
黑缘烟蟋螽丝腺结构
2021-08-18 10:59窦玉洁赵会敏石福明常岩林
昆虫学报 2021年7期
窦玉洁, 赵会敏, 石福明, 常岩林
(河北大学生命科学学院, 河北保定 071002)
昆虫唾液腺是开口于口前腔的多细胞腺体,依据其开口的位置,分为上颚腺、下颚腺和下唇腺(黄志君等, 2007),以下唇腺最为普遍。对昆虫唾液腺的研究,模式生物果蝇研究得相对较清楚(Berendes, 1965; Harrod and Kastritsis, 1972; Thomopoulos and Kastritsis, 1979; Thomopoulosetal., 1992)。直翅目中,沙漠蝗Schistocercagregaria的唾液腺为泡状腺体,腺泡位于胸部两侧,导管末端开口于舌基部的唾窦。每个腺泡由壁细胞、酶原细胞、导管细胞、气管母细胞、鞘细胞和色素细胞组成(Kendall, 1969)。飞蝗Locustamigratoria唾腺细胞超微结构显示,细胞核很大,在腺泡中的分泌物排出后,腺泡空泡化,呈现出较多的粗面内质网和线粒体(甘雅玲和郭中伟, 2002)。对半翅目叶蝉Hishimonuslamellatus、长翅目的蝎蛉科、膜翅目小家蚁Monomoriumpharaonis、缺翅目昆虫唾液腺的形态和结构也有一些报道(Maetal., 2011; Boonen and Billen, 2016; Dallaietal., 2017; Daietal., 2019)。
吐丝昆虫的丝腺是由外胚层发育来的,有真皮腺来源的丝腺、马氏管或附性腺来源的丝腺和下唇腺特化的丝腺(Sehnal and Akai,1990; 黄志君等, 2007)。下唇腺特化的丝腺主要集中在鳞翅目(Akai, 1971, 1984, 1986; Akaietal., 1993; Victoriano and Gregorio, 2002; Sorensenetal., 2006)和纺足目(Büsseetal., 2015, 2016)。纺足目和直翅目的蟋螽是仅有的幼期和成虫期均具吐丝行为的类群(Scholtzetal., 2018)。蟋螽若虫与成虫均可吐丝,用丝把植物的叶片缀起来(对折)做成巢,其白天隐藏巢内,夜晚外出捕食、求偶和交配等,蟋螽若虫在孵化后数天内开始筑巢,并且能够准确识别自己的巢,蟋螽不同个体间巢的结构和丝的密度基本一致(Rentz and John, 1990; Lockwood and Rentz, 1996; Hale, 2000)。
