
我国见血封喉适生区的气候特征分析
2021-08-06 08:54方良吴志华
热带作物学报 2021年2期
关键词:气候特征
方良 吴志华

摘 要:見血封喉具有重要的药用价值和使用价值,是我国热带季节性雨林的主要树种之一。了解见血封喉的适生区的气候因子情况,有利于其资源的保护和开发利用。本研究基于生态位模型理论对我国见血封喉地理分布区气候因子进行分析。以见血封喉在国内的现有天然地理分布数据为基础,通过DIVA-GIS获得见血封喉地理分布区气候因子;以分布样地内的海拔和气候数据20个指标进行主成分分析,结果获得4个主成分,其方差总贡献率为93.7%,其中第1主成分为海拔、降雨量和温度的综合因子;第2主成分为主要与低温有关的因子;第3主成分为主要与降雨量相关的因子;第4主成分为主要与极端温度相关的因子,其方差贡献率分别为42.9%、31.3%、13.0%和6.5%。以最大熵模型理论对见血封喉建立在我国适生区预测模型,预测的模型经ROC验证,模型的训练AUC为0.990,数据检验显示所建立的模型优良,预测精度高;刀切法检验结果表明,年平均气温bio1为影响见血封喉地理分布区气候影响分布的最关键生物气候因子,其次为平均日较差bio2。见血封喉在我国的适生区为广东、广西、海南、云南、台湾。最适生区面积最大的是广东,其次为广西,第三是海南;而高适生区和一般适合区主要是广西;云南和台湾为低适区。
关键词:见血封喉;气候特征;最大熵模型;适生分布
Abstract: Antiaris toxicaria Lesch. is one of the dominant tree species of tropical seasonal rain forest in China, it has important medicinal and use values. To understand the climate factors in the suitable area of A. toxicaria is conducive to its protection, development and utilization of resources. Based on the niche model theory, in this study the climatic characteristics and suitable distribution area of A. toxicaria were studied. Based on the existing natural geographical distribution data of A. toxicaria in China, the climatic data in the geographical distribution area of A. toxicaria were obtained from Worldclim data by DIVA-GIS. Principal component analysis (PCA) was carried out with 20 altitude and climate indicators. The results of the principal component analysis showed that there were four principal components obtained, the total contribution rate of variance was 93.7%. The first principal component was a comprehensive factor of altitude, rainfall and temperature, the second was related to low temperature,the third was mainly related to rainfall, the fourth was related primarily to extreme temperature, and its variance contribution rate was 42.9%, 31.3%, 13.0% and 6.5%, respectively. Based on the maximum entropy model theory, the prediction model of the A. toxicaria suitable area in China was established. The prediction model was validated by ROC, and the AUC of the training data was 0.990, and these results showed that the model was excellent with high prediction accuracy. The results of Jackknife test indicated that annual mean temperature (bio1) was the most key factor of biological climate which affecting the geographical distribution area of A. toxicaria, followed by mean diurnal range (bio2). The suitable areas for Antiaris toxicaria were Guangdong, Guangxi, Hainan, Yunnan and Taiwan in China. The excellent suitable area was Guangdong, followed by Guangxi and Hainan, while the high suitable and suitable area was Guangxi. Yunnan and Taiwan were the low suitable areas.
