
连作百香果对土壤理化性质和微生物特性的影响及病原真菌的分离与鉴定
2021-08-06 08:19陈晓婷王裕华林立文张清旭汪鹏丁力郭万财王海斌
热带作物学报 2021年2期
陈晓婷 王裕华 林立文 张清旭 汪鹏 丁力 郭万财 王海斌
摘 要:為了分析连作对百香果土壤理化性质及微生物的影响,本研究以种植0(未种植)、1、2、3 a的百香果根际土壤为材料,分析连作百香果对土壤理化指标、自毒潜力、微生物数量的影响,并对连作百香果根际土壤病原真菌进行分离、鉴定。结果表明,随着种植年限的增加,百香果根际土壤pH呈显著下降趋势,土壤总氮、磷、钾含量变化较小,而有效性氮、磷、钾含量及有机质含量呈上升趋势。自毒潜力分析结果表明,与对照(0 a,未种植)相比,种植1、2、3 a的百香果根际土壤对受体莴苣的抑制率分别为39.84%、54.36%、61.83%。微生物数量分析结果表明,随着百香果种植年限的增加,土壤细菌与放线菌数量呈显著下降趋势,而真菌数量呈显著上升趋势。相关性分析结果表明,百香果种植土壤的年限与土壤pH、细菌、放线菌数量呈极显著负相关,而与土壤有效氮、有效磷、有效钾含量及真菌数量呈显著或极显著正相关。微生物分离与鉴定结果表明,共分离到9种不同类型的真菌,其中真菌BXG 3、BXG 7、BXG 9可显著影响百香果的正常生长;鉴定结果表明,真菌BXG 3、BXG 7、BXG 9分别为层出镰刀菌(Fusarium proliferatum)、棕黑腐质霉菌(Humicola fuscoatra)和毛栓菌(Trametes hirsute)。
关键词:百香果;连作;土壤微生物;真菌
Abstract: In order to analyze the effect of continuous cropping of passion fruit on the soil physicochemical property and microorganism, the rhizospheric soils cultivated for 0, 1, 2 and 3 years were used as the materials. Soil physical and chemical indicators, autotoxicity potential, and microbial quantity were analyzed. Furthermore, the pathogenic fungi in rhizosphere soil were isolated and identified. The results showed that the pH value of the passion fruit rhizosphere soil showed a significant downward trend with the passage of planting. Total nitrogen, phosphorus and potassium content changed little, while available nitrogen, phosphorus, potassium and organic matter content increased. The results of autotoxicity potential analysis showed that the inhibition rates of 1-, 2-, 3-year-old rhizosphere soil on lettuce was 39.84%, 54.36% and 61.83%, respectively, compared with the control (0 years). The results of microbial quantitative analysis indicated that the number of soil bacteria and actinomycetes decreased significantly with the passage of planting, while the number of fungi increased significantly. Correlation analysis suggested that the planting years of the passion fruit soil was significantly negatively correlated with pH value, and the content of bacteria and actinomycetes, but significantly positively correlated with the content of available nitrogen, available phosphorus, available potassium and the number of fungi. The results of microbial isolation showed that nine different kinds of fungi were isolated, and the fungi of BXG 3, BXG 7 and BXG 9 could significantly affect the growth of passion fruit. The identification results showed that BXG 3, BXG 7 and BXG 9 fungi were Fusarium proliferatum, Humicola fuscoatra and Trametes hirsute, respectively.
