保鲜膜密封,分别放置于4 ℃和25 ℃下贮藏。分别在接种后5、10、15 d进行取样,每次取3 个平行。无菌条件下,取病斑腐烂处样品1.00 g于10 mL离心管内,加入5 mL 84%乙腈溶液。漩涡振荡混匀后,10 000 r/min离心10 min,取上清液经0.22 μm有机滤膜过滤,滤液进行超高效液相色谱-质谱分析。
1.3.3 液相色谱-质谱条件
液相色谱条件:Hypersil GOLD™ C18色谱柱(100 mm×2.1 mm,1.9 μm);柱温:25 ℃;流动相:A相为0.05%(质量分数)氨水,B相为0.1%甲酸-乙腈溶液;梯度洗脱程序:0.0~1.0 min,5% B;1.0~3.0 min,5~30% B;3.0~4.0 min,30~40% B;4.0~5.5 min,40~55% B;5.5~7.5 min,55~95% B;7.5~9.0 min,95~5% B;9.0~10.0 min,5% B;进样量:5 μL;流速:0.3 mL/min。
质谱条件:离子源参数:鞘气流速:45 L/min;辅助气流速:10 L/min;挡锥气流速:0 L/min;电喷雾电压:3.5 kV;离子导管温度:320 ℃;离子源温度:350 ℃;采集的模式为负离子模式下的一级全扫描和数据依赖性二级质谱扫描dd-MS2。
一级全扫描的具体参数:分辨率:70 000;离子注入数目:3×106;最大离子注入时间:250 ms;扫描范围:120~900m/z。
数据依赖性二级质谱扫描dd-MS2的具体参数:分辨率:17 500;离子注入数目:1×105;最大离子注入时间:64 ms;循环次数:5 次;隔离窗口:1.2m/z;归一化碰撞能量:20、40、60 eV。dd参数:注入离子数目:1×103;顶点激发:2~6 s;同位素峰排除:开;动态排除:8.0 s。
1.3.4 分析工具及参数设定
使用MZmine 2软件的碎片过滤诊断对偶联产物进行 筛选,本研究中设置诊断中性丢失值为m/z79.956 8,m/z公差(tolerance)为0.02或5×10-6,基峰设置为20%。使用MZmine 2软件的质谱扫描和色谱峰建立功能进行过滤和建立色谱峰;使用色谱峰的分离和同位素峰去除功能进行色谱峰的分离和去同位素峰;使用列表对齐功能使所有数据特征峰根据保留时间和特征列表进行对齐;使用自定义数据库对化合物进行鉴定。最后,导出包含完整一级质谱和二级质谱的特征文件用于GNPS分析。
将MZmine 2软件导出的特征文件上传至GNPS平台,使用FBMN工具进行分析。设置母离子质量误差和子离子碎片质量误差均为m/z0.02。使用边缘过滤,使其余弦值大于0.7并且匹配的峰超过6 个,建立分子网络。导出分子网络数据文件,在Cytoscape软件中进行可视化分析。
1.4 数据分析
利用Xcalibur软件对5 种已有商业标准品的链格孢毒素(AOH、AME、TeA、ALT、TEN)的保留时间、母离子、子离子等相关信息进行确定,样品采用相同的仪器条件进行分析,研究3 次取样中链格孢霉毒素的产生情况。对质谱采集后的数据文件使用MZmine 2软件的碎片过滤诊断功能,对所有硫酸盐结合的二项代谢物进行筛查研究。在MZmine 2软件筛查结果的基础上,采用Xcalibur软件对筛查后的结果进行数据分析,基于保留时间(硫酸盐结合产物极性增强,保留时间提前)、母离子和子离子碎片(脱SO3的质量碎片)、精确质量数、中性丢失值(m/z79.9568)、同位素峰(含S同位素的1.995质量位移)等进行确证。利用Mzmine软件中自定义数据库筛查功能,对链格孢霉毒素筛选结果建立热图,对各个毒素的含量随时间的变化进行可视化分析。最后借助GNPS平台的基于特征的分子网络功能构建分子代谢网络,对链格孢霉毒素代谢通路进行分析。