Keywords: Antiaris toxicaria; climate characteristics; maximum entropy model; suitable distribution
见血封喉(Antiaris toxicaria Lesch.),又名毒箭木,高25~40 m,胸径30~40 cm,为桑科(Moraceae)见血封喉属(Antiaris)常绿乔木,多生于海拔1000 m左右的热带雨林中,分布区域为我国的广东、海南、广西、云南以及斯里兰卡、印度、缅甸、泰国、马来西亚等地[1]。
见血封喉为剧毒植物和药用植物,其乳白色树液中含强心甙(antiarin)活性成分[2],已分离鉴定出37个化合物,其中26个强心苷及其苷元[3],民间入药,用于强心、催吐、泻下、麻醉等[4]。树叶中有(3S,5R,6S,7E,9R)-3,6-dihydroxy-5,6-dihy dro-β-ionol、(5R)-4,5-二氢布卢门醇A、槲皮素-3-O-β-D-葡萄糖苷、异鼠李素-3-O-β-D-芸香糖苷、山柰甲黄素-3-O-β-D-葡萄糖苷和环氧松柏醇等化学成分[5]。根中含有黄酮类化合物[2]。见血封喉树皮厚,纤维细长柔韧,为人造纤维原料,木材可作胶合板芯层、板凳以及包装箱用材等[6],有较高的经济价值。同时见血封喉树形高大挺拔,树形优美,枝叶繁茂,有层次,板状根奇特,耐贫瘠,抗风性强,为海南、广东省雷州半岛等地具有观赏价值的风水林树种。由于生境条件的变迁、人为因素及自然灾害等因子影响,其分布格局受到影响[7],大多呈单株分布,罕见小群体存在,其区域生态环境脆弱性显著化,生态景观呈现破碎化趋势[8],其自然种群的延续受到威胁,被列入2018年第一批广东省重点保护野生植物,中国珍稀濒危保护植物名录中的三级保护植物[7, 9-10]。目前,关于见血封喉生物学特性[6]、药用成分和药理性[2, 4-5]及栽培繁殖[10]方面的研究报道较多,但涉及其物种分布进行研究不多,仅限于马其侠等[11]以见血封喉的生态和地理分布等特征对华南的热带与亚热带界线划分研究。
植物的地理分布主要取决于气候条件,其中热量、水分以及耦合程度状况反映了气候的根本特征[12]。在空間地理大尺度上开展植物的地理分布的研究可以反映其固有特征及其对水热条件的需求。物种分布模型是将某物种在已知区域的发生率或丰度数据(分布数据)与这些位置的环境特征信息关联分析[13]。随着地理信息系统技术的迅速发展,目前物种分布预测模型主要有生物气候分析系统(BIOCLIM)、生态位因子分析模型(ENFA)、最大熵模型(MaxEnt)、基于规则集的遗传算法模型(GARP)等[14-15]。随着物种分布预测模型的应用,生物物种的自然分布可被精准地描述,并且这些信息已经被应用于研究各种科学和应用问题[13]。MaxEnt是基于最大熵方法来物种生态位建模和分布分析,是以生态位理论为基础的新兴研究领域[13]。当前在诸多预测模型中,MaxEnt是最有效的,特别是在物种分布数据较少的情况下,其仍然能得到较为满意的结果[16-17]。
在当前全球气候变化的背景下,了解见血封喉的地理分布格局对气候因子的响应情况,有利于见血封喉种质资源的保护和开发利用。本研究通过野外实地调查和文献查阅,充分收集了见血封喉资料,分析了其地理分布现状和特征,采用国际上常用的研究指标和方法结合相应气候资料,分析了见血封喉地理分布区的气候特征,并确定影响其地理分布的限制性气候因子,探究其适生区域,以期为见血封喉的保护及利用提供支撑。
1 材料与方法
