
不同功能群及先锋种在高寒人工草地建植初期的作用
2021-08-03 13:37:14李兰平张慧敏李宏林梁德飞武学霞李希来
草地学报 2021年7期
李兰平, 张慧敏, 李宏林, 梁德飞, 武学霞, 李希来*
(1.青海师范大学青海省自然地理与环境过程重点实验室, 青海 西宁 810016; 2.青海大学省部共建三江源生态与高原农牧业国家重点实验室, 青海 西宁 810016; 3.青海大学农牧学院, 青海 西宁 810016)
人工草地是在人工措施下完全或部分破坏原有天然草地植被的基础上,通过综合的农业培育技术措施所建立的人工草地群落,其产量可达天然草地的10~20倍[1-2]。建设优质高效的人工草地与半人工草地是解决我国优质饲草料严重短缺,进而遏制过度放牧引起的草地大面积退化、生态功能衰退等生态安全问题的重要途径[2]。混播人工草地通常比单播人工草地具有更高更稳定的生产力、更好的牧草品质和更强的土壤改善作用[3-4],因此混播人工草地相关研究也引起了众多学者的关注。目前我国混播人工草地相关研究在草地建植和收获管理的具体措施上取得了较多成果,但这些研究大多以豆科植物和少数几个禾本科牧草混播组合为对象,对于更多的包括不同功能群组合在内的草种配置研究较为缺乏[2]。
功能群被定义为对特定环境因素有相似反应的一类物种,它是基于生理、形态、生活史或其它对某一生态系统过程相关以及与物种行为相联系的一些生物学特性来进行划分的[5]。