2 结果与分析
2.1 接种后番茄果实在不同贮藏条件下的外观变化
人工接种链格孢霉分生孢子后,番茄果实在不同贮藏条件下的外观变化如图1所示。25 ℃条件下贮藏5 d时,刺伤接种处开始出现黑色病斑,随着贮藏时间延长,病斑直径也增大,10~15 d病斑增大速度加快,15 d后番茄底部开始出现自发的腐烂。4 ℃下贮藏15 d的过程中,番茄果实表皮逐渐褶皱,但刺伤接种处无黑色病斑出现。结果表明,低温抑制了链格孢霉的生长和产毒,能够起到有效的保鲜作用。
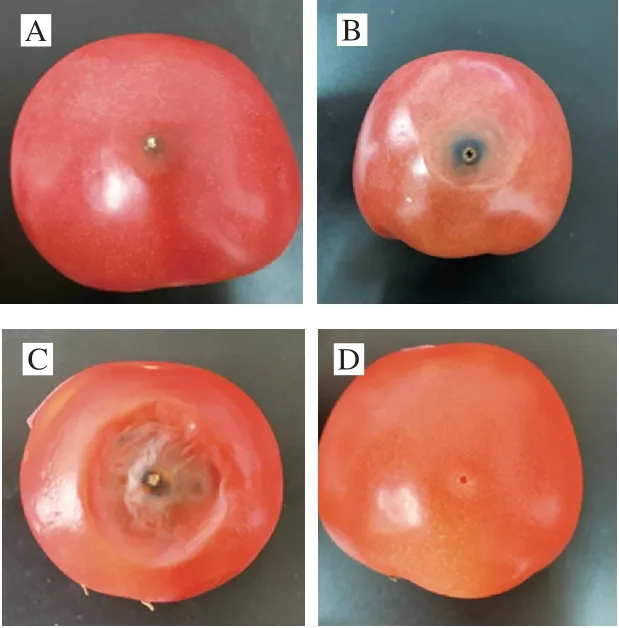
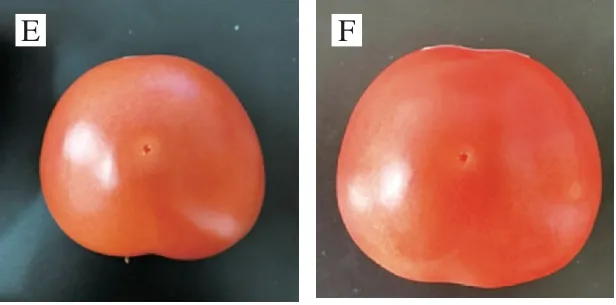
图1 接种后番茄果实在不同贮藏条件下的外观变化Fig.1 Change in visual appearance of tomato fruit under different storage conditions
2.2 不同贮藏条件下番茄样品中5 种真菌毒素的含量 变化检测结果
100 μg/L 5 种真菌毒素标准溶液的母离子色谱图如图2所示。不同贮藏条件下番茄样品中5 种真菌毒素的含量变化如表1所示。在4 ℃下贮藏15 d的过程中,接种处并无黑色病斑发生,也未检出5 种链格孢毒素。在25 ℃下,AOH、AME、TeA、TEN在3 次取样(5、10、15 d)中均有检出;ALT直至第15天才在取样中检出。

图2 5 种真菌毒素标准品母离子色谱图Fig.2 Mother ion chromatograms of five fungal toxin standards
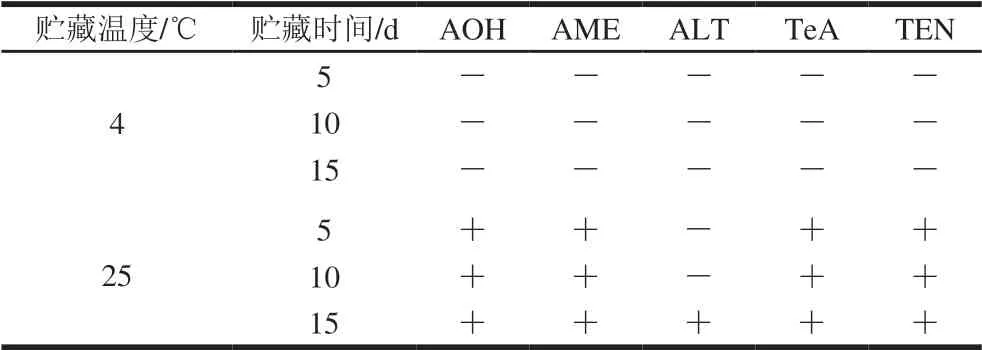
表1 不同贮藏温度下被浸染番茄的链格孢毒素检测结果Table 1 Results of detection of Alternaria toxins in infected tomatoes during storage at different temperatures
2.3 隐蔽型真菌毒素分析
对负离子模式二级质谱的原始数据的初步检测显示,几种代谢物解离的中性丢失值为m/z79.956 8,表明存在SO3。为了充分了解代谢物与硫酸盐的结合程度,使用MZmine软件对完整原始数据集进行碎片过滤诊断[26],设置诊断中性丢失值为m/z79.956 8,生成中性丢失图,中性丢失图可以使失去的相应SO3(m/z79.956 8)分子质量的前体离子进行可视化展示[27]。如图3所示,在接种链格孢霉后的番茄提取物中存在大量的硫酸化反应。
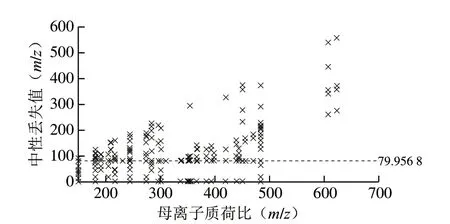
图3 接种链格孢霉番茄提取物的中性丢失图Fig.3 Mass spectrum showing neutral mass losses of metabolites extracted from tomatoes inoculated with Alternaria spp.