1.1 见血封喉自然分布数据
根据近年来国家林业部门古树名木实地调查数据、标本数据库的搜索、相关论文文献记载,收集见血封喉的分布信息(表1)。通过查询广东古树名木信息管理系统,广西古树名木信息管理系统、全球生物多样性信息网(GBIF)、国家标本资源共享平台(NSII)和中国知网。结合现场踏查、文献资料记载,排除重复样本、无照片、错误鉴定、人工引种及文献来源模糊或地理信息模糊不清的标本分布点,最终获得见血封喉461个自然分布点,其地理位置信息落实到县级以下的行政单位。利用GoogleEarth和百度地图提取分布点的经纬度,至少精确到0.1。
1.2 环境数据获取
气候数据均来源于世界气候数据库(http:// www.worldclim.org/)。为Worldclim Version1.3,提供现在(1950—2000)气候数据,空间分辨率为2.5。利用DIVA-GIS软件,对分布点气候数据的19个生物气候因子进行提取(表2)。
1.3 气候特征及影响因子分析
在DIVA-GIS(Version 7.5)软件中提取见血封喉实际分布点的气候数据,获得19个气候指标(bio1~bio19)。以MaxEnt分析见血封喉分布点的气候特征,分别预测见血封喉的适生区,在上述基础上,以DIVA-GIS软件包进行地理适生分布区绘制。
1.4 分布模型精度检验
将见血封喉地理分布数据与提取的特定空间分布区域内的环境因子变量数据导入MaxEnt(Version 3.3.3)模型软件中。随机选取所有数据的75%作为训练集(training data),将剩余25%的数据作为测试集(testing data),用来验证模型[18]。构建见血封喉地理分布与气候关系MaxEnt模型,采用工作特征曲线ROC(receiver operating characteristic curve)分析法检验模型精度,ROC曲线基于非阈值依赖评判(threshold independent evaluation)模型精度。AUC值是ROC曲线与横坐标围成的面积值(the area under the ROC curve),取值范围0~1,评估标准分为极好(0.90~ 1.00)、好(0.80~0.90)、一般(0.70~0.80)、差(0.60~ 0.70)、失败(0.50~0.60),AUC值越接近1,区域环境变量与物种的区域分布的相关性越强,模型预测效果越好[16]。
在DIVA-GIS中加载MaxEnt的运算结果,进行适生等级划分和可视化。应用MaxEnt预测时,选择刀切法(Jackknife test),评估每个环境因子在10次重复预测中的重要性,并产生了环境因子对预测作用的响应曲线。设置了10次重复。并产生了10个预测随机模型,在此基础上,选择其平均进行分布预测分析。刀切法检验可获得反映不同环境变量对于分布增益的贡献大小的因子。依据模型“唯此变量(with only variable)”“除此变量(without variable)”以及“全变量(with all variable)”模拟时的训练得分进行判定标准为:“唯此变量”时得分较高,说明该因子重要,对物种分布贡献较大。
2 结果与分析
2.1 地理分布概况
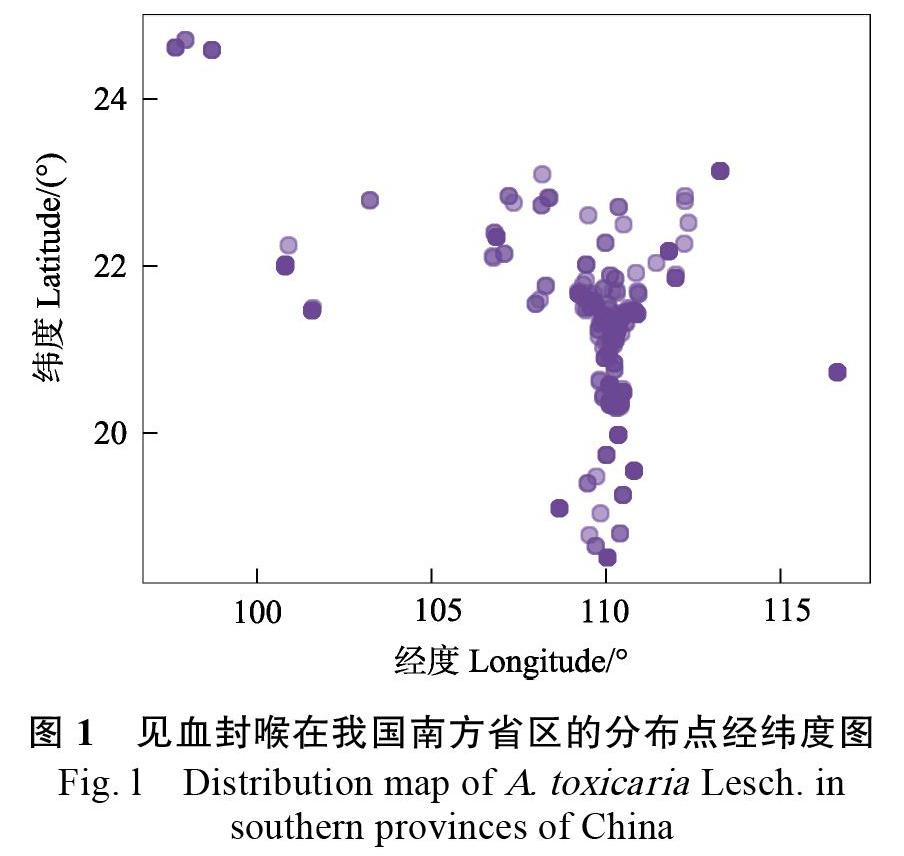
