
黄土丘陵区退耕草地群落优势种叶片光合生理对氮磷添加的响应
2021-08-06 10:24赖帅彬简春霞陈志飞周俊杰徐炳成
生态学报 2021年13期
赖帅彬,徐 升,简春霞,陈志飞,周俊杰,杨 全,王 智,,徐炳成,,*
1 西北农林科技大学 黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100 2 吉林农业科技学院, 吉林 132101 3 中国科学院水利部水土保持研究所, 杨凌 712100
近年来,大量矿物燃料燃烧以及农业中氮肥施用等致使大气氮沉降递增[1]。长期氮沉降在消除或缓解氮限制的同时,会造成陆地生态系统转为磷限制或氮磷共同限制[1-3]。氮素是蛋白质、核酸、酶和叶绿素等重要组分,是限制植物光合作用的关键因子[4]。磷是光合过程中核酸、ATP和磷脂等化学物质的主要成分,也是土壤中最易被固定和转化的元素,但即使向土壤中施加磷肥绝大部分会被有机物或金属离子固定,转化成缓效态和难效态,能为植物利用的有效磷非常少[5]。研究表明,氮限制下适宜添加氮能提高植物光能利用效率,促进植物生长和增加地上生物量[6],但当氮素超过某一阈值后,氮添加会转变为抑制植物生长[6-7]。磷素能提高扁豆根瘤固氮菌活性,从而增加叶片氮磷含量和提高光合速率[8]。因此土壤氮磷供应状况会对植物光合生理过程产生影响[9-10]。
不同植物由于其光合途径、养分敏感性和分配策略差异等导致其对氮磷添加响应存在差异[11-13]。光合氮利用效率(PNUE)和光合磷利用效率(PPUE)是表征植物叶片养分利用及光合生理特征的重要参数[14]。一般来说,C4植物的PNUE和PPUE高于C3植物,主要是C4植物的CO2浓缩机制使得1,5-二磷酸核酮糖羧化酶/加氧酶(ribulose-1,5-bisphosphate carboxylase/oxygenase,Rubisco)周围CO2达到饱和,使其在有效氮磷条件下拥有较高的净光合速率[11];另一方面,PNUE和PPUE的差异受叶片氮和磷分配影响[11]。氮素充足时,相比C3植物,C4植物分配少量N到光合酶中即可获得最大光合速率,将其余氮素分配到根茎部以获得较高生长速率[12]。相比禾本科植物,豆科植物将更多叶氮分配于光合系统,但禾本科植物在光合系统内部的氮分配机制更为有效[15]。由此可见,环境变化过程中,物种属性在决定其光合氮、磷分配及利用效率上发挥着重要作用。
退耕草地在保持水土、涵养水源、调节气候及防风固沙等方面具有重要生态保护作用[16]。对土壤养分贫瘠的退耕草地而言,合理均衡的氮磷添加可提升土壤养分含量,改变植株叶片氮磷含量,提高叶片光合能力,促进植被生长,加快草地恢复进程[7,9]。研究表明,内蒙古温带草原在适宜氮添加后羊草(Leymuschinensis)光合能力显著增强,地上生物量显著增加[7];适宜氮添加后,青海高寒草甸区禾本科植物净光合速率显著提升,生长促进明显[9],而随施氮量增加豆科植物叶片氮含量未发生变化但生物量逐渐下降[10]。因此,氮磷添加会对植物群落组成及各植物光合生理过程产生影响。