目前,对蟋螽吐丝的研究仅局限于吐丝行为的观察和丝腺外部形态的描述以及丝蛋白成分的分析(Walkeretal., 2012),蟋螽的丝腺来源于唾液腺(下唇腺)(Sutherlandetal., 2010),丝腺至少分泌2种蛋白——丝胶蛋白和丝心蛋白(Sehnal and Akai, 1990; Hale, 2000)。对于蟋螽丝腺的显微和超微结构未见报道。本研究对蟋螽科黑缘烟蟋螽Capnogryllacrisnigromarginata(Lietal., 2014)成虫丝腺的整体形态、显微结构和超微结构进行了观察,研究发现蟋螽丝腺分泌细胞分为Ⅰ型分泌细胞和Ⅱ型分泌细胞,并根据围细胞的位置和形态结构,推测分泌物从分泌细胞向外运输过程中,围细胞微绒毛腔的微丝束对分泌物的外排提供推动力。
1 材料与方法
1.1 试虫和腺体样品
黑缘烟蟋螽雄性成虫,于2019年6月20日采自广东省乳源县南岭(24.93°N, 113.02°E),成虫活体带回实验室,单头单笼短暂饲养。
解剖野外采集的黑缘烟蟋螽雄性成虫,手捏前胸背板,剪去足,并从背部剪开,暴露出整个消化道,摘除消化道;沿舌基部的腺管向后,从胸部腹侧内壁轻轻刮取,得到完整的腺体样品。
1.2 解剖学观察
将解剖分离的腺体用甲醇∶冰醋酸(3∶1)溶液固定15~20 min后,相机拍照。所用单反相机型号为Canon E0S 7D,微距镜头型号为Canon MACRO 100 mm。
1.3 苏木精-伊红(H.E.染色)染色观察
将解剖分离的腺体置于Bouin’s固定液中固定24 h。70%, 85%, 95%和100%梯度酒精脱水,环保型脱蜡透明液透明,石蜡渗透包埋,石蜡切片机切片,粘附于载玻片上,切片厚度为6 μm。室温下,根据标准流程进行常规的苏木精-伊红(hematoxylin-eosin, H.E.)染色,Olympus BX51光学显微镜观察拍照。
1.4 PAS-苏木精染色观察