Keywords: passion fruit (Passiflora edulis Sims); continuous cropping; soil microorganism; fungi
百香果,学名西番莲(Passiflora edulis Sims),又名鸡蛋果、热情果等,具有菠萝、香蕉、苹果、柠檬等多种水果的复合芳香,是西番莲属草质藤本植物[1]。目前,我国种植的百香果主要有黄色百香果(P. flavicarpa)和紫色百香果(P. edulis)2种。百香果是多年生的藤本植物,第一次种植时年产量较高,然而从第二年开始,产量逐渐下降,病虫害暴发频率增多,这种现象产生后,主要以施肥和使用农药为主进行改善,但效果并不明显[2-3]。
土壤是植物种植的载体,微生物作为土壤微生态系统的重要组成部分,发挥着极其重要的作用,例如,在土壤物质转化、分解,土壤肥力改变,土壤质地的改善等[4-5]。细菌、真菌、放线菌等微生物类群是土壤微生物群落结构的主要组成部分,其在土壤中的数量、种类及其结构的变化,直接或间接地影响着植物根际土壤的微生态功能[6-7]。土壤微生态系统中的微生物总量与活性和有益微生物数量的高低是判断土壤活跃性的重要指标,而作物连作极易导致植物释放物质的积累加剧,物质长期大量积累抑制了土壤有益微生物生长,滋生病原菌,使连作土壤中的微生物结构发生变化,总量减少[8-10]。此外,人为因素的干扰也加剧了土壤微生态环境的变化。例如,百香果的单一种植导致病虫害的爆发并蔓延,因此种植户为了有效控制百香果病虫害,导致农药的大量使用[11]。农药的大量使用打破了原有的土壤微生态系统,致使土壤中益生菌数量减少,病原菌数量增多且产生了一定的抗药性。例如,连作玉米导致根际土壤中的荧光假单胞菌数量增加,释放的代谢产物氢氰酸含量加大,对玉米自身生长的毒害作用加剧[12]。可见,作物长期连作后,土壤微生物种群发生变化,土壤微生态系统的平衡失调,进而导致作物生长受阻。Wang等[13]前期研究发现,连作导致百香果的生理保护能力降低,光合作用能力降低,最终导致百香果的产量和品质下降。然而,这种现象的产生是否与土壤中的微生物,特别是病原微生物存在一定的联系,尚未见相关文献报道。
据此,本研究以种植1、2、3 a的百香果根际土壤及未种植(0 a)百香果的土壤为材料,分析连作百香果对土壤理化指標、自毒潜力,微生物数量的影响,并分离、鉴定连作百香果根际土壤病原真菌,以期为连作百香果土壤的修复和连作障碍的消减提供一定的研究基础。
1 材料与方法
1.1 材料
以种植0、1、2、3 a的百香果根际土壤为材料,其中0 a土壤为同一区域未种植百香果的土壤。取样点位于福建省龙岩市新罗区,海拔600~ 700 m,北纬250°,东经116°,年降雨量1700 mm,相对湿度80%,年平均气温16~20 ℃。
收集种植1、2、3 a的百香果根际土壤,取样方法:每个年份的百香果种植地随机选择12株百香果,移除土壤表面的树叶,挖去周围土壤,收集百香果根际土壤。每个年份百香果种植地,4株百香果为1个样本,以同一区域未种植百香果的土壤为对照(0 a),收集3个重复样本,每个样本300 g。收集的土壤材料分为2部分,一部分鲜土用于土壤自毒潜力测定、土壤微生物数量测定及土壤真菌的分离;另一部分土壤自然风干后,粉碎并过60目筛,采用四分法取样,用于土壤理化指标测定,土壤理化指标测定参照文献[14]的方法。
1.2 方法