优势种在群落中占据着重要生态位,对群落结构和功能植被生态系统稳定起着至关重要的作用,旱区植被对环境条件变化的响应,主要取决于其优势种的响应和适应能力[17-18]。因此,明确主要优势种对氮磷添加的生理生态响应与变化特征,对揭示氮磷添加对退耕草地植被生产力和物种多样性的影响具有一定意义[18]。近年来,关于氮磷添加对退耕草地的效应研究主要集中在植物生长方面,而关于优势种在氮磷添加下光合生理生态研究较少[13,19]。白羊草(Bothriochloaischaemum)、长芒草(Stipabungeana)和达乌里胡枝子(Lespedezadavurica)为陕北黄土丘陵区草地群落中常见种,三者在该区退耕草地群落中均具有较广的分布幅度和占比[20],其中白羊草和长芒草分别为禾本科多年生C4和C3植物,达乌里胡枝子为豆科多年生C3半灌木。因此,本研究以这3种常见优势种为对象,通过测定分析3个优势种在不同配比氮磷添加下的光合生理参数、叶绿素荧光参数以及叶片氮磷含量等,探讨不同优势种光合生理对氮磷添加的响应特征与差异,以期为黄土丘陵区退耕草地恢复过程中的施肥管理与调控提供科学依据。
1 材料与方法
1.1 研究区概况
试验区位于陕西省延安市安塞区纸坊沟小流域,地处黄土丘陵沟壑区腹地,属暖温带半干旱大陆性季风气候。地理位置为东经109°13′46″—109°16′03″,北纬36°42′42″—36°46′28″,海拔为1041.5—1425.7 m。年均降雨量为507.2 mm(1951—2018),且季节分配不均,从全年降水量来看,4—10月占85%—95%,7—9月占60%—80%。年均气温8.8°C,辐射总量为493 kJ/cm2。土壤类型主要为黄绵土,植被区划上属暖温带半干旱森林草原区,处于落叶阔叶林区和草原区的过渡带。天然植被中以旱生草本植物群落为主,包括白羊草、长芒草、达乌里胡枝子、铁杆蒿(Artemisiasacrorum)、茭蒿(Artemisiagiraldii)等;有少量天然乔、灌木群落分布,主要由辽东栎(Quercuswutaishanica)、黄刺玫(Rosaxanthina)等组成。
1.2 试验设计
在纸坊沟流域,选择以白羊草、长芒草和达乌里胡枝子为优势种的退耕草地群落为试验样地,坡度为7°,坡向为东偏西46°。采用裂区试验设计,分别设置3个4 m×4 m 主区,区间由1 m宽未施肥缓冲区隔开,每个主区再分为4个2 m×2 m 的副区。主区为施氮处理,副区为施磷处理。根据黄土丘陵区年均氮沉降量(21.76 kg N hm-2a-1)[2],设置2倍和4倍的年均模拟氮沉降水平;参照刘海威等[21]的施磷标准(32.55 kg P2O5hm-2a-1),设置1倍和2倍的年施磷量,3个N添加处理分别为N0(0 kg N hm-2a-1)、N50(50 kg N hm-2a-1)和N100(100 kg N hm-2a-1),3个P添加处理分别为P0(0 kg P2O5hm-2a-1)、P40(40 kg P2O5hm-2a-1)和P80(80 kg P2O5hm-2a-1)。氮肥为硝酸铵钙[5Ca(NO3)2NH4NO310H2O],磷肥为重过磷酸钙[Ca(H2PO4)2H2O]。于2019年2月将上年地表所有枯枝落叶移除,在雨季来临前的6月6日,将提前称好的颗粒状氮肥和磷肥均匀撒入对应小区。
1.3 测定指标及方法
1.3.1光合生理特征