目前研究发现功能群与草地生态系统生产力和稳定性均有一定的关系,如白永飞等[6]发现内蒙古高原针茅草原群落中起重要作用的植物功能群与群落初级生产力及其稳定性呈正相关关系;王长庭等[7]发现不同类型草地中,植物功能群的数量和对群落生产力所起的作用不同;刘士辉[8]则认为在不同群落,功能群与初级生产力之间呈单峰曲线或正相关关系等不同关系模式。虽然目前对植物功能群在天然草地生态系统中的作用有一定的研究,但不同功能群物种在高寒人工草地建植中的作用尚未见报道。先锋种是在植物群落演替早期出现的物种,往往具有生长迅速、种子产量大、传播能力强等特点[9]。先锋种可改善微环境、提高土壤养分并为次生物种提供一定的保护[10-11],因而在森林的恢复及公路护坡上有一定的应用[12-13],但其在人工草地建植中的作用尚有待验证。
在牧区,建植人工草地的主要目的是用于饲用。禾本科(Gramineae)植物大多为优质牧草,豆科(Leguminosae)植物则可提高牧草蛋白含量及减少氮肥施用量。因此以生产为目的的混播人工草地主要为禾草混播草地和豆禾混播草地。目前,学术界逐渐开始关注草地生态系统的多功能性[14-15],生物多样性与生态系统功能及其维持具有密切的关系[14]。增加人工草地草种配置的多元化,将非禾本科与非豆科植物纳入人工草地建设有助于提高人工草地物种多样性,进而影响草地的多功能性。本文通过不同物种组合混播试验,研究不同功能群及先锋种植物在高寒人工草地建植中的作用;本试验首次将非禾本科、非豆科植物用于高寒人工草地建植,以期为高寒人工草地建设中不同功能群物种及先锋种植物的选择提供一定的依据。
1 材料与方法
1.1 研究区概况
