
剑豆主要农艺性状多样性研究与综合评价
2021-08-06 07:49刘明骞陈丽君周玮丁美美陈晓阳
热带作物学报 2021年2期
刘明骞 陈丽君 周玮 丁美美 陈晓阳


摘 要:剑豆为优良的绿肥和药食同源植物。本研究以来源于11个国家的19份剑豆种质为材料,采用R-3.6.2软件,通过相关性分析、主成分分析和聚类分析等方法分析剑豆22个农艺性状,并对其进行综合评价。结果表明:剑豆不同性状间差异均达到显著或极显著水平,遗传变异丰富,具较大的选择潜力;单株荚数和种子形态指数性状、叶形指数性状、地径、节间距之间呈极显著正相关关系,叶形指数性状和种子形态指数性状之间也呈极显著的正相关关系;主成分分析结果显示,前4个主成分累计贡献率已达86.201%,可代表22个性状的主要信息,若以产量为选择目标,应选择种子、叶面积、地径和节间距大且始花期晚的种质;聚类分析结果表明,19份剑豆种质可分为3类,分别为早熟不攀爬型、晚熟半攀爬型和晚熟善攀爬型,可根据不同用途和育种需求进行选择。
关键词:剑豆;农艺性状;多样性
Abstract: Canavalia ensiformis is an excellent green fertilizer and medicinal & edible homologous plant. 19 accessions of C. ensiformis from 11 countries were used in this study. R-3.6.2 software was adopted to analyze 22 agronomic traits through correlation analysis, principal component analysis and cluster analysis, and the comprehensive evaluation was made. The results showed that there were significant or extremely significant differences among different traits. There was an extremely significant positive correlation between pod number per plant and seed shape index traits, leaf shape index traits, ground diameter and pitch spacing, and there was also an extremely significant positive correlation between leaf shape index traits and seed shape index traits. The cumulative contribution rate of the first four principal components reached 86.201%, which could represent the main information of 22 traits. For yield, the germplasms with large seed, leaf area, diameter and pitch spacing and late flowering should be selected. Based on this, the germplasms could be divided into three categories, namely early-maturing non-climbing type, late-maturing semi-climbing type and late-maturing good climbing type, which could be selected according to different needs.