采用CIRAS- 2光合仪(美国PP System 公司),在无风晴朗天气里连续3天依次测定白羊草(抽穗期)、长芒草(开花期)和达乌里胡枝子(开花前期)的光合生理特征(每天完成一个优势种的光合生理指标测定)。本次测定于2019年7月19—21日09:00—11:30进行。每种植物测定时选取生长状况一致的植株,相同叶位且完全展开的成熟叶片,3个生物学重复。测定项目包括:净光合速率(Pn,μmol m-2s-1)、蒸腾速率(Tr,mmol m-2s-1)、气孔导度(Gs,mmol m-2s-1)、胞间CO2浓度(Ci,μmol/m)等。测定时使用自带红蓝光源(光强设置为1200 μmol m-2s-1),叶温设为25℃,相对湿度为60%,大气CO2浓度为380 μmol/mol。3种植物单叶面积均小于光合仪叶室面积,因此在测定时,使用佳能D7500相机对叶室内测定叶片拍照,后期在室内用Adobe Photoshop将图片处理后,再用Image-J计算出实际光合叶面积,将求得叶面积值输入光合软件(CIRAS- 2 remote control software)进行换算。叶片瞬时水分利用效率(WUEi,μmol/mmol)=Pn/Tr[22];气孔限制值(Ls)=1-Ci/Ca[23]。
1.3.2叶绿素荧光参数
采用连续激发式植物效率分析仪Handy PEA(英国 Hansatech 公司),在测定光合气体交换参数后,选择同一植株测定叶绿素荧光参数。测定前用暗适应夹夹住叶片暗适应30min(夹叶片时避开叶脉,当叶片太小时,将两叶片并排以填满暗适应夹圆形孔隙)。测定指标包括叶绿素初始荧光(Fo)、最大荧光(Fm)、PSⅡ最大光化学效率(Fv/Fm)和PSⅡ潜在活性(Fv/Fo)[24],每处理3次重复。
1.3.3叶片氮、磷含量
在叶片光合和荧光参数测定结束后,迅速采集同一株植物相同叶位5—6枚叶片装于自封袋中,带回实验室后将叶片平铺于A4纸上,用佳能750D相机垂直拍照并用Image-J计算叶片面积后,将叶片置于105℃烘箱杀青15min,75℃烘至恒重(0.0001g)。叶片采用MM400高通量组织研磨仪(Retsch,德国)研磨,经H2SO4-HClO4消煮后使用全自动凯氏定氮仪(FOSS- 8400,丹麦)测定叶片氮含量(Nmass)。采用钼蓝比色法(UV- 2600 紫外可见分光光度计,日本)测定叶片磷含量(Pmass)。叶片单位面积氮含量[Narea,g(N) m-2]和单位面积磷含量[Parea,g(P) m-2]分别为单位质量叶片氮含量(Nmass,g/kg)和叶片磷含量(Pmass,g/kg)与比叶面积(SLA)的比值,其中SLA为叶面积与叶片干重之比。光合氮利用效率[PNUE,μmol(CO2) g(N)-1s-1]和光合磷利用效率[PPUE,μmol(CO2) g(P)-1s-1]分别为净光合速率(Pn)与Narea和Parea的比值[11]。计算公式分别为:
Narea=Nmass/SLA
(1)
Parea=Pmass/SLA
(2)
PNUE=Pn/Narea
(3)
PPUE=Pn/Parea
(4)
1.4 数据分析
利用单因素方差分析(One-way ANOVA)分别检验各指标(光合气体交换参数、荧光参数、光合氮和磷利用效率)在不同氮磷添加处理下的差异显著性(P=0.05)。利用双因素方差分析(Two-way ANOVA)分别检验氮磷添加的交互作用对各优势种指标影响的差异显著性。采用线性模型拟合净光合速率与PSⅡ最大光化学效率和PSⅡ潜在活性以及净光合速率与叶片氮含量和叶片磷含量间的关系。采用SPSS 20.0处理数据,OriginPro 2016绘图。
2 结果与分析
2.1 光合生理特征