试验样地位于青海省黄南藏族自治州河南蒙古族自治县克其合滩(34°41′07″ N,101°46′02″ E),属于黄河源区,海拔高度为3 743 m。该地区属高原亚寒带湿润气候区,年平均气温0℃,年平均降水量580.1 mm,降水主要集中在5月至10月;全年日照时数为3 241.8 h[16]。研究区草地类型是高寒草甸,退化斑块约占30%,处于中度退化阶段,建群种为矮生嵩草(Kobresiahumilis)和高山嵩草(Kobresiapygmaea),优势物种包括密花香薷(Elsholtziadensa)、细叶亚菊(Ajaniatenuifolia)和西藏微孔草(Microulatibetica)等[17]。
1.2 试验设计
本试验在克其合滩原生植被相对较少区域经铲车移除地上草皮和翻耕后设置样区。为防止高原鼠兔(Ochotonacurzoniae)进入样地,我们在样地安装了防鼠围栏,防鼠围栏由地面以上 0.6 m防鼠镀锌铁皮(型号:35)和地面以下0.5 m防鼠钢纱网(网孔0.8 cm)组成,钢纱网和镀锌铁皮用铆钉连接,每隔2 m用1.4 m长的角铁固定,防鼠围栏外围安装普通围栏防止家畜进入[17]。
试验按照不同物种数量进行组合,即从所选的22种物种(主要为本土物种,表1)中随机抽取1,3,6,9,12,15种物种进行组合,组合数量分别为7,11,13,6,6,6,共计49个组合和1个对照[17-18]。参照王长庭[7]、刘育红[19]、Hector[20]等人研究,我们将这些物种和样地中出现的物种分为禾本科、豆科、杂类草和先锋种,其中所选物种中的先锋种为播娘蒿(Descurainiasophia)和菥蓂(Thlaspiarvense)[21-22]。播种组合中包含禾本科、豆科、杂类草和先锋种的组合数量分别为33,29,38,23。