将解剖分离的腺体置于新鲜配制的4%多聚甲醛溶液中,在4℃下固定48 h。然后在0.5 mol/L蔗糖中脱水,于-20℃下用O.C.T.包埋。用Leica CM1860冷冻切片机将包埋块切成5 μm厚的冷冻切片,并粘附于载玻片上。室温下,用高碘酸Schiff液(periodic acid Schiff, PAS)以及苏木精染色,Olympus BX51光学显微镜观察拍照。
1.5 激光共聚焦显微镜观察
将解剖分离的腺体置于新鲜配制的4%多聚甲醛溶液中,在4℃下固定12~24 h。然后在0.5 mol/L蔗糖中脱水,于-20℃下用O.C.T.包埋。用Leica CM1860冷冻切片机将包埋块切成5 μm厚的冰冻切片,并粘附于载玻片上。在室温下,用1% Triton X-100将组织处理30 min。然后用5% BAS封闭处理1 h;接着用异硫氰酸标记的抗α-微管蛋白鼠单克隆抗体(anti-α-tubulin-FITC, mouse monoclonal, Sigma-Aldrich)1∶100稀释,室温下孵育1 h,PBS漂洗。再将组织用鬼笔环肽-罗丹明(phalloidin-tetram ethylrhodamine B isothiocyanate, Sigma-Aldrich)染色45 min,PBS漂洗。接着,用DAPI染色15 min,PBS漂洗。之后在FV3000激光共聚焦显微镜下观察,拍照。
1.6 扫描电镜观察
将解剖分离的腺体迅速切割成小块,置于2.5%戊二醛溶液中,4℃固定1 h,PB漂洗3次;转至四氧化锇溶液中,4℃固定2 h,PB漂洗3次;梯度酒精脱水后,丙酮置换。将样品置于Leica EM CPD300全自动临界点干燥仪干燥,用导电胶将样品置于日立TM300台式扫描电子显微镜样品台上观察(加速电压5 kV),拍照。
1.7 透射电镜观察
将解剖分离的样品迅速切割成1 mm3的小块,置于2.5%戊二醛溶液中,4℃固定1 h,PB漂洗3次;转至四氧化锇溶液中,4℃固定2 h,PB漂洗3次;梯度酒精脱水后,相继在丙酮/环氧树脂Epon812(1∶1, v/v)和纯环氧树脂Epon812中渗透,再转至环氧树脂Epon812中于60℃下聚合包埋48 h。Reichert-Jung超薄切片机切片,1 μm半薄切片用甲苯胺蓝染色定位,Olympus BX51光学显微镜观察拍照。50 nm超薄切片用柠檬酸铅和醋酸铀双染,JEM-100SX透射电子显微镜下观察(加速电压80 kV),拍照。照片通过Adobe Photoshop CS(ps)软件进行处理排版,使用画图软件为CorelDRAW12。