由图1可知,单独氮添加下,白羊草和达乌里胡枝子的Pn相比未施肥显著升高(P<0.05)。白羊草的Pn在N50和N100处理间差异不显著,达乌里胡枝子的Pn在N100下显著高于N50处理(P<0.05),长芒草的Pn仅在N100下显著高于未施肥处理。单独磷添加下,仅达乌里胡枝子的Pn较未施肥显著增加,且P40和P80处理间差异显著。在N50和N100处理下,磷添加对白羊草和长芒草的Pn无显著影响。施磷显著提高了达乌里胡枝子的Pn,N50处理下P40和P80处理间差异显著,而N100下P40和P80处理间差异不显著。氮磷添加对白羊草的Pn交互作用显著,而对长芒草和达乌里胡枝子无显著影响。
单独氮添加下,达乌里胡枝子的Tr相比于未施肥显著增加,且N100显著高于N50处理(P<0.05);白羊草和长芒草的Tr仅在N100下显著高于未施肥处理(P<0.05),分别为后者的2.13和1.54倍。单独磷添加下,白羊草的Tr无显著变化,长芒草和达乌里胡枝子的Tr在P80和P40处理下均显著降低(P<0.05)。N50下,白羊草、长芒草和达乌里胡枝子的Tr均随施磷量增加显著增加,其中白羊草和长芒草的Tr在P40和P80处理间差异不显著,而达乌里胡枝子的Tr在各处理间差异显著(P<0.05);N100下,仅达乌里胡枝子的Tr施磷后显著增加,且P40和P80处理间差异显著(P<0.05)。氮磷添加对3个优势种的Tr交互作用均显著(P<0.05)(图1)。
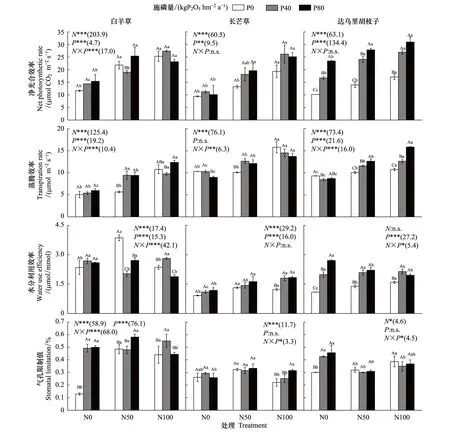
图1 不同氮磷添加处理下三种优势植物叶片净光合速率、蒸腾速率、水分利用效率和气孔限制值Fig.1 Leaf net photosynthetic rate (Pn), transpiration rate (Tr), water use efficiency (WUE) and stomatal limiting value (Ls) of the three dominant species under different N and P addition treatmentsN0表示0 kg N hm-2 a-1;N50表示50 kg N hm-2 a-1;N100表示100 kg N hm-2 a-1; 柱上不同大写字母表示同一施氮处理下施磷处理间差异显著,不同小写字母表示同一施磷处理下施氮处理间差异显著(P<0.05);括弧内数字为不同氮磷处理下的F值;*,**和***分别表示P≤0.05,P≤0.01和P≤0.001
单独氮添加下,三个优势种的WUEi相比未施肥处理显著升高(P<0.05)。白羊草和长芒草的WUEi在N50和N100处理间差异不显著,达乌里胡枝子的WUEi在N100处理下显著高于N50处理(P<0.05)。单独磷添加下,仅达乌里胡枝子的WUEi较未施肥处理显著提升,且P80和P40处理间差异显著(P<0.05)。N50处理下,磷添加对长芒草的WUEi无显著影响,显著降低了白羊草的WUEi,显著提升了达乌里胡枝子WUEi。N100处理下,磷添加显著提升了长芒草和达乌里胡枝子的WUEi,但P40和P80处理间差异不显著;白羊草的WUEi在P40处理下显著提升,但P80处理下显著降低。氮磷添加对白羊草和达乌里胡枝子的WUEi交互作用显著,对长芒草的WUEi无显著影响(图1)。