试验重复3次,小区总数量为150,大小为2 m×2 m,间距为1 m,所有小区按随机区组设计布置。播种量为5 g·m-2,组合内各物种按种子重量等量混匀。当组合物种数量为1时,该物种播种量为5 g·m-2;组合物种数量为3时,组合内各物种播种量为1.67 g·m-2;组合物种数为6时,各物种播种量为0.83 g·m-2;以此类推,组合物种数为15时,各物种播种量为0.33 g·m-2。于2018年6月初进行撒播播种,播种完成后覆盖无纺布一层。样地全年禁牧,所有小区均未进行施肥、灌溉等处理。

表1 播种物种
1.3 群落数据收集
于2018年和2019年8月中下旬进行群落数据收集,具体方法为在各小区内设置0.5 m×0.5 m样方,记录样方内植物群落盖度、出现的物种及其分盖度、多度和高度,其中各物种高度测量其3株个体,不足3株按实际株数测量。之后将样方中植物样品齐地面剪取,装入档案袋,85℃烘干至恒重后称量。相对盖度、相对多度和相对高度用于计算各物种的重要值,其公式为:重要值=(相对高度+相对盖度+相对多度)/3。
1.4 数据处理与分析
根据播种组合中是否包含禾本科、豆科、杂类草、先锋种对各人工草地植物群落地上生物量、盖度、群落高度和非播种物种数进行方差分析,并用禾本科、豆科、杂类草和先锋种种子播种量、重要值与地上生物量、盖度、群落高度、非播种物种数进行线性回归分析。禾本科、豆科、杂类草和先锋种种子播种量为其在组合中所含物种种子播种量之和,禾本科、豆科、杂类草和先锋种重要值为其在群落中所出现物种重要值之和。用Pearson相关性分析分析不同功能群和先锋种重要值与其种子播种量之间的关系。采用R 3.3.5进行回归分析,SPSS 17.0进行方差分析、相关性分析,SPSS 17.0和Excel 2016制图。