为了进一步验证硫酸盐结合的隐蔽型真菌毒素的存在,使用Xcalibur软件通过分辨率70 000以上的同位素模式进行确认。Heuillet等[28]的研究结果表明,高分辨质谱仪完全可以辨别出同位素信号,这对于化合物分子式的确认有极大的帮助。如图4、5所示,通过使用精确的链格孢霉毒素标准品m/z信息和m/z79.956 8的中性丢失值,可以得到AOH和AME的硫酸盐结合形式的色谱和质谱 信息。如图4所示,硫酸盐结合的毒素出峰时间早于未结合毒素的出峰时间,符合极性大的分子先出峰的原则。如图5所示,自然丰度为4.21%的34S同位素峰在单同位素峰分别有m/z1.995 54和m/z1.995 36的质量位移,13C×2同位素峰存在m/z2.004 94和m/z2.004 89的质量位移,由此证实了两种毒素硫酸盐结合产物的存在。
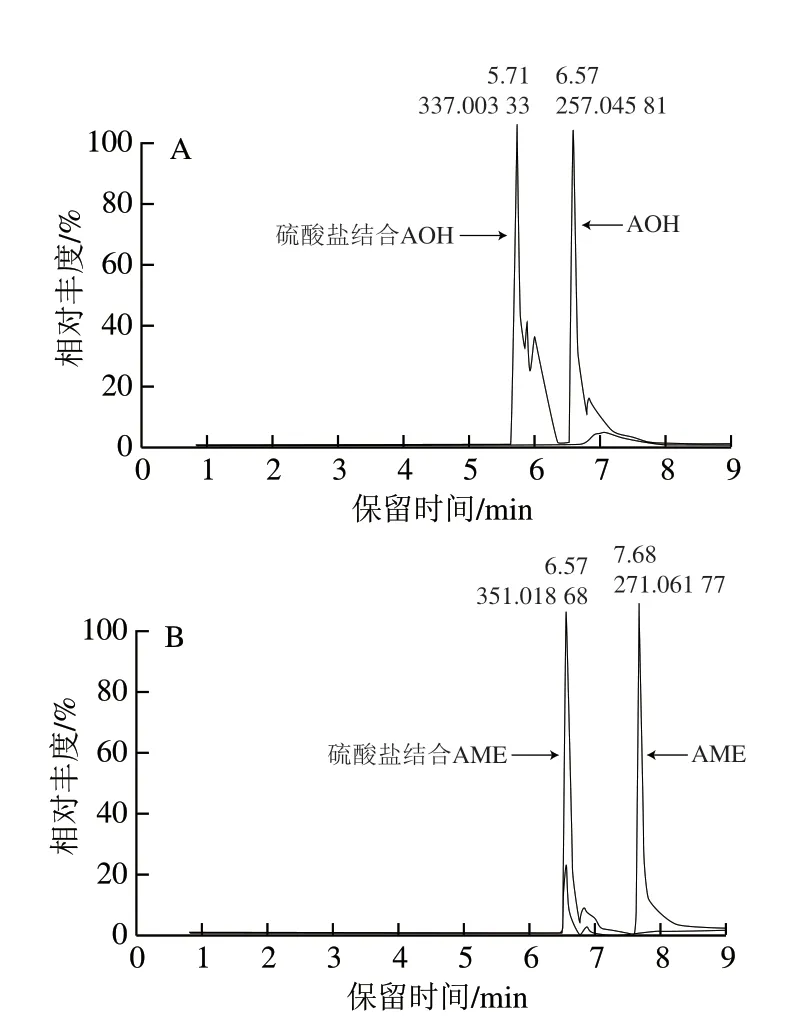
图4 硫酸盐结合和未结合的链格孢毒素色谱图Fig.4 Chromatograms of sulfate-bound and free Alternaria toxins

图5 SO3(m/z 79.956 8)的中性丢失和34S、13C同位素峰的质谱图Fig.5 Mass spectrum showing neutral mass loss of SO3 (m/z 79.956 8) and isotope peaks of 34S and 13C
2.4 隐蔽型真菌毒素的MZmine数据库及含量热图分析
对链格孢霉菌在番茄中产生的隐蔽型真菌毒素进行研究,结果发现番茄中链格孢毒素的隐蔽形式主要为硫酸盐结合产物。本实验中检测到AOH和AME的硫酸盐结合产物,其保留时间、实验m/z、理论m/z和质量测量准确度见表2。

表2 链格孢毒素及其与硫酸盐结合产物保留时间、 实验m/z、理论m/z和质量测量准确度Table 2 Retention time, measured m/z, calculated m/z and mass measurement accuracy of Alternaria toxins and their sulfated products
基于以上精准的分析数据,建立自定义数据库,数据库列表包括序号、m/z、保留时间、毒素名称和化学分子式等信息。利用Mzmine软件中自定义数据库筛查功能, 导入自定义数据库,设置m/z公差为0.02或5×10-6,对原数据集列表中所有的离子峰进行定性、定量和筛选。将筛选后的结果利用TBtools软件建立热图,热图可对各毒素的含量随时间的变化进行可视化分析。如图6所示,番茄接种链格孢霉后,5 d后便检测出AOH、硫酸盐结合AOH、AME、硫酸盐结合AME、TEN、TeA,直到15 d时才检测出ALT。贮藏期间各链格孢霉毒素整体含量呈增长的趋势,其中TeA的含量明显高于其他毒素。
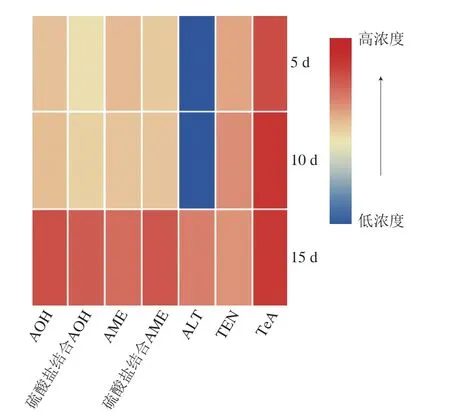