2 结果
2.1 丝腺形态
黑缘烟蟋螽雄性成虫(图1: A)丝腺由丝腺导管和葡萄样腺泡组成,腺泡附着于胸部腹板两侧,对称分布,腺管的总导管从舌基部下方与下唇之间的唾窦发出,在离开唾窦不远处分为左右2支,向后延伸至胸部,逐级分支变细,并与分布于胸部腹侧的成簇的腺泡相连,腺体呈乳白色(图1: B)。每个腺泡可能含有数个细胞群(图1: C, D)。在扫描电镜下观察到腺泡表面凹凸不平,突起常为三角形,分支导管末端深入腺泡内部,腺泡周围有许多微气管伸入组织内部(图1: D),导管横截面有纵向的纹路,外表面光滑(图1: E)。

图1 黑缘烟蟋螽雄性成虫(A)与丝腺的解剖结构(B~E)
2.2 腺泡细胞结构
黑缘烟蟋螽丝腺腺泡外围由鞘细胞伸长形成的结缔组织鞘包围(图2: A, B),鞘细胞的细胞核扁平且长,具有明显的细胞核边界。细胞质在细胞核周围为一薄层,细胞质中细胞器较少,伸长的结缔组织纤维是分层的(图2: C)。

图2 黑缘烟蟋螽丝腺腺泡的鞘细胞
丝腺腺泡内包含4种类型的细胞,Ⅰ型分泌细胞、Ⅱ型分泌细胞、围细胞和腔细胞(图3~6)。
分泌细胞又可以分为2类:Ⅰ型分泌细胞(图3)和Ⅱ型分泌细胞(图4)。Ⅰ型和Ⅱ型分泌细胞PAS染色呈不同颜色,Ⅰ型分泌细胞靠近腺泡中心,细胞质为红色;Ⅱ型分泌细胞位于腺泡外围,未被染成红色。
腺泡内有2种运输细胞:围细胞(图5)和腔细胞(图6)。运输细胞较分泌细胞小,细胞核明显位于细胞一侧。
2.2.1Ⅰ型分泌细胞:细胞靠近腺泡中心,细胞很大,形状不规则,PAS染色为红色,PAS染液与细胞内的多糖结合,因此推测S1分泌颗粒为糖蛋白颗粒(图3: A)。甲苯胺蓝染色显示,细胞质中有大量的着色颗粒(图3: B)。
细胞核大,圆形,直径为11~13 μm,核孔明显(图3: C)。胞质内含大量的粗面内质网,内质网上附着有核糖体,胞质中游离核糖体较少,它们大多沿粗面内质网的膜排列。在电子显微照片中,这些膜系统通常以松散的平行堆叠的形式出现,内质网排列规则(图3: D, E),细胞质内有大量的球形分泌颗粒,大小不一,分泌颗粒为均匀的电子致密颗粒(图3: C, D)。