Keywords: Canavalia ensiformis; agronomic traits; diversity
剑豆(Canavalia ensiformis),为豆科刀豆属植物,广泛分布于亚洲、美洲、非洲等热带、亚热带地区,我国长江流域以南各省零星栽培。剑豆根系发达,适应能力强,耐贫瘠,可作为土壤改良和覆草植物[1-6]。剑豆种子不仅富含蛋白质、人体必需氨基酸和矿质元素等营养物质,还含有刀豆氨酸、刀豆球蛋白A、类黄酮等物质[7-9],《本草纲目》和《救荒本草》中均有其作为药物的记载,为卫生部公布的第一批药食兼用的植物[10],也被美国科学院认为是热带地区潜在的食物来源[11]。我国广西、湖南、四川等地区常将其嫩荚腌制后食用,某些地区也将剑豆种子粉末作为高蛋白饲料,添加至兔[12]、鱼[13-14]和鸡[15]等的饲料之中[16-17]。
植物种质资源是生产和遗传改良的物质基础,也是优良新品种(品系)选育与利用的源头。种质资源的收集、鉴定评价与创新利用是一项长期的基础性工作[18-19]。剑豆用途广泛,但国内外对于剑豆的研究大多集中于种子中凝集素的提取、药用功能的分析及种子营养物质含量等方面,剑豆的引种及种质评价方面研究较少。本研究以来源于11个国家的19份种质资源为研究对象,调查了主要生长性状、种实性状和生长节律,进行分析及综合评价,旨在为剑豆种质资源的收集、引种和育种材料的筛选、研究提供理论参考。
1 材料与方法
1.1 材料
试验材料为来源于澳大利亚热带作物和饲料研究中心(AusTRCF)、巴西、中国等地(表1)的19份剑豆种质资源。种植于广东省广州市华南农业大学教学科研基地,该地气候与环境特征详见文献[17]。
1.2 试验数据的采集
试验地采用完全随机区组设计,每小区(面积为5 m2)6株,5次重复,株行距為60 cm× 100 cm。于3月种植后进行平常的除草、灭虫等田间管理。于5—6月每日早晚各观察1次,记录初始开花时间,统计始花期[Flowering period(FP)/d]。于7—8月所有种质都已开花时,每份种质选取30轮共90片位于植株中部、向阳面且生长完好的叶片采用WinFOLIA Pro 2008a LEAF软件(Regent Instruments,Canada)采集叶形指数性状数据[20]:叶面积[Leaf area (LA)/cm2]、叶周长[Leaf perimeter (LP)/cm]、叶长[Leaf length (LL)/cm]、叶宽[Leaf width (LW)/cm]和叶宽长比[Ratio of width to length of leaf (RWLL)];测量后称重得其叶鲜重[Leaf fresh weight (LFW)/g]。于11月种子成熟时,统计各种质单株荚数[Yeild per plant (YPP)],用电子游标卡尺测量其地径[Root (Root)/mm]大小,用直尺测量主茎节间距[Internodal length (II)/cm]。每份种质分别取30个生长完好的豆荚,测量和统计豆荚相关性状:荚长[Pod length (PL)/cm]、荚宽[Pod width (PW)/cm]、荚厚[Pod thick (PT)/cm]、每荚粒数[Number of seeds per pod (NSP)],种子形状相关性状:种子长[Seed length (SL)/mm]、种子宽[Seed width (SW) / mm]、种子厚[Seed thick (ST)/mm]、种脐长[Hilum length (HL)/mm]、种子脐长比[Ratio of hilum length to seed length (RHS)]、种子宽长比[Ratio of width to length of seed (RWLS)]、种子厚宽比[Ratio of thick to width of seed (RTW)]及百粒重[100-seeds-weight (HSW)/g]等性状。