2 结果与分析
2.1 禾本科、豆科、杂类草和先锋种植物对人工草地植物群落的影响
方差分析表明,2018年禾本科植物显著增加人工草地群落地上生物量、盖度、高度和减少非播种物种数(P<0.05),而豆科、杂类草和先锋种植物对群落地上生物量、盖度、群落高度和非播种物种数均无显著影响(图1)。2019年禾本科植物依然显著增加人工草地群落地上生物量、群落高度和减少非播种物种数(P<0.05),但对群落盖度无显著影响(图2)。豆科和杂类草均显著降低人工草地群落地上生物量和群落高度(P<0.05),对群落盖度和非播种物种数无显著影响(图2)。先锋种植物对人工草地群落性状无显著影响(图2)。
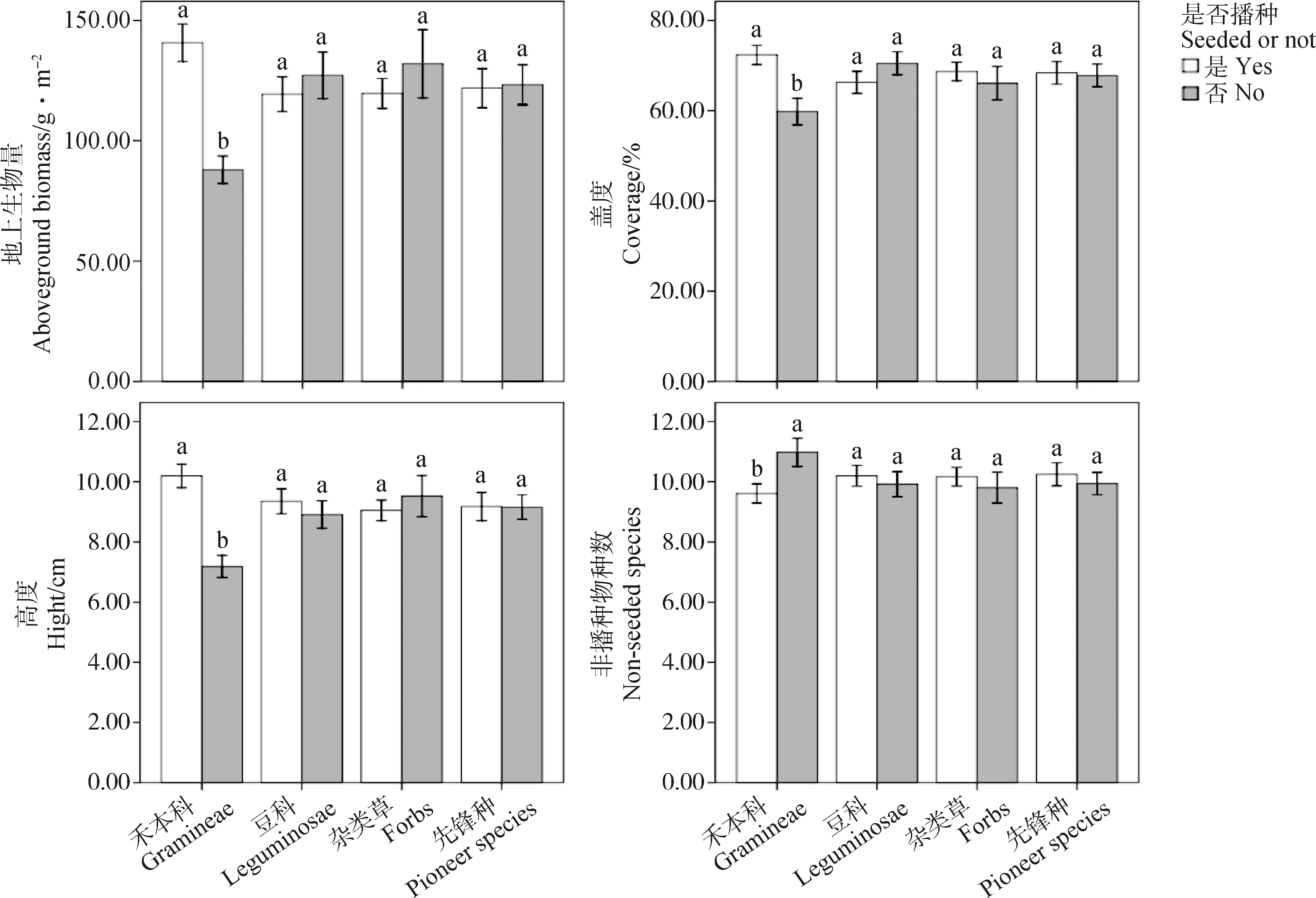
图1 2018年播种组合是否包含各功能群对群落性状的影响以及播种组合是否包含先锋种对群落性状的影响
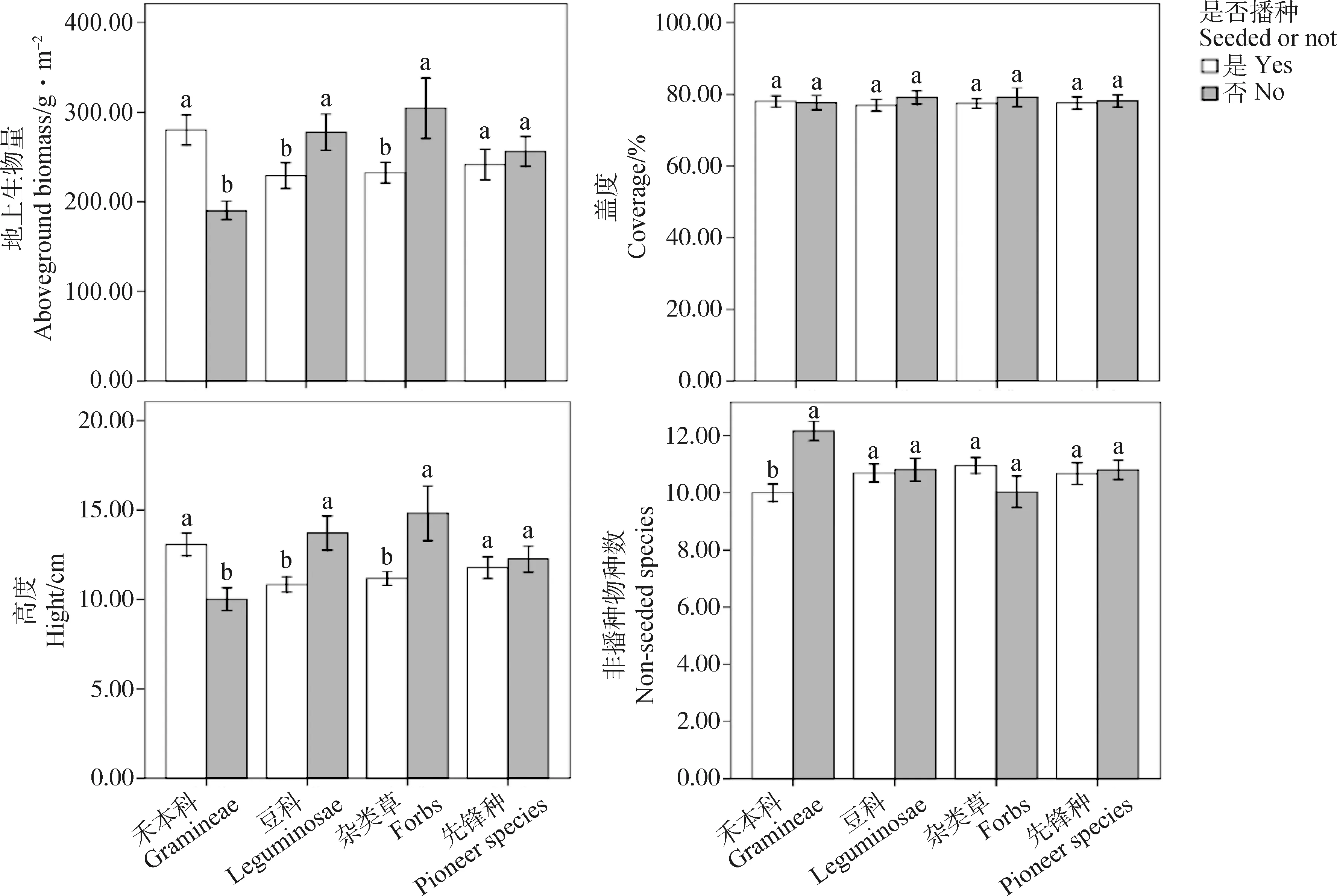
图2 2019年播种组合是否包含各功能群对群落性状的影响以及播种组合是否包含先锋种对群落性状的影响
2.2 禾本科、豆科、杂类草和先锋种种子播种量对人工草地植物群落的影响