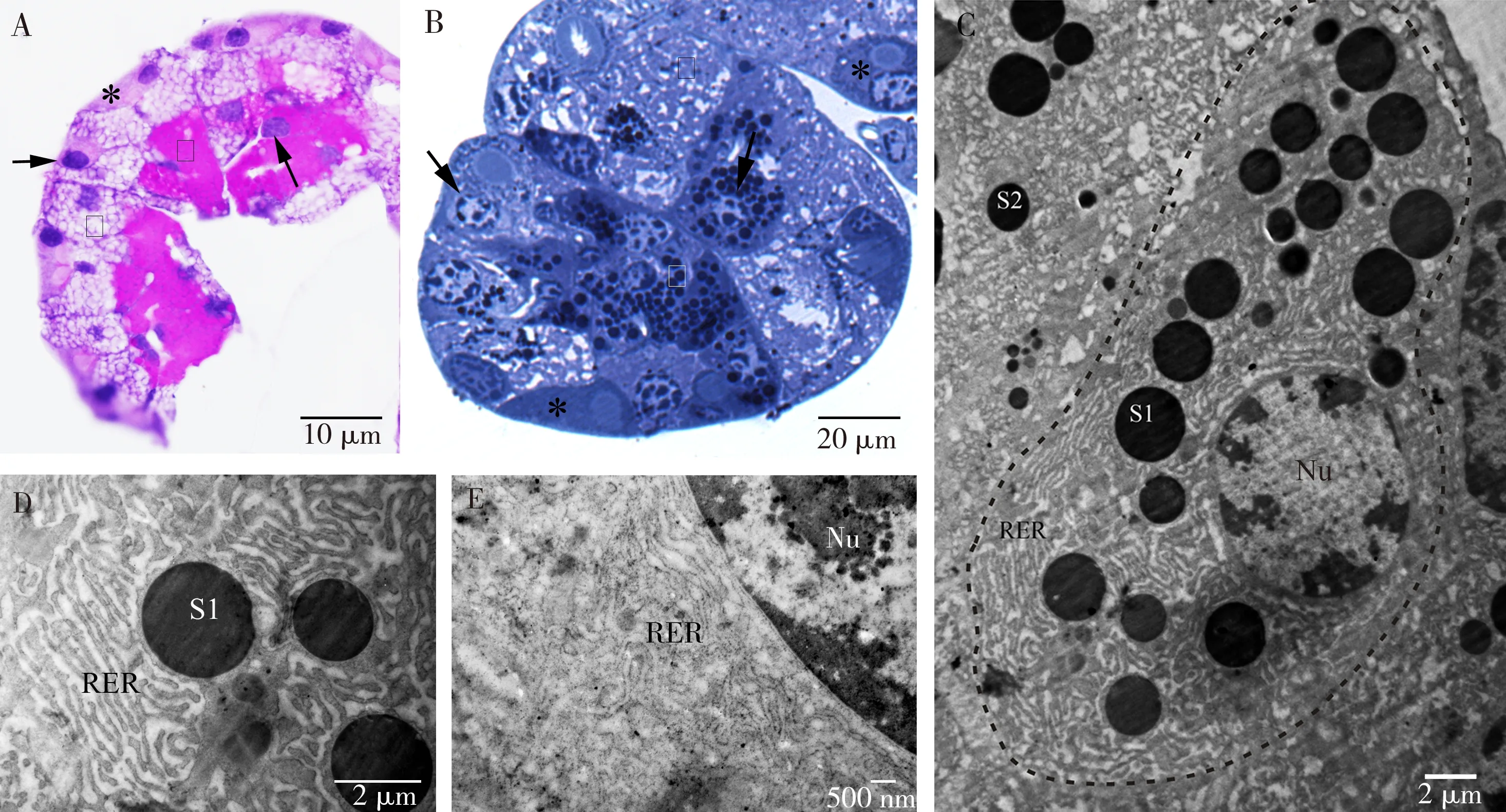
图3 黑缘烟蟋螽丝腺腺泡的Ⅰ型分泌细胞
2.2.2Ⅱ型分泌细胞:一般位于Ⅰ型分泌细胞和围细胞(或结缔组织鞘)之间(图4: A),细胞很大,形状不规则,PAS不能将其染为红色(图4: A),甲苯胺蓝染色显示,细胞质着色浅,甲苯胺蓝着色颗粒无或极少(图3: B)。
细胞核圆形,直径12~16 μm,核孔明显,核仁位于细胞核近中央(图4: B)。细胞质结构松散,胞质内散布短的排列不规则的滑面内质网,其间分布有大量圆形或椭圆形分泌颗粒。颗粒大小不一,内部可见螺旋片层结构,电镜观察结果电子致密程度不同,片层结构类指纹状(图4: C, D)。

图4 黑缘烟蟋螽丝腺腺泡的Ⅱ型分泌细胞
2.2.3围细胞:常成对排列,位于腺泡外层,与结缔组织鞘接触,三角形,宽基部靠近结缔组织鞘,较尖端指向腺泡中心,插入分泌细胞之间。细胞核位于细胞宽基部,圆形,直径10~12 μm,细胞质内具1微绒毛腔,微绒毛腔为锥形,微绒毛腔与细胞核大小相似,由细胞膜内陷形成(图5: A~C)。微绒毛腔内充满排列整齐的微绒毛(图5: D, E, H),细胞质内可见大量的线粒体(图5: G),常聚集于微绒毛腔周围(图5: H)。
鬼笔环肽-罗丹明荧光染色表明有成对存在的红色环形,即微绒毛腔,微绒毛腔的细胞质一侧以及细胞质基质被染成绿色(图5: I~L)。由于鬼笔环肽特异性地与细胞中的微丝结合,因此推测微绒毛腔由密集微丝束支撑。异硫氰酸标记的抗α-微管蛋白抗体证明,围细胞的细胞质中存在大量的微管蛋白。
2.2.4腔细胞:分布于分泌细胞之间(图6),细胞较小,形状不规则,细胞核形状不规则,占据细胞的大部分空间。腔细胞包围1圆形的胞外空腔通道,细胞及细胞核形态与胞外通道相适应,腔细胞连接处为加厚的膜结构(图6: F),在未加厚的部位,分泌细胞向通道内延伸形成短的微绒毛结构,不同分泌细胞的分泌颗粒聚集在这些微绒毛周围,向通道内分泌物质。
免疫荧光染色显示,有微绒毛的部位呈现红色,细胞外通道被异硫氰酸标记的抗α-微管蛋白抗体染成绿色,胞外通道内的微绒毛与围细胞微绒毛腔的微绒毛均由微丝蛋白支撑(图6: G~J)。

图6 黑缘烟蟋螽丝腺腺泡的腔细胞