见血封喉主要分布在我国华南以及西南各省区,从图1可见,见血封喉自然居群主要见于我国广东、广西、海南、云南。从已有文献可查到其水平地理分布范围为北纬18°3036— 24°4236、东经97°3936—116°3748,最北可至云南龙陵一带,南至海南,东至东沙群岛,西至云南盈江一带(图1)。根据地形及气候特点,见血封喉的地理分布区主要为4个区域:(1)广东,主要分布于雷州半岛,包括雷州、徐闻、遂溪;其次是茂名、化州、信宜、阳江阳春等地。(2)海南,主要分布于海口、琼山、澄迈、儋州、昌江、文昌、万宁等地。(3)云南,主要分布于云南南部区西双版纳至西南部的勐腊等部分县市。(4)广西,主要分布于广西东部北流、博白,南部合浦、防城、龙州、南宁等地。
2.2 气候特征及主导因子分析
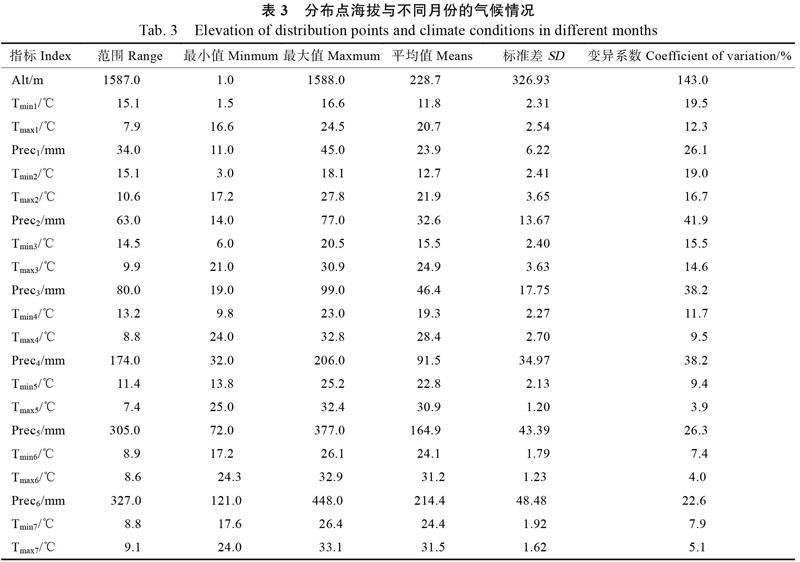
利用DIVA-GIS软件提取见血封喉不同的分布地点海拔(altitude,Alt)数据和1—12月的最低温(minimum temperature,Tmin)、最高温度(maximum temperature,Tmax)和降雨量(precipitation,Prec)气候数据,然后进行气候特征分析,结果见表3。见血封喉分布区海拔为1~1588 m,变异范围大,变异系数为143.0%,平均分布在228.7 m的低海拔区域;分布区最低气温在1月,月极端最低温平均为1.5 ℃,其次月最低温为2月和12月,月平均最低温均为3.0 ℃,月最低温度变异系数最大的为1月,为19.5%,最小为6月,变异系数为7.4%;最高温度在7月,月极端最高温平均为33.1 ℃,月最高温度变异系数最小的为10月,仅为3.0%,而最大的为2月,变异系数为16.7%;月均降水量为23.9~ 262.5 mm,最低为1月23.9 mm,其降雨量变化为11.0~45.0 mm,最高月为8月,平均为262.5 mm;最湿月(8月)降水量变化范围为145.0~528.0 mm,其月平均降水量变异最小,变异系数为20.0%,而月平均降水量变异最大的是10月,变异系数为44.4%。
作为一种常绿落叶乔木,见血封喉喜热不喜寒,从图2的分布点气候变量bio1、bio6频率分析可看出,见血封喉多分布在年平均气温(bio1)在20~25 ℃,集中分布在23 ℃的区域,最冷月最低温度(bio6)介于6~16 ℃,平均为12 ℃。
以分布樣地的海拔(Alt)和气候数据(bio1~bio19)20个指标进行主成分分析,结果降维获得4个主成分(表4),第1主成分PC1,方差贡献率为42.9%,第2主成分PC2,方差贡献率为31.3%,第3主成分PC3,方差贡献率为13.0%,4个主成分的累计方差贡献率为93.7%。按照表5系数绝对值大小,主成分PC1中,主要为海拔Alt(?0.92)、等温性bio3(?0.91)、平均日校差bio2(?0.90)、最干季度降水量bio17(0.89)、最干月降水量bio14(0.87)、最湿季度平均温度bio8(0.87)、最暖季度平均温度bio10(0.87),主成分PC1为海拔、降雨量和温度的综合因子;主成分PC2中主要为最冷季度平均温度bio11(?0.83)、最暖季度降水量bio18(0.83)、年平均气温bio1(?0.82)、最冷月最低温度bio6(?0.76),为低温相关的因子。主成分PC3中主要与降雨量相关的最湿月降水量bio13(0.63)、最湿季度降水量bio16(0.62)、最干季度平均温度bio9(0.60)、年降水量bio12(0.57)因子。主成分PC4中主要与极端温度相关的因子,主要为年均温变化范围bio7(0.65)、最暖月最高气温bio5(0.62)、平均日较差bio2(0.35)。