1.2.1 土壤自毒潜力测定 采用土壤琼脂三明治法对不同种植年限的土壤样本进行自毒潜力测试[10]。具体步骤:将冷却至45 ℃的30 mL琼脂(0.8%)加入培养皿(直径5 cm)中,取土壤样品各15 g,混合固化后,在土壤-琼脂层表面添加琼脂(0.5%)2 mL,以减少水分流失。冷却后,将10颗预萌发的莴苣种子置于琼脂培养基中,放置于培养箱中培养,培养条件为温度25 ℃、光照12 h(7:00—19:00),培养3 d后测定莴苣的根长,设置6个重复。根长相对抑制率(IR)的计算公式:IR=(1-处理值/对照值)×100%。
1.2.2 土壤微生物数量测定 采用稀释涂抹平板计数法[15]测定土壤中细菌、真菌、放线菌的数量。细菌采用葡萄糖牛肉膏蛋白胨培养基培养,真菌采用马丁孟加拉红培养基培养,放线菌采用淀粉硝酸钾培养基培养,微生物数量以每克样品的菌数表示(CFU/g)。
1.2.3 土壤微生物的qRT-PCR定量分析 土壤微生物总DNA的提取采用Bio 101 FastDNA SPIN Kit(Bio 101, Inc., USA),土壤用量为0.5 g鲜土[16]。DNA检测采用1%的琼脂糖凝胶电泳,DNA纯化采用胶回收试剂盒(TianGen Biotech Co., Ltd),纯化后的DNA置于-20 ℃中保存,备用。
细菌定量分析所用的引物[17]为F27:5- AGAGTTTGATCMTGCCTCAG-3,R1492:5- TACHHYTACCTTGTTACGACTT-3,PCR反应体系25 μL,其中模板DNA、SYBR Premix Ex Taq(TaKaRa Biotechnology)、F27引物、R1492引物、ddH2O等,分别为0.5、12.5、1、1、10 μL;PCR反应程序:94 ℃预变性4 min;94 ℃、1 min,55 ℃、1 min,72 ℃、1 min,收集荧光信号,30个循环。
真菌定量分析所用的引物[18]为5.8S:5- CGCTGCGTTCTTCATCG-3,ITSIF:5-CTT GG T CATTTAGAGGAAGTAA-3,PCR反应体系25 μL,其中模板DNA、SYBR Premix Ex Taq(TaKaRa Biotechnology)、5.8S引物、ITSIF引物、ddH2O等,分别为1、12.5、1、1、9.5 μL;PCR反应程序:95 ℃预变性5 s;94 ℃、30 s,53 ℃、30 s,72 ℃、30 s,收集荧光信号,40个循环。
放线菌定量分析所用的引物[19]为Act920F:5-TACGGCCGCAAGGCTA-3,Act1200R:5- TCRTCCCCACCTTCCTCCG-3,PCR反应体系25 μL,其中模板DNA、SYBR Premix Ex Taq(TaKaRa Biotechnology)、Act920F引物、Act1200R引物、ddH2O等,分别为1、12.5、1、1、9.5 μL;PCR反应程序:95 ℃预变性10 min;95 ℃、15 s,65 ℃、30 s,72 ℃、15 s,收集荧光信号,40个循环。
1.2.4 土壤真菌分离 采用传统的平板稀释涂布法分离菌株,分离获得的真菌菌株用马铃薯葡萄糖琼脂培养基(PDA)做成的固体斜面在4 ℃下保藏[20]。
1.2.5 真菌对百香果组培苗的侵染测定 将健康的百香果茎端(表面灭菌)移入经高温高压灭菌处理的培养基中心,每瓶1株,培养30 d后,挑选长势一致的百香果组培苗进行接种试验。无菌条件下,在离百香果组培苗茎端2 cm处接种一圈培养过夜的真菌菌液,15 d后,观察百香果组培苗长势,对照组以添加等量PDA培养基代替[21]。