图6 链格孢霉毒素及其与硫酸盐结合产物的含量随时间变化的热图Fig.6 Heat map for changes in free and sulfate-bound Alternaria toxins over time
2.5 基于FBMN的代谢通路分析
使用MZmine 2软件对原始质谱数据进行质量检测和过滤,建立色谱峰。对色谱峰进行分离和去同位素峰处理,使用Join aligner功能将所有特征峰值列表对齐,最后导出包含完整一级质谱和二级质谱的特征文件上传至GNPS平台,使用FBMN工具创建一个分子网络[29]。设置母离子质量公差和子离子碎片质量公差均为m/z0.02。利用Cytoscape软件对分子网络结果进行可视化分析[30]。
图7为TeA(m/z196.097 92,化学式C10H15NO3,保留时间6.02 min)分子集群,该集群包括54 个节点,每个节点代表一种化合物。其中与TeA分子节点直接相连的有10 个节点。m/z182.081 3(保留时间为5.16 min)的节点是TeA脱去了分子某一侧的甲基后形成了分子式为C9H13NO3的化合物,其保留时间早于TeA,与其极性大、分子质量小有关。从GNPS平台上给出的FBMN结果表明,m/z644.228 5(保留时间为6.03 min)、612.276 3(保留时间为6.02 min)和415.192 3(保留时间为5.99 min)的节点为TeA的前体物质,它们具有和TeA相同的保留时间。其他6 个直接相连节点为TeA的一些其他代谢产物。

图7 基于FBMN的TeA代谢通路图Fig.7 Metabolic pathway map of TeA based on FBMN
由于AOH(m/z257.045 55,化学式C14H10O5,保留时间为6.58 min)和AME(m/z271.061 19,化学式C15H12O5,保留时间为7.71 min)有着相似的分子结构,所以处于同一分子网络集群。如图8所示,该网络包含23个节点。与AOH直接相连的有5 个节点。由FBMN工作流结果显示,m/z336.999(保留时间为5.77 min)、337.00 9(保留时间为7.80 min)和515.111 4(保留时间为6.48 min)为AOH的前体物质。其中m/z337.009的节点由于其保留时间晚于AOH,并不是前文所分析的硫酸盐结合的隐蔽型真菌毒素。m/z337.095 9(保留时间为5.89 min)和257.1757(保留时间为5.89 min)的节点为AOH的代谢产物。其中337.095 9节点为前文所分析的硫酸盐结合AOH(m/z337.0023 6,化学式C14H10O5SO3, 保留时间为5.74 min),由于处理软件的不同,其保留时间稍有偏差。毒素AME相连点只有3 个,其中m/z273.04(保留时间为6.77 min)的节点为AME的前体物质。m/z351.010 6(保留时间为6.55 min)的节点为前文所分析的硫酸盐结合AME(m/z351.018 01,化学式C15H12O5SO3,保留时间为6.58 min)。m/z287.057 4(保留时间为6.79 min)的节点为AME的另一代谢产物,它与AME的m/z相差16,在分子式上相差1 个氧原子。