2.3 见血封喉的地理分布分析
从图3可见,Maxent模型的AUC值分别达到了0.990,标准偏差为0.005,该模型见血封喉的预测均达到优良等级,显著大于随机分布模型值(0.5),可以满足其适生区分布预测的需求。
依据见血封喉MaxEnt模型所得结果,按适宜指数进行适生区的划分为不适合区、低适区、一般适合区、高适合、最适合5类,并进行地理适生分布区分区(表6)。
预测见血封喉在我国的适生区如表7和表8所示。总体来看,见血封喉实际分布区与模拟的主要适生分布区较为一致,见血封喉在我国的适生区主要分布在我国南方海南、广东、台湾、广西、云南省(区),而最适生区、高适生区主要分布在海南、广东的雷州半岛以及广西南部和东部区域。最适生区面积最大的是广东省,为17 397.0 km2,其次为广西(10 309.6 km2),第三是海南,而高适生区和一般适合区均以广西最大,分别为18 083.9 km2和12 954.6 km2。云南除了南部西双版纳外,见血封喉最适生区和高适生区在云南其他区域仅为零星分布,不但分布个体数量少,而且可适生的区域不多。从分析结果可以看出台湾南部区域和岛屿有适生的区域,但总体来说,见血封喉的适生区域非常有限。根据包括低适应区在内的总适生区面积排名为:广西(59 270.3 km2)>海南(48 159.1 km2)>广东(44 946.6 km2)>云南(11 007.1 km2)>台湾(4270.3 km2)。
2.4 MaxEnt中各变量权重
季度降水量bio17(6.9%)5个因子。另外,最冷季度降水量bio19贡献率为3.5%,温度季节性变化标准差bio4贡献率3.4%。各个气候变量影响见血封喉生长适宜度中所占的权重结果显示(图4),唯此变量增益最高的环境变量是年平均气温bio1,其训练增益超过2.5,因此bio1是影响见血封喉分布的关键环境变量。平均日较差Bio2也是较为重要的环境变量;另外训练增益超过2.2的3个种主导变量的依据大小排序为:最干季度平均温度bio9>最干季度降水量bio17>最暖季度平均温度bio10。
3 讨论
环境因子对植物生长和分布以及多样性的形成具有重要影响[19]。对全球尺度而言,气候条件是影响限制植物物种分布和多样性的主要因素[20-21]。在植物资源保护和开发过程中遵循“适地适树”原则对树种研究,确定气候变化下适宜分布的空间,以确保达到预期目标。基于生态位理论的物种分布模型,就是依据物种的生态需求,以计算机模型研究气候变化对物种的影响以及预测物种的分布。目前已广泛应用于生态学与生物地理学[22]、外来入侵物种风险的评估[23]、濒危物种的保护[24]及气候变化对物种分布的影响等研究领域。
路丹桂等[25]在我国桑科植物属的空间分布及多样性研究中指出,见血封喉属从最西的云南盈江县到最东的广东陆丰县,最北至广西临桂县。
分布范围与本文研究预测结果基本一致,但需要指出广西临桂已经处于桂北冷凉的亚热带地区,在整个桂中区域不存在见血封喉的过渡区以及天然分布的其他文献资料,广西区内见血封喉仅分布于桂南的南宁、北海、合浦、陆川、博白、北流、崇左、龙州、凭祥区域[26],因此推测广西临桂采集的见血封喉标本可能来源于人工栽培植株。同样,尽管在我们研究的结果中显示粤东为见血封喉的潜在分布区域,但从《广东植物志》[27]以及近年来广东省林业古树名木普查中也没有见血封喉在该区域分布的文献。