2.2.5围细胞与腔细胞的连接:围细胞的微绒毛腔向腺泡中心延伸,与腔细胞包围的胞外通道相连(图5: D),在微绒毛和通道连接的空腔内看到小的电子致密颗粒和角质层的碎片,表明微绒毛腔为胞外空腔通道的末端(图5: F)。
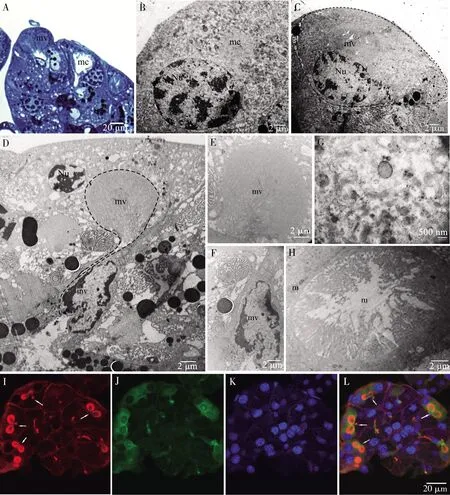
图5 黑缘烟蟋螽丝腺腺泡的围细胞
2.3 丝腺导管
丝腺导管连接唾窦和胸腔内腺泡,在蟋螽吐丝过程中起到运输丝蛋白的作用。显微观察显示丝腺的总导管和分支导管细胞在结构上无差别。
显微结构显示,导管横切面为环形,由单层细胞构成,细胞核较大,位于细胞中央,管腔内侧具明显的突起,细胞质内具清晰的纵纹(图7: A)。
透射电镜下观察导管细胞结构与腺细胞结构显著不同。细胞质内未观察到分泌物质,细胞外围可观察到显著的规则排列的深的质膜内陷,延伸至细胞的约1/3处(图7: B),形成细胞质隔室,细胞质隔室内可观察到大量的长的线粒体(图7: C, D)。细胞核形状不规则,位于细胞中央。细胞靠近管腔一侧具一层连续的小的突起,排列整齐,在导管壁的表皮下紧密聚集(图7: E)。
荧光染色观察显示,导管细胞外围红色染色较深(微丝),细胞靠近管腔内侧绿色染色较深(微管蛋白)(图7: F~I)。
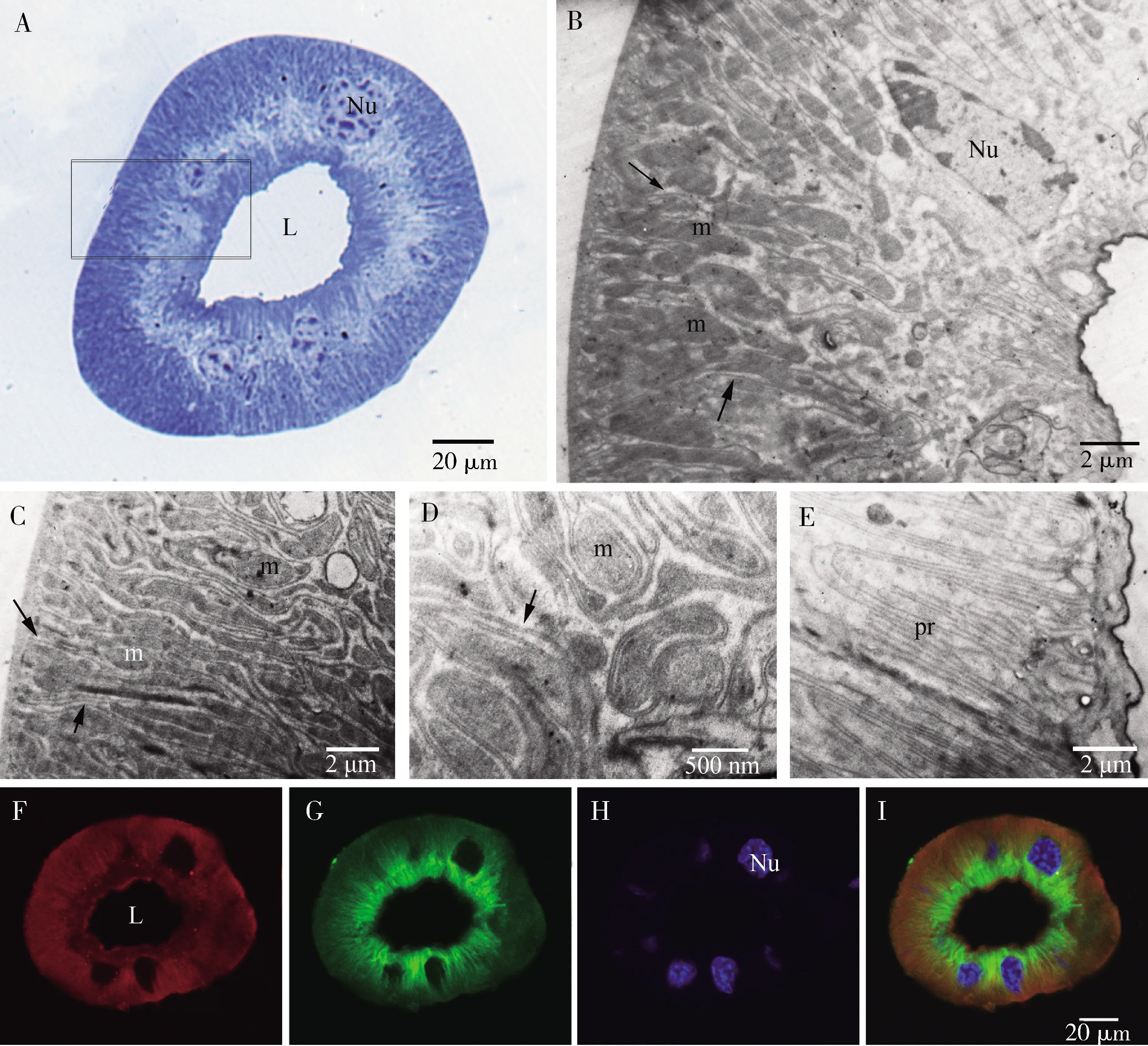
图7 黑缘烟蟋螽丝腺导管
2.4 丝腺腺泡细胞结构模式
依据黑缘烟蟋螽丝腺腺泡显微结构观察结果(图2~4),结合其超微结构(图2~6),绘制了丝腺腺泡结构示意图(图8):黑缘烟蟋螽丝腺泡外被鞘细胞形成的结缔组织,腺泡的主体包含4种细胞,即Ⅰ型分泌细胞、Ⅱ型分泌细胞、腔细胞和围细胞。分泌细胞大,细胞质内含有大量的内质网和分泌颗粒,Ⅰ型分泌细胞内含糖蛋白,超微结构显示为均匀致密的圆形颗粒;Ⅱ型分泌细胞的分泌颗粒为螺旋片层结构,染色深浅不一,但不含糖蛋白。腺泡内的胞外通道由单个腔细胞包围而成,腔细胞及细胞核形状不规则,分泌颗粒常常聚集在胞外通道周围。围细胞成对存在于腺泡的外围,呈三角形,细胞内具1由微丝束支撑的微绒毛腔。

图8 黑缘烟蟋螽丝腺腺泡细胞结构示意图
3 讨论
3.1 黑缘烟蟋螽吐丝行为
大多数鳞翅目仅幼虫具吐丝行为,蟋螽成虫和若虫均具有吐丝行为(Hale, 2000)。蟋螽吐丝时,将树叶卷曲,用丝从内侧缀起来形成巢穴,白天躲藏在内。在饲养蟋螽时,观察到黑缘烟蟋螽用丝将树叶卷曲成巢,当下唇接触到叶片表面时,在叶片表面黏合一层片状的丝,从片状丝抽离之后形成纤维状丝,再与另一侧的叶片表面通过片状丝相连,最终将树叶缝合成一个巢穴状的空间。