1.2.6 真菌的鉴定 形态学鉴定。菌株的形态学鉴定以菌株的菌落及分生孢子形态为鉴定依据,将分离的菌株接种至PDA培养基中,28 ℃培养3 d,观察菌落形态,在显微镜下随机测量50个分生孢子的大小并记录其形态特征。
分子鉴定法。DNA提取采用生工生物工程(上海)股份有限公司购买的真菌基因组提取试剂盒。真菌鉴定的特异性引物为NS1:5-GTAG TC ATATGCTTGTCTC-3,NS6:5-GCATC AC AGA CCTGTTATTGCCTC-3,对菌株的18S rDNA进行扩增。PCR程序为:预变性94 ℃、5 min,循环94 ℃、30 s,55 ℃、35 s,72 ℃、1 min,35个循环,72 ℃,延伸8 min。采用2%的琼脂糖凝胶电泳检测PCR反应结果[20]。DNA提取和测序委托生工生物工程(上海)股份有限公司完成,将测得序列到NCBI数据库进行比对,获取同源性最高的微生物。
1.3 数据处理
采用Excel软件进行实验数据的统计与初步分析,采用SPSS软件进行数据的方差、显著性分析和相关性分析。
2 结果与分析
2.1 百香果根际土壤的理化性质分析
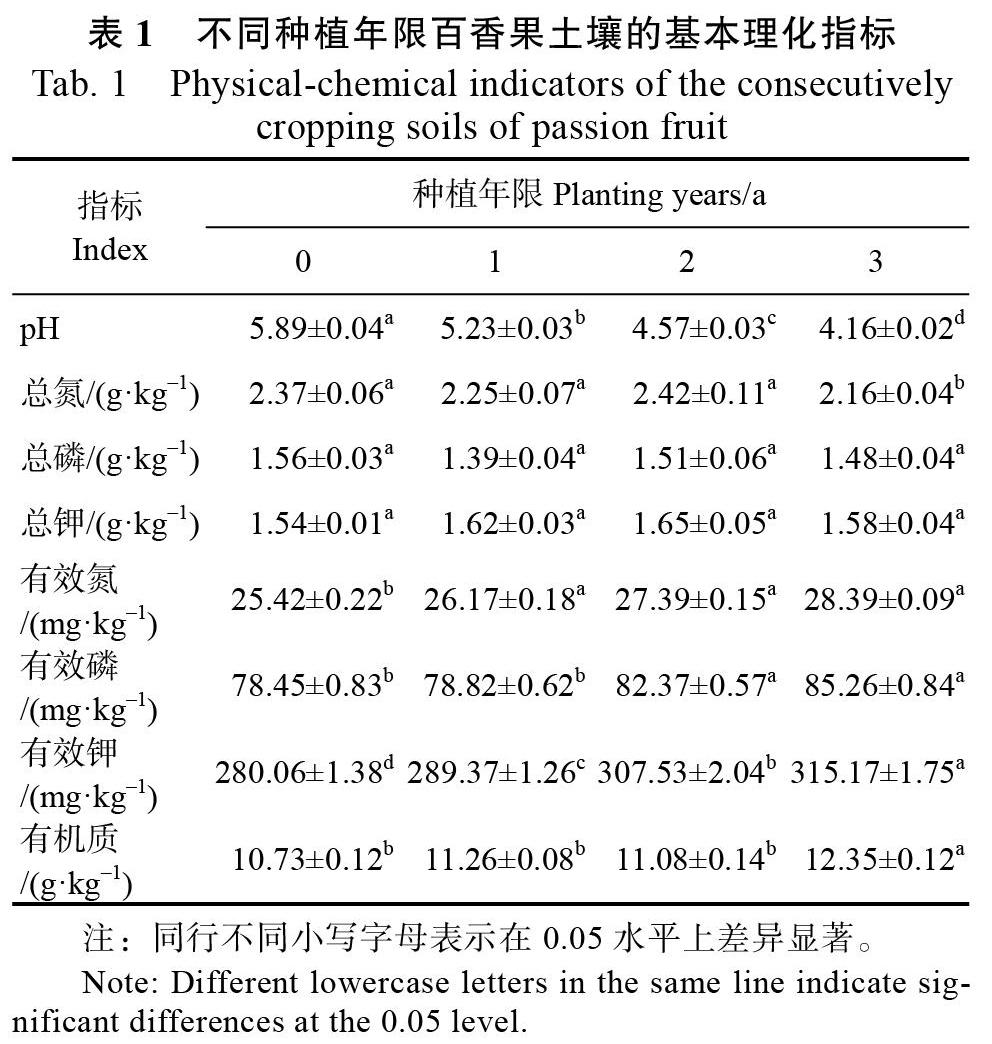
不同种植年限百香果根际土壤的理化指标分析结果表明(表1),土壤pH随着种植年限的增加,呈显著下降趋势。除总氮在第3年土壤有显著下降外,土壤总磷、总钾在不同种植年限间无显著差异。土壤有效氮、有效磷、有效钾及有机质含量呈上升趋势,以种植3 a的土壤最高。其中,种植1、2、3 a间的土壤有效氮含量无显著差异,但与对照间呈显著性差异;种植0 a和1 a间、2 a和3 a间土壤有效磷无显著差异,但种植0、1 a土壤与种植2、3 a年土壤之间呈显著性差异;种植0、1、2 a间土壤有机质含量无显著性差异,而与种植3 a的土壤呈显著性差异。其次,不同种植年限间,土壤有效钾含量均呈显著性差异。可见,连作百香果根际土壤理化性质的变化,主要表现为土壤酸化趋势明显,有效性养分含量增加。
2.2 百香果根际土壤的自毒潜力分析
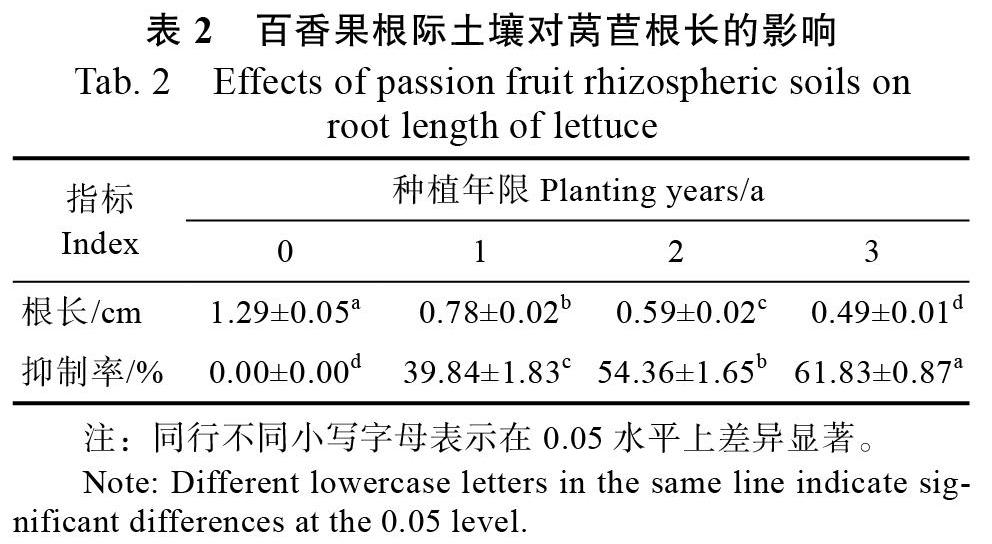
不同种植年限百香果根际土壤的自毒潜力分析结果表明(表2),随着种植年限的增加,百香果根际土壤对受体莴苣根长的抑制率呈显著上升趋势。与对照相比(0 a),种植1、2、3 a的百香果根际土壤对受体的抑制率分别为39.84%、54.36%、61.83%。可见,连作百香果根际土壤存在一定的自毒作用。
2.3 百香果根际土壤微生物数量分析
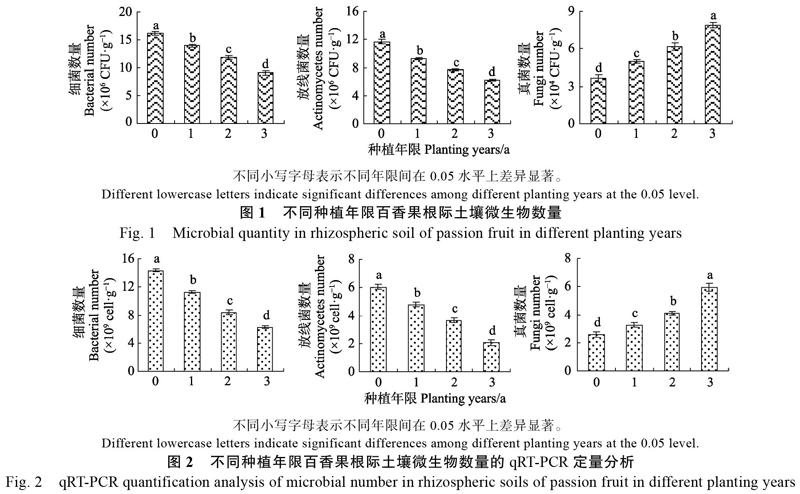
稀释涂抹平板计数法测定结果表明(图1),随着百香果种植年限的增加(0~3 a),土壤细菌与放线菌数量呈显著下降趋势,即细菌从16.18× 106 CFU/g下降到9.14×106 CFU/g,放线菌从11.65×106 CFU/g下降到6.24×106 CFU/g;而真菌数量则呈显著上升趋势,即从3.62×104 CFU/g上升到7.89×104 CFU/g。荧光定量PCR法测定结果表明(图2),随着百香果种植年限的增加(0~3 a),土壤细菌与放线菌数量依然呈显著下降趋势,真菌数量呈显著上升趋势。可见,随着百香果连作年限的增加,百香果根际土壤微生物数量发生变化,表现为细菌、放线菌数量逐年减少,真菌数量逐年增加。