单独氮添加下,白羊草的Ls相比未施肥处理显著增加(P<0.05)。白羊草的Ls在N50和N100处理间差异不显著,长芒草的Ls在N50处理下显著高于N100处理(P<0.05),达乌里胡枝子的Ls仅在N100处理下显著增加。单独磷添加下,仅白羊草的达乌里胡枝子的Ls相比未施肥显著增加(P<0.05),且均在N50和N100处理间差异不显著。N50处理下,磷添加对长芒草和达乌里胡枝子的Ls无显著影响,白羊草的Ls仅在P80处理下显著增加;N100处理下,磷添加对达乌里胡枝子的Ls无显著影响,白羊草的Ls仅在P40处理下显著增加,长芒草的Ls仅在P80处理下显著增加(P<0.05)。氮磷添加对三个优势种的Ls交互作用显著(图1)。三个优势种的Pn与Nmass均呈显著正相关关系,仅长芒草和达乌里胡枝子的Pn与Pmass呈显著正相关关系(图2)。
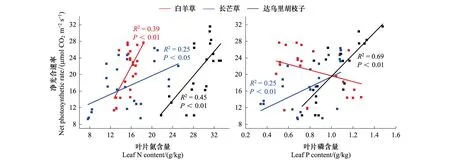
图2 不同氮磷添加处理下三种植物叶片净光合速率与叶片氮磷含量关系Fig.2 Relationships between net photosynthetic rate with Leaf N and P contents of the three dominant species
2.2 叶绿素荧光特征
单独氮添加下,白羊草和达乌里胡枝子的Fo相比未施肥处理显著降低(P<0.05),长芒草的Fo相比未施肥处理显著增加(P<0.05);白羊草的Fv/Fm和Fv/Fo仅N100下显著高于未施肥处理(P<0.05),而长芒草和达乌里胡枝子的Fv/Fm和Fv/Fo在各处理间差异不显著。单独磷添加下,白羊草和达乌里胡枝子的Fo相比未施肥处理显著降低(P<0.05),其中白羊草的Fo在P80处理下显著低于P40处理,达乌里胡枝子的Fo在P40和P80处理间差异不显著(P<0.05);白羊草的Fv/Fo相比未施肥处理显著增加,且P40和P80处理间差异不显著,而Fv/Fm仅在P40处理下显著高于未施肥处理(图3)。
N50下,磷添加显著降低了白羊草的Fo,显著增加了达乌里胡枝子的Fv/Fo;白羊草的Fo在P40和P80处理间差异显著,长芒草和达乌里胡枝子的Fo仅在P80处理下显著降低;白羊草的Fv/Fm和Fv/Fo仅在P40处理下显著增加,长芒草的Fv/Fm仅在P80处理下显著增加,达乌里胡枝子的Fv/Fo在P40和P80处理间差异不显著(P<0.05)。N100下,磷添加显著降低了白羊草和达乌里胡枝子的Fo,显著增加了长芒草和达乌里胡枝子的Fv/Fm和Fv/Fo;白羊草和达乌里胡枝子的Fo在P40和P80处理间差异不显著,长芒草的Fo仅在P80处理下显著增加;在P40和P80处理间,长芒草和白羊草的Fv/Fm和Fv/Fo和达乌里胡枝子的Fv/Fm差异不显著,达乌里胡枝子的Fv/Fo差异显著。氮磷添加仅对三个优势种的Fo和白羊草的Fv/Fm、达乌里胡枝子的Fv/Fo交互作用显著(图3)。
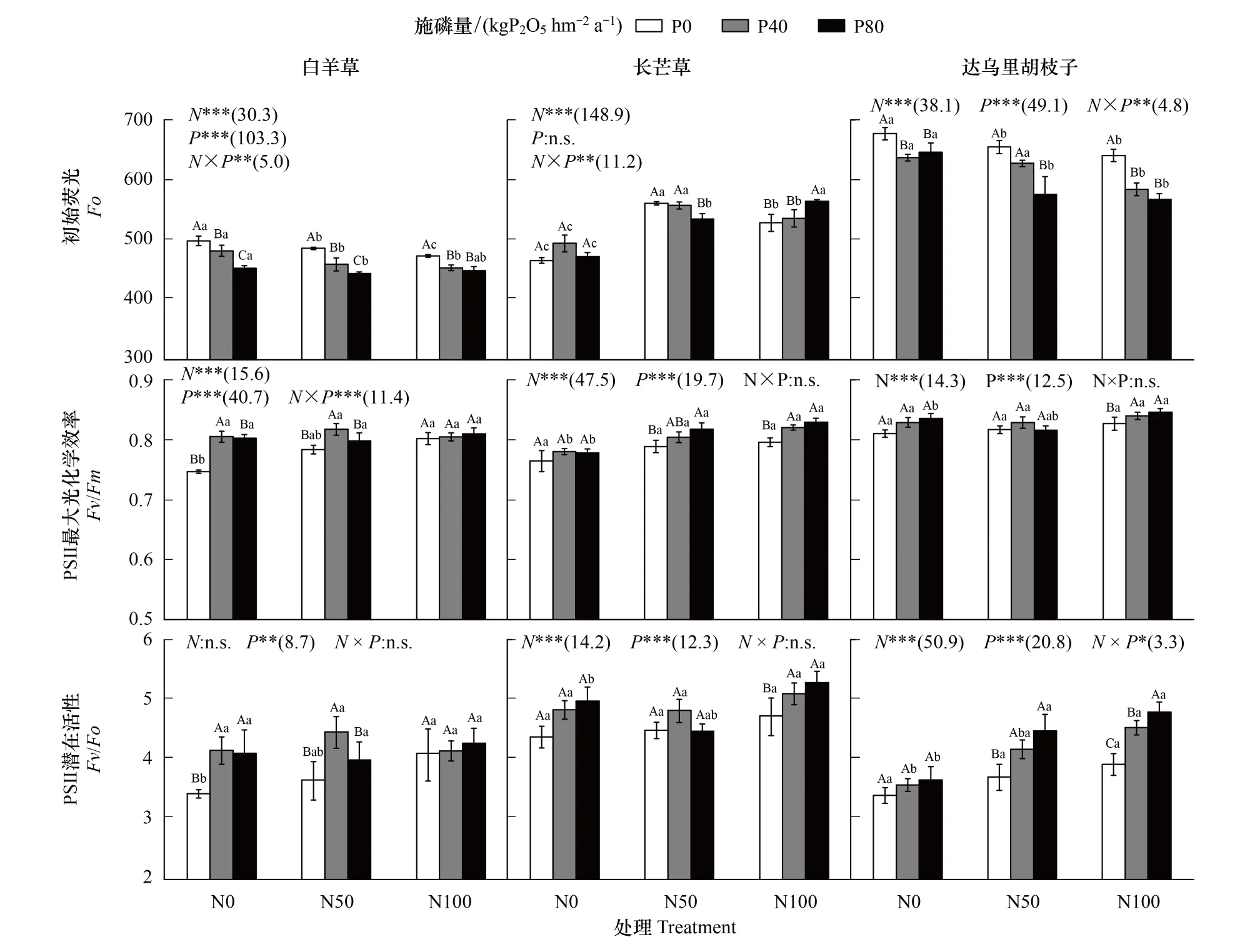
图3 不同氮磷添加处理下三种优势植物初始荧光、PSⅡ最大光化学效率和PSⅡ潜在活性Fig.3 Initial fluorescence, PSⅡ maximum photochemical efficiency and PSⅡ potential activity of the three dominant species under different N and P addition treatments