1.3 数据处理
对所得性状数据进行方差分析、相关性分析、主成分分析和系统聚类分析。试验数据均用Excel 2007软件记录、整理,R-3.6.2软件Psych、Hmisc、Corrgram、Corrplot、FactoMineR、Factoextra等数据包进行分析[21]。
2 结果与分析
2.1 主要性状的方差分析
对剑豆19份种质的豆荚、种子、叶片等22个性状进行调查,方差分析结果(表2)显示:
除地径的种质间差异达到显著(P<0.05)外,其他各性状在不同种质间的差异均达到极显著(P<0.01)。在所观测的性状中,单株荚数的变异系数最大,达45%,变幅为7.85~57.64;其次是节间距,变异系数为41%,变幅为5.82~22.02。变异系数最小的为叶宽/叶长和种子厚/种子宽,分别为6%和7%,变异幅度为0.57~0.70和0.57~ 0.73。说明剑豆不同种质间存在较大的差异,变异范围广,遗传变异丰富,选择育种潜力大,可根据不同需求,选择不同种质进行利用[22]。
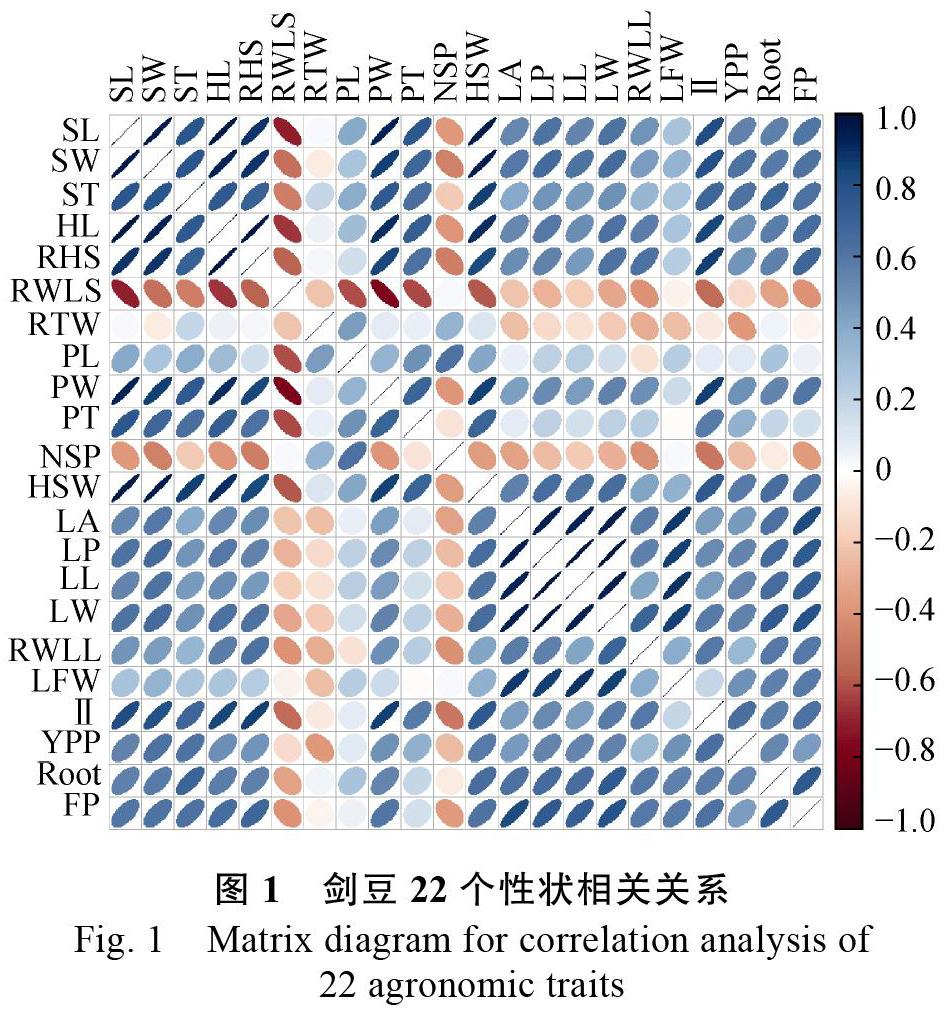
2.2 不同性状间相关性分析
通过Corrplot数据包对剑豆22个主要性状进行相关性分析,得到性状间相关关系(图1)。结果表明:单株产量和种子形态指数性状(种子长、宽、厚,种脐长,百粒重)、叶形指数性状(叶面积、叶周长、叶长、叶宽、叶鲜重)、地径、荚宽、节间距之间呈极显著正相关关系,与荚厚、叶干重之间呈显著正相关关系;叶形指数性状和种子形态指数性状之间也呈极显著正相关关系;荚厚和荚宽与种子指数性状间呈极显著正相关关系;节间距、始花期、地径和种子形态指数性状、叶形指数性状和荚形性状之间多呈极显著正相关关系。说明一般情况下,主茎的节间距越大,始花期时间越晚,其营养生长时间越长,地径也就越大,其叶片就越宽大,豆荚、种子也就越大,相应的产量就越高。说明如果以产量为选育目标,要着重注意对营养生长期长的种质的选择,其生长表现为地径较大,主茎节间距较长,叶面积大,开花较晚。剑豆地上部生物量与种子形态指数性状(种子长、宽、厚,种脐长,百粒重)、叶鲜重和单株结实量呈极显著的正相关关系,说明地上部生物量越大,产量越高。剑豆的生物量和产量可协同选择。
2.3 剑豆不同种质间性状主成分分析
主成分分析能在不损失信息量的前提下将多个可能相关的指标简化为几个独立的综合指标,使分类方法得以简化而易于操作。基于主成分分析的主成分聚类,能在二维和三维图上直观地体现出种质之间的遗传距离远近[23]。通过R-3.6.2软件FactoMineR、Factoextra等数据包进行主成分分析。结合主成分碎石图及主成分分析(表3)可以看出,前4个主成分即可代表原22个性状的主要信息。
前4个特征根累计贡献率已达86.201%。其中第1主成分贡献率达55.255%,通过不同性状对主成分的贡献度(图2)可以看出,其特征向量所凝聚的生物学信息主要是种子形态指数指标(种子的长、宽、厚和种脐的长度、种子的百粒重等)、叶形指数相关形态特征(叶面积、叶周长、叶长、叶宽等)以及与以上性状有极顯著正相关关系的荚宽、节间距、始花期、地径和单株产量。其结果与相关性分析相似,可以称第1主成分是产量相关构成因子。
第2主成分贡献率为15.657%,其特征向量所凝聚的生物学信息主要是叶鲜重和叶面积,因此可以称其为叶片质量构成因子。第3主成分贡献率为10.312%,其特征向量所凝聚的生物学信息主要是每荚粒数和荚长,其与产量相关性也较大。第4主成分的贡献率为4.977%,其特征向量所凝聚的生物学信息主要是单株荚数。