2.4 土壤年限与莴苣根长、土壤理化指标及微生物数量的相关性分析
相关性分析结果表明(表3),百香果种植土壤的年限与pH、细菌、放线菌数量呈现极显著负相关,即随着土壤年限增加,该指标呈显著下降趋势;而与土壤有效氮、有效磷、有效钾含量及真菌数量呈显著或极显著正相关,即随着土壤年限的增加呈显著上升趋势。莴苣根长与土壤pH和细菌、放线菌数量呈显著正相关,而与真菌数量呈显著负相关。其次,pH与土壤有效氮、有效钾含量及真菌数量呈显著或极显著负相关,而与细菌、放线菌数量呈显著或极显著正相关。可见,随着土壤年限的增加,百香果根际土壤酸化加剧,土壤中有效性养分含量增加,土壤中的细菌数量下降,真菌数量上升,对莴苣的抑制作用增强。
2.5 百香果根際土壤真菌的分离、鉴定及侵染效果评价
以种植3 a的百香果根际土壤为材料,分离培养百香果根际土壤真菌,成功分离并纯化9种不同类型的真菌(图3),分别标记为BXG 1~BXG 9。将分离获得的真菌接种到百香果组培苗上,在无菌条件下进行培养与观察,发现与对照相比,菌株BXG 3、BXG 7、BXG 9感染后的百香果长势明显下降(图4)。可见,菌株BXG 3、BXG 7、BXG 9可影响百香果的正常生长。
进一步对菌株BXG 3、BXG 7、BXG 9进行鉴定,结果表明,BXG 3菌落白色,蓬松凸起,棉絮状,培养基反面为深黄色,具有大、小型分生孢子,大型分生孢子呈镰刀形或纺锤形,无色透明,大小为21.3~50.1 μm × 2.2~4.7 μm,小型分生孢子呈梨形或肾形,无色透明,大小为5.4~ 13.2 μm × 2.1~4.5 μm。BXG 3序列长度532 bp,與层出镰刀菌(Fusarium proliferatum)的同源性达99%。形态学与分子鉴定结果表明,BXG 3属于镰刀菌属的层出镰刀菌。BXG 7菌落絮状,羊毛状白色至浅灰色,培养基反面为浅茶色至黑色,有末端生厚壁孢子及间生厚壁孢子,大小分别为4.3~5.6 μm×5.4~7.6 μm和3.9~5.5 μm×5.2~7.1 μm。BXG 7序列长度541 bp,与棕黑腐质霉菌(Humicola fuscoatra)的同源性达99%。形态学与分子鉴定结果表明,BXG 7属于腐质霉属的棕黑腐质霉菌。BXG 9菌落呈圆形至贝壳状,有细长毛和绒毛,培养基反面为灰褐色至灰黑色至黑色,孢子无色,光滑,长椭圆形,大小为4.3~6.9 μm× 1.5~2.5 μm。BXG 9序列长度603 bp,与毛栓菌(Trametes hirsuta)的同源性达99%。形态学与分子鉴定结果表明,BXG 9属于栓菌属的毛栓菌。
3 讨论
土壤作为植物养分吸收及储藏的载体,土壤肥力指标的变化对于植物的生长、产量及品质具有一定的影响。本研究结果表明,随着百香果种植年限的增加,土壤的pH呈显著下降趋势,总氮、总磷、总钾等含量变化较小,有效氮、有效磷、有效钾及有机质含量呈上升趋势。相关性分析表明,百香果种植年限与pH呈极显著负相关,而与土壤有效氮、有效磷、有效钾含量呈显著或极显著正相关。据报道,作物连作极易导致土壤酸化,可提高土壤速效氮、磷、钾的含量,但也容易导致其流失,而对全氮、全磷、全钾影响则较小[22-24]。可见,随着百香果连作年限的增加,土壤酸化程度加剧,土壤中有效性养分增加。