相关性分析表明,三个优势种的Pn与Fv/Fm呈显著正相关(白羊草:R2=0.31,P<0.01;长芒草:R2=0.35,P<0.01;达乌里胡枝子:R2=0.63,P<0.01),其中长芒草和达乌里胡枝子的Pn—Fv/Fm斜率明显高于白羊草。三个优势种仅长芒草和达乌里胡枝子的Pn与Fv/Fo呈显著正相关(长芒草:R2=0.33,P<0.01;达乌里胡枝子:R2=0.70,P<0.01),其中达乌里胡枝子的Pn—Fv/Fo斜率高于长芒草(图4)。
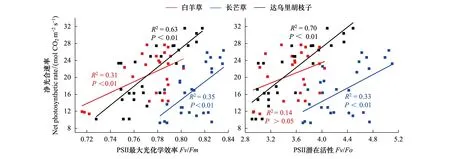
图4 不同氮磷添加处理下三种植物叶片净光合速率与PSⅡ最大光化学效率和PSⅡ潜在活性关系Fig.4 Relationships between maximum net photosynthetic rate with PSⅡ maximum photochemical efficiency and PSⅡ potential activity of the three dominant speciesFv/Fm:光系统Ⅱ最大光化学效率 PSⅡ maximum photochemical efficiency;Fv/Fo:光系统Ⅱ潜在光化学活性 PSⅡ potential activity
2.3 PNUE和PPUE
单独氮添加下,白羊草和达乌里胡枝子的PNUE、白羊草和长芒草的PPUE相比未施肥处理显著提升,长芒草的PNUE相比未施肥处理显著降低(P<0.05)。白羊草在N100下的PNUE显著高于N50(P<0.05),长芒草和达乌里胡枝子的PNUE在N50和N100间无差异;白羊草和长芒草的PPUE在N50和N100间差异显著,达乌里胡枝子的PPUE仅在N50下显著增加。单独磷添加下,长芒草的PNUE和白羊草的PPUE相比未施肥处理显著降低(P<0.05)。长芒草的PNUE仅在P80处理下显著降低,而达乌里胡枝子的PNUE仅在P80处理显著增加;白羊草的PPUE在P40和P80间无差异,长芒草的PPUE在P40下显著增加而在P80下显著降低,达乌里胡枝子的PPUE仅在P80处理显著增加(图5)。
N50下,磷添加后长芒草的PNUE显著增加,而白羊草的PPUE显著降低。长芒草的PNUE在P40和P80间无差异,白羊草和达乌里胡枝子的PNUE仅在P80下显著增加;白羊草的PPUE在P40和P80间无差异,达乌里胡枝子的PPUE仅在P40下显著增加。N100下,磷添加后仅达乌里胡枝子的PNUE显著增加。达乌里胡枝子的PNUE在P80下显著高于P40,长芒草的PNUE仅在P40处理下显著增加;白羊草的PPUE仅在P40下显著增加,长芒草的PPUE仅在P80下显著降低。氮磷添加对三优势种的PNUE和白羊草的PPUE交互作用显著,对长芒草和达乌里胡枝子的PPUE无影响(图5)。
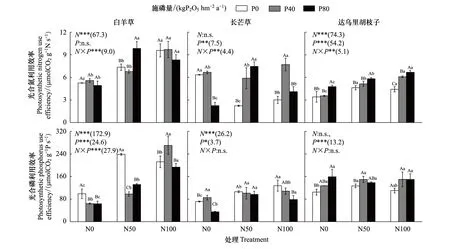
图5 不同氮磷添加处理下三个优势植物叶片光合氮和光合磷利用效率 Fig.5 Leaf photosynthetic N and P utilization efficiency of the three dominant species under different N and P addition treatments
3 讨论