图8 基于FBMN的AOH和AME代谢通路图Fig.8 Metabolic pathway maps of AOH and AME based on FBMN
如图9所示,ALT(m/z291.087 41,化学式C15H16O6,保留时间为5.89 min)与TEN (m/z413.219 43,化学式C22H30N4O4,保留时间为6.57 min)为两个孤立的节点。未在番茄果实内检测到其代谢产物及其隐蔽型真菌毒素,这与Li Yanshen等[15]在人工培养基中培养的链格孢霉实验结果相似。

图9 基于FBMN的TEN和ALT代谢通路图Fig.9 Metabolic pathway maps of TEN and ALT based on FBMN
3 结 论
本实验利用高分辨质谱结合GNPS、Xcalibur、MZmine软件或平台对链格孢霉感染番茄后产毒机制进行了分析。番茄果实接种链格孢霉后在不同温度下贮藏15 d,每5 d取样1 次进行分析。在4 ℃下贮藏过程的番茄果实中未检测到任何毒素;在25 ℃下贮藏的番茄果实中,除ALT外,AOH、AME、TeA、TEN在5、10、15 d时均被检出,ALT在贮藏15 d后才被检出。链格孢霉感染后的番茄在贮藏过程中,TeA含量最高。相较于室温(25 ℃),番茄更适合在低温(4 ℃)下贮藏。使用Mzmine软件的诊断碎片过滤工具,根据质谱中m/z79.956 8的特异性中性丢失值筛选出了硫酸盐结合的隐蔽型真菌毒素,并使用Xcalibur软件通过与5 种毒素标准品比对进一步验证了AOH和AME的硫酸盐结合隐蔽型真菌毒素的存在。将原始质谱数据集经过Mzmine软件处理,导入GNPS后软件利用FBMN软件建立了毒素的可视化代谢通路图。FBMN结果显示,3 种毒素(AOH、AME、TeA)有明显的代谢通路,代谢产物的种类较多,而TEN和ALT在代谢通路图中节点孤立存在,未在番茄中检测到代谢产物。结果表明MZmine、Xcalibur结合GNPS的数据分析方法适用于真菌天然产物的分析,实验对于番茄的链格孢霉污染防控和链格孢霉毒素代谢研究提供了理论依据和数据支撑。
猜你喜欢
现代仪器与医疗(2022年1期)2022-04-19
哈尔滨工业大学学报(2022年5期)2022-04-19
烟草科技(2022年2期)2022-03-04
建材发展导向(2021年15期)2021-11-05
甘肃教育(2021年10期)2021-11-02
绿色科技(2021年16期)2021-09-09
食品安全导刊(2021年20期)2021-08-30
现代仪器与医疗(2021年2期)2021-07-21
塔里木大学学报(2020年3期)2020-10-29
浙江中医药大学学报(2020年2期)2020-04-01