作为一种常绿落叶乔木,见血封喉喜热不喜寒,见血封喉多分布在年平均温度(bio1)为20~25 ℃,集中分布在23 ℃的区域,最冷月最低气温(bio6)为6~16 ℃,平均为12 ℃。MaxEnt模型的刀切法检验进一步表明年平均气温bio1是影响见血封喉分布的关键环境变量,其次平均日较差bio2也是较为重要的环境变量。因此常夏无冬、热量资源丰富、寒潮发生较少、年均温高的地理区域有利于见血封喉分布生长。
本次研究将更为稳定和精准的MaxEnt模型应用于见血封喉潜在地理分布区预测,通过MaxEnt模型10次重复检测结果显示,训练集的 AUC值为0.990,模型的预测结果达到优秀水平,此次预测的地理分布结果与实际分布区域的相符度较高?见血封喉在我国天然分布于广东、广西、海南和云南四省(区),也是见血封喉最适生区、高适生区主要分布区域。见血封喉最适生区面积最大的是广东,其次为广西,第三是海南,而高适生区和一般适合区主要是广西。云南和台湾为低适区。总适生区面积分别为广西59 270.3 km2、海南48 159.1 km2、广东44 946.6 km2、云南11 007.1 km2、台湾4270.3 km2。将MaxEnt模型预测的见血封喉适生区分布与实际分布点进行比较,发现其分布点主要分布于高适生区内,研究结果的可靠性较高,可为见血封喉的保育提供科学支撑。
近几十年来,由于人们对热带森林的不合理开发,导致见血封喉资源日益匮缺,使其有濒临灭绝的危险。目前广东、广西古树名木调查发现见血封喉散生较多,种群多分布自然村落边,在受人类干扰较明显,种间关系极不稳定[7]。尽管见血封喉种子发芽率高,但寿命短,苗木易遭老鼠破坏,幼树成活率低[7],物种扩散能力有限,加之目前古树多以年代久远,空心、根腐现象严重,易风倒,这些均不利于其种群保护和复壮。有必要加强其原生境和现有植株的保护,并通过人工繁殖扩大栽培。因此,了解见血封喉与其生物学特性密切相关的气候特征及其适生区分布是非常必要的。见血封喉的分布跟许多外界因素有关。由于条件的限制,加之海拔因子为一个与气候因子密切相关的综合的生态因子[7],在模型初次预测中发现其影响不大,见血封喉在广东省主要分布区为土壤肥力低的砖红壤,广西分布区均为有机含量一般的赤红壤,因此本研究仅以19个生物气候因子以10次重复的随机模型进行预测,发现实际上有见血封喉生长的云南盈江分布地,预测不一定是其潜在的适生区,表明见血封喉在该分布地的生长发育更易受气候等其他因素变化的影响,今后该地区需更多注意见血封喉植物的保护和利用。同样,模型所得到一些潜在適生区也不一定都适合见血封喉的生长。本研究重点关注了气候因素对见血封喉的适生区的影响,而在以后的研究中,需要进一步关注如土壤、植被、小区域、人为干扰等其他环境变量的影响。
参考文献
中国科学院昆明植物研究所. 云南植物志: 第6卷[M]. 北京: 科学出版社, 1995: 595.
阙东枚, 梅文莉, 吴 娇, 等. 见血封喉根中类黄酮的结构鉴定[J]. 有机化学, 2009, 29(9): 1371-1375.
梅文莉, 干玉娟, 戴好富. 见血封喉化学成分与药理活性研究进展[J]. 中草药, 2008, 39(1): 151-154.
干玉娟, 梅文莉, 曾艳波, 等. 见血封喉乳汁脂溶性成分及其抗氧化活性研究[J]. 热带亚热带植物学报, 2008, 16(2): 144-147.
李 辉, 干玉娟, 梅文莉, 等. 见血封喉树叶化学成分研究[J]. 热带亚热带植物学报, 2009, 17(3): 302-304.
管志斌, 彭朝忠, 管松山. 见血封喉的生物学特性[J]. 南京林业大学学报(自然科学版), 2003, 27(5): 77-79.