土壤有效性养分增加对于植物的生长是有利的,然而本研究发现,随着百香果种植年限的增加,土壤对受体的抑制作用增强,自毒潜力加剧。Wang等[13]研究发现,随着百香果连作年限的增加,百香果产量和品质呈下降趋势。可见,虽然百香果连作土壤有效性养分增加,但受土壤酸化的影响,以及自毒潜力的增加,百香果对养分的吸收或利用效率下降,进而影响其生长。这种影响是否与土壤中的微生物存在一定的关系。据报道,中性偏碱的土壤环境较适合细菌、放线菌的繁殖与生长,而真菌则偏向于中性偏酸环境[25-26]。Latz等[27]研究发现,作物生物群落多样性提高,可促进作物根际土壤有益菌的生长,抑制土传病害传播。恶劣环境条件下,真菌比细菌具有更强的生命力[28],而连作极易导致细菌数量减少、真菌数量增加,进而导致地力衰竭,病虫害加剧[29]。本研究发现,随着百香果种植年限的增加,百香果根际土壤的细菌、放线菌数量呈下降趋势,真菌数量呈上升趋势。相关性分析表明,百香果种植年限与pH、细菌、放线菌数量呈极显著负相关,而与真菌数量呈显著或极显著正相关。土壤pH与土壤真菌数量呈显著负相关,而与细菌、放线菌数量呈显著或极显著正相关。可见,随着百香果种植年限的增加,百香果根际土壤酸度增强,土壤细菌、放线菌数量下降,真菌数量上升,土壤微生物从细菌型向真菌型转化,进而导致百香果生长发育不良。
在上述研究的基础上,进一步从种植3 a的百香果根际土壤分离真菌并对其侵染百香果能力进行评价,结果表明,共分离、纯化获得9个真菌菌株,将菌株回接种至百香果组培苗中进行侵染测定,发现被菌株BXG 3、BXG 7、BXG 9侵染后的百香果苗长势与对照相比明显下降。鉴定结果表明,菌株BXG 3、BXG 7、BXG 9分别为层出镰刀菌(Fusarium proliferatum)、棕黑腐质霉菌(Humicola fuscoatra)和毛栓菌(Trametes hirsuta)。据报道,层出镰刀菌是引起根腐病的病原菌,在染病的杂草黑燕麦中也曾分离到该病原菌[30-31]。高玉峰[32]研究连作蔬菜根际土壤真菌多样性时,从土壤中分离出61种病原真菌,其中包含了棕黑腐质霉菌。而毛栓菌则具有极强的木质素纤维降解能力,极易导致植物根系被快速降解,降低植物根系的生长能力[33-34]。由此可见,百香果连作后,土壤中存在的毛栓菌可快速降解和分离植物根系,降低根系的抵抗力,使其更容易让病原真菌——层出镰刀菌和棕黑腐质霉菌侵染,进而降低百香果对养分的吸收能力,导致百香果病害加剧,生长受阻。
综上所述,随着百香果连作年限的增加,土壤pH值降低,酸化程度加剧,土壤自毒潜力加强,土壤微生物从细菌型向真菌型转化,更利于土壤病原菌的繁殖与侵染,进而影响百香果的生长。
参考文献
Xiong F, Li X, Zheng L, et al. Characterization and antioxidant activities of polysaccharides from Passiflora edulis Sims peel under different degradation methods[J]. Carbohydrate Polymers, 2019, 218: 46-52.
Andrad F H A, Sousa A A, Araujo C S P, et al. Cattle manure and liquid biofertilizer for biomass production of yellow passion fruit seedlings[J]. African Journal of Agricultural Research, 2017, 12(17): 1430-1436.