气体交换参数反映植物光合速率变化的内在特征[6],其中Pn是表征植物光合作用大小的直接指标,是植物形成生物量的关键。植物种生理属性影响Pn对氮磷添加的响应[6,25-26]。本研究中,单独的氮添加下,白羊草和达乌里胡枝子在N50和N100处理下的Pn均显著高于未施肥对照,且白羊草的Pn在两个氮添加处理间差异不显著,而长芒草的Pn仅在N100处理下显著高于未施肥对照,表明禾本科C4植物白羊草在较低氮添加下即可达到较高的Pn,这可能是白羊草的CO2富集机制使其具有较高的Rubisco活性所致[12]。N50和N100处理下,达乌里胡枝子的Pn显著增加且各处理间差异显著,这可能是氮添加后改变了其光合氮与非光合氮分配比例[25](图5)。单独磷添加下,仅达乌里胡枝子的Pn显著增加,说明具有固氮能力的达乌里胡枝子对磷素添加更为敏感[10]。氮添加提高了禾本科和豆科植物的Pn,磷添加仅显著提升了豆科植物的Pn,这与两种功能型物种对土壤氮磷有效性变化的敏感度有关[7,10]。不同物种的Pn与叶片氮磷含量之间的相关性存在差异[27-28]。本研究结果表明,白羊草的Pn与Nmass呈正相关关系但与Pmass不相关,达乌里胡枝子和长芒草Pn与Nmass和Pmass均呈显著正相关关系(图2),说明单独的氮或磷添加,通过提高优势种叶片氮或叶片磷含量增加了Pn[6-7]。白羊草的Pn与Pmass无相关性,这可能是由于作为C4植物白羊草具有更高的Rubisco酶效率因而拥有较高的PPUE,分配较少的磷于光合组分中即可拥有较大的Pn[11,29]。相比于长芒草,达乌里胡枝子的Pn与Pmass的相关性更强,这可能是由于磷素能够促进达乌里胡枝子结瘤和提高其根瘤固氮酶活性,增加了叶片氮和光合能力[8]。
合理的氮磷添加能调节植物光合和蒸腾速率,进而影响植物WUEi[6,25]。本研究中,单独氮添加下长芒草和达乌里胡枝子的Tr相比于Pn增幅较小,引起WUEi显著提升;白羊草WUEi仅在N50处理下显著提升,N100处理下白羊草WUEi降低源于蒸腾速率增幅较大。氮添加下,白羊草较高的WUEi是由于其相对较高的Pn与相对较低的Tr所致,其较低Tr与C4植物较高的叶肉阻力和气孔阻力有关[30]。单独磷添加下达乌里胡枝子WUEi的显著提升归因于Pn的增幅较高(图1)。氮添加下,三个优势种的Ls增加;磷添加下仅白羊草和达乌里胡枝子Ls显著增加;氮磷添加下三个优势种Pn和Tr无统一变化规律,表明氮磷添加通过影响蒸腾速率和气孔限制因素改变植物光合能力[31-32]。
氮磷添加在影响植物光合作用的同时,对植物荧光特征产生影响[6]。研究表明,适宜氮磷添加常伴随PSⅡ反应中心光化学效率和潜在活性的提高,即Fo降低和Fv/Fm与Fv/Fo的增加[6]。本研究中,单独氮或磷添加下,仅白羊草的Fo显著降低而Fv/Fm和Fv/Fo值均显著增加,长芒草和达乌里胡枝子的Fv/Fm和Fv/Fo虽增加但均不显著。说明C4植物白羊草的PSⅡ反应中心对氮磷添加的敏感性高于C3植物长芒草和达乌里胡枝子,这可能是由于C3和C4植物类囊体膜的光化学和非光化学过程存在差异[33-34]。白羊草在P40处理下的Fv/Fm和Fv/Fo与P0的差异显著,但与P80差异不显著,说明施磷过高可能对白羊草光能转化和传递能力产生抑制作用[27]。Pn与Fv/Fm和Fv/Fo的拟合结果表明(图3),适宜氮或磷添加增强了各优势种PSⅡ反应中心光化学效率和潜在活性,提高了光合能力[6]。本研究中,氮磷共同添加下三个优势种Pn、PSⅡ活性及长芒草的WUEi高于单独氮或磷添加,双因素分析也显示氮磷共同添加对光合生理特征参数有交互作用(图1),说明氮磷共同添加在提高三种植物光合生理特征参数方面优于单独氮或磷添加[35-37]。