3.2 黑缘烟蟋螽丝腺来源于下唇腺,主要包含Ⅰ型和Ⅱ型分泌细胞
直翅目蝗虫唾液腺开口于舌下方,在唾液管离开不远处分为左右2支,皆通入胸部腹侧,每支腺管连接葡萄丛状的泡状腺。腺泡由2种细胞组成:酶原细胞和壁细胞(Kendall, 1969; 郭郛等, 1991)。蟋螽丝腺显微及超微结构观察显示出与直翅目唾液腺相似的结构,表明蟋螽丝腺与直翅目其他昆虫的唾液腺同源,蟋螽的丝腺来源于下唇腺(Hale, 2000),通过对丝腺形态和吐丝行为的观察表明,黑缘烟蟋螽丝腺由连接着唾窦的导管和葡萄样腺泡组成,腺泡用于分泌丝蛋白,腺管用于运输丝蛋白。与前人的研究结果相同,本研究认为黑缘烟蟋螽丝腺来源于下唇腺。
对黑缘烟蟋螽丝腺显微和超微结构的研究表明,丝腺腺泡外围由鞘细胞伸长形成的结缔组织鞘包围,包含Ⅰ型分泌细胞、Ⅱ型分泌细胞、围细胞和腔细胞4种细胞,分泌细胞起分泌作用,围细胞和腔细胞运输分泌物。PAS染色可以显示细胞内糖的分布(Suetal., 2017)。本研究通过PAS染色将黑缘烟蟋螽丝腺分泌细胞分为Ⅰ型和Ⅱ型,其中Ⅰ型分泌细胞靠近腺泡中心,发生PAS反应,证明Ⅰ型分泌细胞含有糖蛋白成分;Ⅱ型分泌细胞在腺泡外周,位于Ⅰ型分泌细胞与围细胞或结缔组织鞘之间,没有PAS反应(图3: A; 图4: A),说明Ⅱ型分泌细胞内不包含糖类物质。
电镜超微结构观察也显示,Ⅰ型分泌细胞和Ⅱ型分泌细胞分别分泌不同的颗粒,胞质内观察到大量的内质网和分泌颗粒。围细胞与鞘细胞接触,腔细胞分散在分泌细胞之间。分泌细胞的蛋白分泌物超微结构观察显示有2种:1种为均质的球形分泌泡(Ⅰ型分泌细胞分泌),PAS染色显示为糖蛋白;1种为具螺旋片层结构的分泌泡(由Ⅱ型分泌细胞分泌),形状为圆形或椭圆形(图3和4)。我们推测2种分泌物可能是丝的不同组成蛋白,对于2种物质的组成及其作用,还需进一步研究验证。蟋螽的丝腺没有用于储存丝蛋白的腔,推测丝腺可能是根据需求来进行分泌的。
3.3 微绒毛推动黑缘烟蟋螽丝腺分泌物从分泌细胞向丝腺导管的运输