从产量因子考虑,第1主成分越大越好。若以产量为目标,应主要对第1主成分进行选择。
即选择种子大、叶片宽大、地径大、节间距大、始花期晚的种质。
2.4 不同剑豆种质的聚类分析
根据不同性状对各主成分的贡献度,重新计算19份种质的主成分得分,并依此进行聚类分析。通过Factoextra数据包采用组内误差平方和方法进行分析判断,19个剑豆种质可分成3个类群(图3)。
第一类群为早熟不攀爬型,包括PI 358592、PI 279593、PI 470242、NT 576和BN-13474-63、B1、B2、B3、CHN3、CHN4、210共11份种质,分别来源于埃塞俄比亚、菲律宾、印度尼西亚、苏丹、巴西、中国和巴拉圭。此类群植株为直立或半直立型,主干明显,不善攀援,分枝较多,属于早熟型。
第二类群为晚熟半攀爬型,包含CHN1、Tce 1、NT 3073、NT 3074、CPI 50103和CPI 58560共6份种质,分别原产于中国广东、尼日利亚、哥伦比亚和委内瑞拉,植株为半蔓生型,可以攀爬,主干粗壮,分枝较多,生物量大,结实多,且为晚熟型。
第三类群为晚熟善攀爬型,包含CHN2、PI 487475共2份种质,来源于中国广西和伊拉克。该类群主茎不明显,极善攀爬,节间距最大,叶片也比较宽大,种子大且为红色,生长快,结实量大,且为晚熟型。
3 讨论
通过表型性状进行种质资源鉴定和筛选更为方便和直观。表型是由环境和基因型共同调控的,在育种的整个进程中,表型监测不仅可对育种前期的室内种质筛选进行指导,而且能在后期推广种植中对品种的田间表现进行评估[21]。本研究以来源于11个国家的19份剑豆种质资源为研究对象,对22个表型性状的方差分析结果显示,各性状间的差异均达到显著或极显著水平。说明剑豆的不同种质间的基因交流非常低,可能与剑豆以自花授粉为主、异交率低有直接关联[1]。而且剑豆多为零星分布,地理距离较远,且其传粉媒介主要为蚂蚁和蜜蜂,活动范围有限,在一定程度上限制了杂交的可能。此外,剑豆的座果率很低,大约只有10%,更加限制了基因的流动[24]。可根据不同利用和育种需求,选择不同种质进行利用。
剑豆的生长性状(包括叶片的大小、节间距的长度、株高、地径等)与剑豆的主要繁殖性状(包括始花的早晚、荚果大小、种子大小、单株荚数等)之间的相关关系紧密。营养生长时间越长,植株越高大或藤越长,地上部生物量越大,则剑豆的结荚数一般也就越多,荚果和种子也就越大。剑豆地上部生物量与种子形态指数性状和单株结实量呈极显著正相关关系,说明地上部生物量越大,产量越高。剑豆的生物量和产量可协同选择。
主成分分析(PCA)是多元统计分析框架内的一种统计技术,可用来揭示数据中最重要的性状和品种之间的相似性,即用少数变量来反映原来变量的大部分信息[21]。主成分分析结果显示,前4个特征根累计贡献率已达86.201%。其中第1主成分贡献率达55.255%,其特征向量所凝聚的生物学信息主要是种子形态指数指标、叶形指数相关形态特征以及与以上性状有极显著正相关关系的荚宽、节间距、始花期、地径和单株产量。可以称第1主成分是产量相关构成因子。从产量因子考虑,第1主成分越大越好,要获得产量高的种质,可主要对第1主成分进行选择。即选择种子大、葉片宽大、地径大、节间距大、始花期晚的种质[18]。
聚类分析将19份剑豆种质资源划分为三大类群,各类群之间在品质性状上表现出明显的差异性和互补性,与刘明骞等[16]通过SRAP分子标记进行分析的结果相似。第一类群为早熟不攀爬型,植株为直立或半直立型,主干明显,分枝较多,属于早熟型。可用于收获种子、轮作(或间作)改良土壤。但其中种质210较为特殊,其豆荚短且直,其种色为黄褐色或深褐色,主干不明显,茎极细软,极善攀爬。第二类群为晚熟半攀爬型,包含的5份种质,皆属于半蔓生型,可以攀爬,分枝较多,生物量大,结实多且为晚熟型,适用于豆荚采食、林地覆盖、土壤改良和荒坡水土保持等。尤其是种质NT3073和CPI50103,生长期长,为优良覆草和土壤改良植物。第三类群为晚熟善攀爬型,只包含CHN2、PI 487475两份种质,该类群极善攀爬,叶片也比较宽大,主茎不明显,生长快,分枝多,可用作果实采集、庭院遮荫等,但因其节间距较大,藤蔓太长,易于林木缠结,不适于林地覆盖;该类群种子大且为红色,可制作工艺品(市场上称之为魔豆)。可根据不同利用或育种需求,选择不同种质资源。