罗 涛, 杨小波, 李东海, 等. 海口地区见血封喉种群数量特征及空间分布格局[J]. 福建林业科技, 2009, 36(3): 161-166, 174.
韩维栋, 陈 杰. 雷州半岛树木志[M]. 广州: 华南理工大学出版社, 2014.
国家环保局, 中国科学院植物研究所. 中国珍稀濒危保护植物名录[M]. 北京: 科学出版社, 1987.
易观路, 许方宏, 罗建华, 等. 我国濒危植物见血封喉及其栽培技术研究初报[J]. 热带农业科技, 2004, 27(2): 45-46.
马其侠, 王发国, 陈炳辉, 等. 从见血封喉的分布看广东省热带与亚热带的界线划分[J]. 热带亚热带植物学报, 2004, 12(1): 7-12.
阎传海. 植物地理学[M]. 北京: 科学出版社, 2001.
Gomez J J, Cassini M H. Environmental predictors of habitat suitability and biogeographical range of Franciscana dolphins (Pontoporia blainvillei)[J]. Global Ecology & Conservation, 2015, 3(7): 90-99.
乔慧捷, 胡军华, 黄继红. 生态位模型的理论基础、发展方向与挑战[J]. 中国科学: 生命科学, 2013, 43(11): 915-927.
周海涛, 那晓东, 臧淑英, 等. 最大熵(Maxent)模型在物种栖息地研究中的应用[J]. 环境科学与管理, 2016, 41(3): 149-151.
Phillips S J, Anderson R P, Schapire R E. Maximum entropy modeling of species geographic distributions[J]. Ecological Modelling, 2006, 190(3-4): 231-259.
Kruijer H J D, Raes N, Stech M. Modelling the distribution of the moss species Hypopterygium tamarisci (Hypopterygiaceae, Bryophyta) in Central and South America[J]. Nova Hedwigia, 2010, 91(3-4): 399-420.
Scheldeman X, Zonneveld M V.Training Manual on Spatial Analysis of Plant Diversity and Distribution[M]. Rome: Biodiversity International, 2010.
Pauses J G, Austin M P. Patterns of plant species richness in relation to different environments: an appraisal[J]. Journal of Vegetation Science, 2001, 12(2): 153-166.
劉冠成, 黄雅曦, 王庆贵, 等. 环境因子对植物物种多样性的影响研究进展[J]. 中国农学通报, 2018, 34(13): 83-89.
Anderson R P. A framework for using niche models to estimate impacts of climate change on species distributions[J]. Annals of the New York Academy of Sciences, 2013, 1297(1): 8-28.
李国庆, 刘长成, 刘玉国, 等. 物种分布模型理论研究进展[J]. 生态学报, 2013, 33(16): 4827-4835.
Santamarina S, Alfaro-Saiz E, Llamas F, et al. Different approaches to assess the local invasion risk on a threatened species: Opportunities of using high-resolution species distribution models by selecting the optimal model complexity[J]. Global Ecology and Conservation, 2019, 20: e00767.
武晓宇, 董世魁, 刘世梁, 等. 基于MaxEnt模型的三江源区草地濒危保护植物热点区识别[J]. 生物多样性, 2018, 26(2): 138-148.
路丹桂, 邱丽氚, 常 虹, 等. 中国桑科植物属的空间分布及多样性[J]. 西北植物学报, 2019, 39(6): 1127-1134.
李树刚. 广西植物志: 第3卷 种子植物[M]. 南宁: 广西科学技术出版社, 2011: 55.
中国科学院华南植物所. 广东植物志: 第1卷[M]. 广州: 广东科技出版社, 1987: 186.
责任编辑:黄东杰
猜你喜欢
农业与技术(2017年5期)2017-04-11
中国民族民间医药·下半月(2017年3期)2017-03-31
科技与创新(2016年24期)2017-03-30
现代农业科技(2016年22期)2017-03-24
科技与创新(2017年3期)2017-03-17
农业与技术(2017年2期)2017-03-16
农业与技术(2016年22期)2017-03-07
现代农业科技(2016年20期)2016-12-20
现代农业科技(2016年20期)2016-12-20
中国高新技术企业(2015年23期)2015-06-24