在黑缘烟蟋螽丝腺导管和腺泡胞外通道中,未发现肌肉组织(图5~7)。围细胞细胞质中观察到大量线粒体,腔细胞细胞质中未观察到线粒体(图5),因此推测腔细胞无法向胞外提供物质运输所需的大量能量。电镜观察结果显示,围细胞的微绒毛腔向腺泡中心延伸,与腔细胞包围的胞外通道相连,在微绒毛和通道连接的空腔内看到小的电子致密颗粒和角质层的碎片(图5),表明腔细胞包围的胞外通道的末端为围细胞的微绒毛腔。荧光染色观察结果证明微绒毛腔的微绒毛主要由微丝束支撑(图5)。微丝参与许多细胞运动,如微绒毛的收缩、细胞的极化、胞内物质运输、细胞运动和胞吞胞吐作用等,微管也常常参与胞内物质运输、细胞器定位与迁移以及细胞运动等作用(Goodeetal., 2000; Lazaro-Dieguezetal., 2006; 邱鸿和于荣, 2009; Tang and Gerlach, 2017)。因此本研究推测,在分泌蛋白从分泌细胞向丝腺导管的运输过程中,动力可能主要是由微绒毛腔的微丝束收缩摆动提供的。
之前对蟋螽丝腺的研究较少,综合显微和超微结构(图8)以及形态、行为观察,推测黑缘烟蟋螽丝蛋白分泌过程大致为:分泌细胞内具有大量的内质网和核糖体,使分泌细胞能够合成大量的分泌颗粒;分泌细胞在与腔细胞包围的胞外通道的接触处,向通道内伸出短的微绒毛;由不同的分泌细胞合成的不同的分泌颗粒,分别聚集在分泌细胞与胞外通道连接处的微绒毛附近,分泌物质在此处从分泌颗粒的膜泡中排出,进入胞外通道;腔细胞包围的胞外通道与丝腺的分支导管相连,分泌物质经腺泡内胞外通道进入分支导管,然后到达总导管;通过唾窦,排出体外,液态蛋白遇空气最终形成固态的丝。即分泌物质在体内产生、分泌并释放到体外的过程依次经过分泌细胞、腔细胞包围的胞外通道、分支导管、总导管、唾窦。
猜你喜欢
中华胰腺病杂志(2022年1期)2022-11-22
西北民族大学学报(自然科学版)(2022年2期)2022-07-06
临床肝胆病杂志(2022年5期)2022-05-14
贵州医科大学学报(2022年2期)2022-03-31
课外生活·趣知识(2022年2期)2022-02-08
纺织科技进展(2021年4期)2021-07-22
兰州理工大学学报(2021年3期)2021-07-05
高考·中(2019年6期)2019-09-10
中华胰腺病杂志(2019年3期)2019-01-04
中国市场(2017年5期)2017-03-15