参考文献
Morris J B. Swordbean (Canavalia ensiformis (L.) DC.) genetic resources regenerated for potential medical, nutraceutical and agricultural traits[J]. Genentic Resources and Crop Evolution, 2007, 54(3): 585-592.
Carlo Acosta S I. Promoting the use of tropical legumes as cover crops in Puerto Rico[D]. Mayaguez (Puerto Rico): University of Puerto Rico, 2009.
Liu M Q, Ding M M, Chen L J , et al. Genetic diversity and relationships among Canavalia ensiformis (L.) DC. Accessions as revealed by sequence-related amplified polymorphism markers[J]. Biochemical Systematics & Ecology, 2014, 57: 242-249.
Baligar V C, Fageria N K. Agronomy and physiology of tropical cover crops[J]. Journal of Plant Nutrition, 2007, 30(8): 1287-1339.
Ma L, Rao X, Lu P, et al. Acid-tolerant plant species screened for rehabilitating acid mine drainage sites[J]. Journal of Soils and Sediments, 2015, 15(5): 1104-1112.
Pohlan H A J, Janssens M J J, Eversbusch B G. Impact of Canavalia cover crop management in Coffea arabica L. on plant-invertebrate associations[J]. The Open Agriculcure Journal, 2008(2): 84-89.
Sasipriya G, Siddhuraju P. Evaluation of growth performance, serum biochemistry and haematological parameters on broiler birds fed with raw and processed samples of Entada scandens, Canavalia gladiata and Canavalia ensiformis seed meal as an alternative protein source[J]. Tropical Animal Health and Production, 2013, 45(3): 811-820.