Oliveira A B, Lopes M M A, Moura C F H, et al. Effects of organic vs. conventional farming systems on quality and antioxidant metabolism of passion fruit during maturation[J]. Horticultural Science, 2017, 222: 84-89.
Paul E A. The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization[J]. Soil Biology and Biochemistry, 2016, 98: 109-126.
Finn D, Kopittke P M, Dennis P G., et al. Microbial energy and matter transformation in agricultural soils[J]. Soil Biology and Biochemistry, 2017, 111: 176-192.
劉 丽, 徐明恺, 汪思龙, 等. 杉木人工林土壤质量演变过程中土壤微生物群落结构变化[J]. 生态学报, 2013, 33(15): 4692-4706.
Girvan M S, Campbell C D, Killham K, et al. Bacterial diversity promotes community stability and functional resilience after perturbation[J]. Environmental Microbiology, 2005, 7(3): 301-313.
Zhou X G, Wu F Z. P-coumaric acid influenced cucumber rhizosphere soil microbial communities and the growth of Fusarium oxysporum f. sp. cucumerinum Owen[J]. PLoS One, 2012, 7(10): e48288.
Li X G, Ding C F, Hua K, et al. Soil sickness of peanuts is attributable to modifications in soil microbes induced by peanut root exudates rather than to direct allelopathy[J]. Soil Biology and Biochemistry, 2014, 78: 149-159.
杨宇虹, 陈冬梅, 晋 艳, 等. 不同肥料种类对连作烟草根际土壤微生物功能多样性的影响[J]. 作物学报, 2011, 37(1): 105-111.
王增炎. 百香果栽培种植管理及病虫防治[J]. 农业开发与装备, 2017(10): 148-148.
周文杰, 吕德国, 秦嗣军. 植物与根际微生物相互作用关系研究进展[J]. 吉林农业大学学报, 2016, 38(3): 253-260.