PNUE和PPUE是表征植物叶片生理特征和氮磷利用及分配的重要参数[14-15]。合理氮磷添加可优化氮磷分配能力,从而提高植物对环境的适应性[6,25]。本研究中,N50和N100处理下白羊草和达乌里胡枝子的PNUE和PPUE增加,且白羊草的PNUE和PPUE在两个氮添加处理下分别为达乌里胡枝子的1.58、2.17和1.88、1.92倍,说明适当氮添加提高了两个优势种光合氮利用效率[38],相比达乌里胡枝子,白羊草具有更高效的氮磷分配机制[15]。N50和N100处理下长芒草的PNUE显著降低,是由于其Pn相比于Narea增幅较小,说明禾本科C3和C4植物对氮磷分配策略存在差异,即白羊草分配更多氮于植物光合系统中,而长芒草则将更多的氮分配于光合系统以外部分(如细胞壁)[9,15]。磷添加下仅达乌里胡枝子的PNUE和PPUE在P80处理下相比于未施肥处理显著增加,这归因于磷添加后达乌里胡枝子的Pn显著增加[15]。氮或磷添加下各优势种PNUE和PPUE差异表明,白羊草和达乌里胡枝子将更多氮素分配到光合系统内部,而长芒草将更多氮素分配于光合系统以外部分,这与物种对资源利用策略有关[15,32]。
氮磷配施下,N50处理下,随着施磷量增加三个优势种PNUE逐渐增加,且两个禾本科植物增加幅度大于豆科植物达乌里胡枝子,其中白羊草和长芒草的PNUE在N50P80处理下已达最大值;N100处理下,三个物种的光合能力相比于N50处理下未显著增加,且白羊草和长芒草的PNUE在P80处理下出现下降(图5)。从光合能力和养分利用效率来看,N50P80是提高优势种光合能力和养分利用效率的最佳组合。氮磷施肥后植物光合氮分配策略的改变与三个优势种对氮磷敏感性差异有关[15],这种差异对增强草地群落适应外部环境条件变化的稳定性具有重要作用。从群落多样性和稳定性方面考虑,氮磷共同添加更有利于退耕草地植被恢复。
4 结论
氮或磷添加下,3个优势种通过增加叶片氮或磷含量,活化了气孔调节能力和PSⅡ活性,提高了光合能力,其中白羊草的光合气体交换参数和叶绿素荧光参数对氮添加的敏感程度高于长芒草和达乌里胡枝子,达乌里胡枝子光合气体交换参数对磷添加的敏感程度高于其余两个优势种。叶片氮含量增加是白羊草净光合速率上升的主要原因,而长芒草和达乌里胡枝子的净光合速率的提升则由叶片氮和磷含量增加所致。适当氮添加显著提高了白羊草和达乌里胡枝子的光合氮利用效率以及三个优势种的光合磷利用效率,三种植物光合生理特征对氮磷添加的响应与物种属性和施肥水平有关。N50P80是提高优势种光合能力和养分利用效率的最佳组合。上述结果表明,适当氮磷添加有利于提高半干旱区草地群落优势种的光合能力,这是施肥提高草地生物量的生理基础;草地群落优势种光合生理特征对氮磷添加响应差异是其对环境变化不同步性的生理表现,不同优势种在光合生理和养分利用方面的差异性表现对退耕草地恢复过程中维持群落多样性和稳定性具有重要意义。
猜你喜欢
小哥白尼(野生动物)(2021年3期)2021-07-21
语文世界(小学版)(2020年11期)2020-01-08
孩子(2019年8期)2019-08-26
中国民族民间医药(2018年13期)2018-07-31
小学阅读指南·低年级版(2017年4期)2017-04-24
大灰狼(2016年9期)2016-10-13
语文世界(小学版)(2016年1期)2016-02-17
中国民族医药杂志(2015年11期)2015-07-12