Sridhar K R, Seena S. Nutritional and antinutritional significance of four unconventional legumes of the genus Canavalia-A comparative study[J]. Food Chemistry, 2006, 99(2): 267-288.
Siddhuraju P, Becker K. Species/variety differences in biochemical composition and nutritional value of Indian tribal legumes of the genus Canavalia[J]. Nahrung, 2001, 45(4): 224-233.
朱建标, 王洪新. 洋刀豆植物资源的综合开发利用[J]. 中国野生植物资源, 2002(3): 17-20.
Vadivel V, Janardhanan K. Diversity in nutritional composition of wild jack bean (Canavalia ensiformis L. DC) seeds collected from south India[J]. Food Chemistry, 2001, 74(4): 507-511.
Esonu B O, Udedibie A B I, Herbert U, et al. Comparative evaluation of raw and cooked jackbean (Canavalia ensiformis) on the performance of weaner rabbits on the performance of weaner rabbits[J]. World Rabbit Science, 1996, 3(4): 139.
Fagbenro O A, Adeparusi E O, Jimoh W A. Nutrient quantity of detoxified jackbean (Canavalia ensiformis L. DC) seeds cooked in distilled water or trona solution and evaluation of the meal as a substitute for soybean meal in practical diets for Nile tilapia, Oreochromis niloticus, Fingerlings[J]. Journal of Applied Entomology, 2007: 289-300.
Osuigwe D I, Obiekezie A I, Ogunji J O. Preliminary Evaluation of Jackbean (Canavalia ensiformis L. DC) Seed Meal as a Substitute for Fishmeal in diets for Clarias gariepinus[Z]. 2002.
Leon A, Vargas R E, Michelangeli C, et al. Detoxification of jackbean (Canavalia ensiformis L.) with pilot scale roasting. II. Nutritional value for poultry[J]. Animal Feed Science and Technology, 1998, 73(3-4): 231-242.
劉明骞, 陈丽君, 丁美美, 等. 基于SRAP分子标记的剑豆遗传多样性分析[J]. 中国农业大学学报, 2015, 20(2): 58-66.
刘明骞, 陈晓阳, 陈丽君, 等. 澳大利亚12个剑豆种源生长节律及主要农艺性状的遗传变异分析[J]. 广东农业科学, 2013, 40(23): 16-20.
顾文亮, 张建禹, 覃永兰, 等. 鹧鸪茶种质资源品质性状的多样性分析[J]. 热带作物学报, 2019, 40(12): 2364-2368.
潘映红. 论植物表型组和植物表型组学的概念与范畴[J]. 作物学报, 2015(2): 175-186.
董必成. 喜旱莲子草克隆片段的适应策略[D]. 北京: 北京林业大学, 2014.
孙维敏, 徐 阳, 龚榜初, 等. 浙江省柿种质资源果实形状多样性研究[J]. 林业科学研究, 2019, 32(5): 42-50.
殷冬梅, 张幸果, 王 允, 等. 花生主要品质性状的主成分分析与综合评价[J]. 植物遗传资源学报, 2011, 12(4): 507-512, 518.
郑本川, 崔 成, 李浩杰, 等. 长江流域甘蓝型油菜育种亲本农艺性状的遗传变异、相关及主成分分析[J]. 南方农业学报, 2019, 50(10): 2196-2204.
Smartt J. Evolution of grain legumes. II. Old and new world pulses of lesser economic importance[J]. Experimental Agriculture, 1985, 21(1): 1.
责任编辑:白 净
猜你喜欢
山东农业科学(2016年12期)2017-01-21
长江蔬菜·学术版(2016年12期)2017-01-12
求知导刊(2016年30期)2016-12-03
戏剧之家(2016年22期)2016-11-30
散文百家·下旬刊(2016年9期)2016-11-23
中国中药杂志(2016年20期)2016-11-19
人间(2016年27期)2016-11-11
农业与技术(2016年15期)2016-11-09