Wang H B, Chen X T, Ding L, et al. Replant problem and soil toxicity of passion fruit (Passiflora edulis Sims) in China[J]. Allelopathy Journal, 2018, 44(1): 1-12.
鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000.
李 坤. 葡萄连作障碍机理及调控途径的研究[D]. 沈阳: 沈阳农业大学, 2010.
王海斌, 陈晓婷, 丁 力, 等. 土壤酸度对茶树根际土壤微生物群落多样性影响[J]. 热带作物学报, 2018, 39(3): 448-454.
顾 卿, 许艳丽, 魏 巍, 等. 土壤细菌群落密度实时荧光定量PCR检测体系的建立[J]. 土壤与作物, 2012, 1(2): 117-120.
Nicolas C P B, Richard C, Samuel D, et al. Validation and application of a PCR primer set to quantify fungal communities in the soil environment by real-time quantitative PCR[J]. PLoS One, 2011, 6(9): e24166.
Gregoris T B D, Aldred N, Clare A S, et al. Improvement of phylum-and class-specific primers for real-time PCR quantification of bacterial taxa[J]. Journal of Microbiological Methods, 2011, 86(3): 351-356.
张 奇, 梁友谊, 胡文文, 等. 稻田土壤中抑草真菌的筛选与鉴定[J]. 中国农学通报, 2015, 31(12): 210-216.
吴林坤. 根系分泌物介导下的连作地黄根际互作与作用机制研究[D]. 福州: 福建农林大学, 2014.
林 生, 庄家强, 陈 婷, 等. 福建安溪不同年限茶树土壤养分与微生物Biolog功能多样性的差异分析[J]. 中国生态农业学报, 2012, 20(11): 1471-1477.
王新超, 杨亚军, 陈 亮, 等. 不同品种茶树氮素效率差异研究[J]. 茶叶科学, 2004, 24(2): 93-98.
朱莜婧, 李晓明, 张 雪, 等. 低碳農业背景下提高肥料利用率的技术途径江[J]. 江苏农业科学, 2010(4): 15-17.
王孝国, 胡文革, 陈登稳, 等. 新疆艾比湖精河入湖口湖滨湿地土壤微生物区系研究[J]. 西南农业学报, 2012, 25(3): 972-976.
谢龙莲, 陈秋波, 王真辉, 等. 环境变化对土壤微生物的影响[J]. 热带农业科学, 2004, 24(3): 39-47.
Latz E, Eisenhauer N, Rall1 B C, et al. Plant diversity improves protection against soil-borne pathogens by fostering antagonistic bacterial communities[J]. Journal of Ecology, 2012, 100(3): 597-604.
Bardgett R D, Lovell R D, Hobbs P J, et al. Seasonal changes in soil microbial communities along a fertility gradient of temperate grasslands[J]. Soil Biology and Biochemistry, 1999, 31(7): 1021-1030.
陈 慧, 郝慧荣, 熊 君, 等. 地黄连作对根际微生物区系及土壤酶活性的影响[J]. 应用生态学报, 2007, 18(12): 2755-2759.
李 河, 周国英, 何末军. 油茶根腐病病原菌分子鉴定及其生物学特性研究[J]. 西南林学院学报, 2008(5): 45-48, 56.
朱海霞, 程 亮, 郭青云. 3株镰刀菌的鉴定、对野燕麦的致病力及其对5种作物的影响[J]. 中国生物防治, 2010, 26(S1): 84-89.
高玉峰. 河北保护地蔬菜土壤真菌多样性研究[D]. 杨凌: 西北农林科技大学, 2008.
李 江, 袁月祥, 闫志英, 等. 毛栓菌原生质体制备和再生及单核菌株产漆酶特性[J]. 菌物学报, 2012, 31(1): 102-109.
贾翠英, 张玉辉, 柴高波. 1株毛栓菌产纤维素酶酶学性质研究[J]. 河南农业大学学报, 2012, 46(6): 678-681.
责任编辑:谢龙莲
猜你喜欢
智慧少年·故事叮当(2019年11期)2019-12-09
智慧少年·故事叮当(2019年7期)2019-08-09
智慧少年·故事叮当(2019年6期)2019-06-28
智慧少年·故事叮当(2019年5期)2019-06-11
大自然探索(2019年1期)2019-01-24
知识就是力量(2017年12期)2018-01-15
大自然探索(2017年3期)2017-04-07
大自然探索(2017年3期)2017-04-07
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