通过线性回归分析,进一步分析不同功能群和先锋种种子播种量对人工草地建植的影响。结果表明,禾本科和杂类草种子播种量可较好地解释人工草地植物群落性状的差异,而豆科和先锋种种子播种量对人工草地植物群落性状的影响较弱或不显著(图3,图4)。具体为增加禾本科种子播种量,在人工草地建植的第一和第二年均可在一定程度上提高群落地上生物量(P<0.01)和群落高度(P<0.01),同时减少非播种物种数(P<0.05),但只在第一年增加群落盖度(P<0.01,图3,图4)。增加豆科种子播种量则会在建植第一年略微降低群落盖度(P<0.01,图3),第二年略微降低群落地上生物量(P=0.05)和群落高度(P<0.05,图4)。增加杂类草种子播种量在人工草地建植第一和第二年均小幅减少群落地上生物量(P<0.05)和群落高度(P<0.01),在第二年促进非播种植物的入侵(P<0.01,图3,图4)。增加先锋种种子播种量对人工草地群落性状无显著影响。回归分析结果与方差分析结果大体一致。
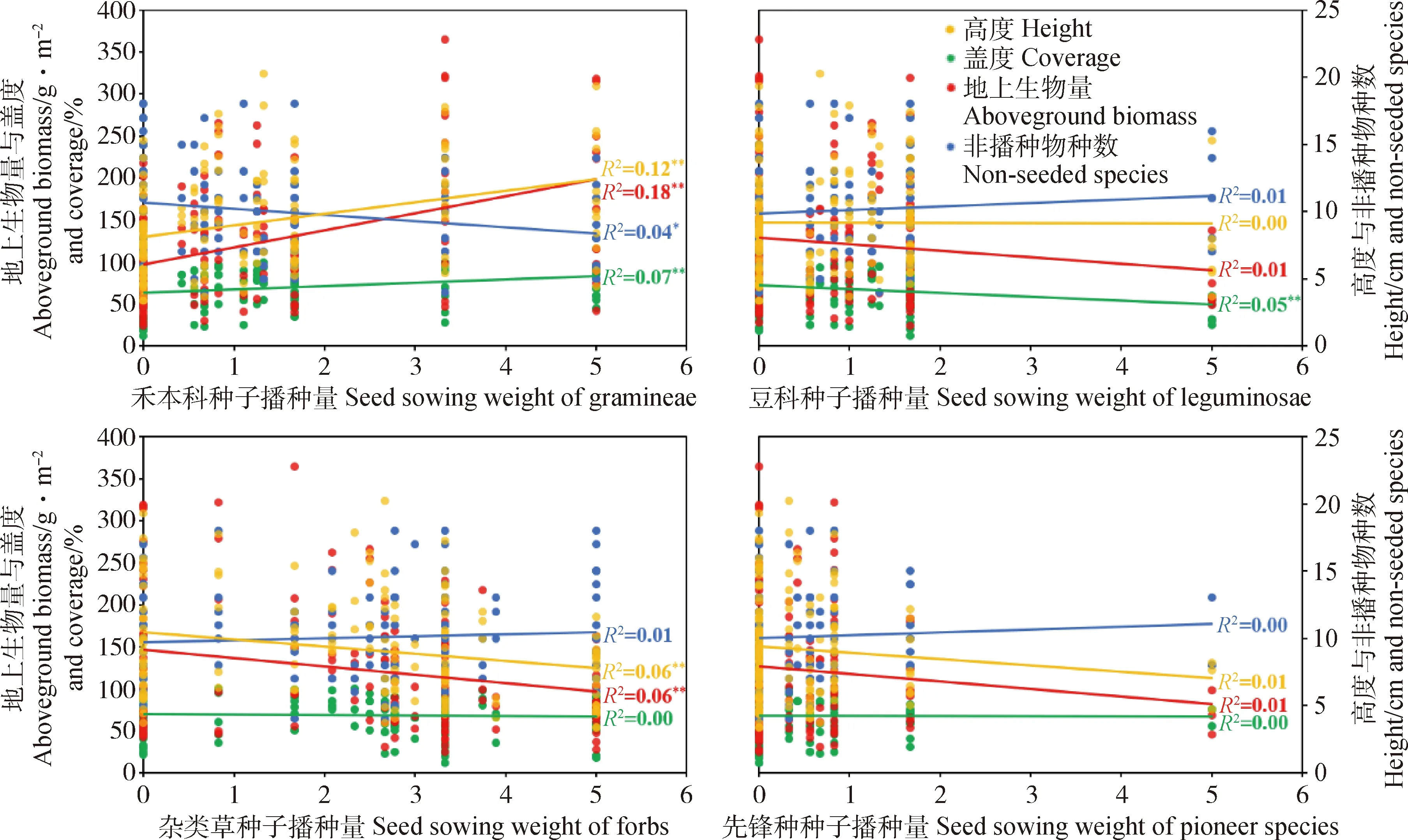
图3 2018年各功能群和先锋种种子播种量与群落性状回归分析结果
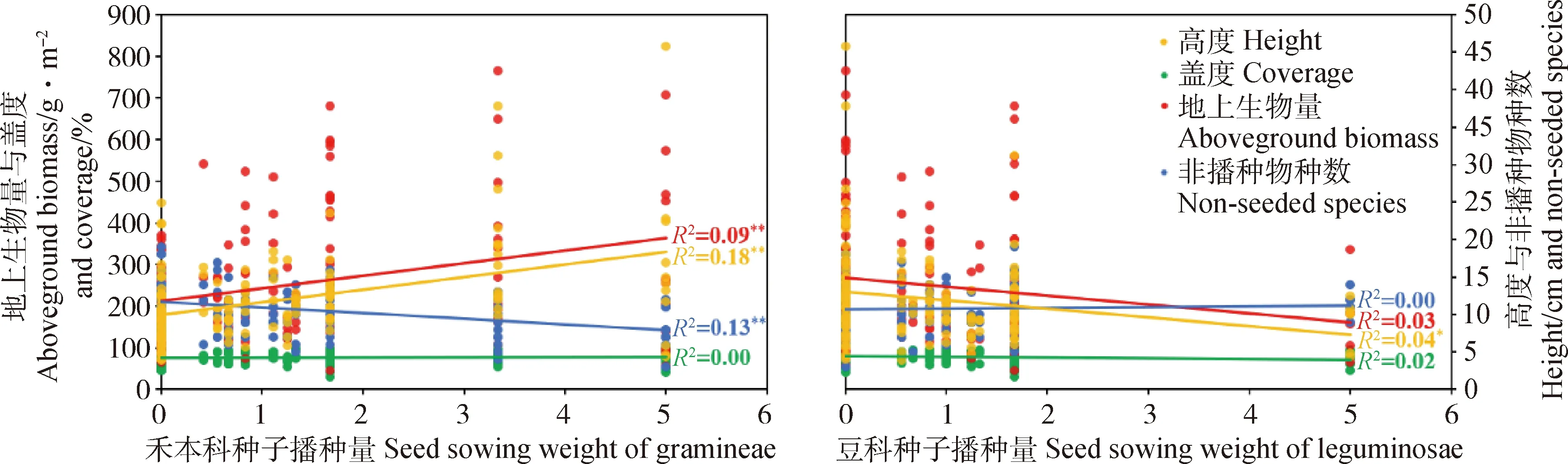
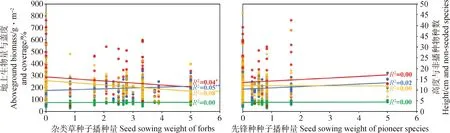
图4 2019年各功能群和先锋种种子播种量与群落性状回归分析结果
2.3 禾本科、豆科、杂类草和先锋种重要值对人工草地植物群落的影响
禾本科和杂类草重要值对于人工草地植物群落性状差异的解释程度较高,而豆科和先锋种重要值对人工草地植物群落性状的影响非常有限(图5,图6)。禾本科重要值增加可大幅提高群落地上生物量(P<0.01)和高度(P<0.01),同时减少非播种物种数(P<0.01,图5,图6),在人工建植第一年可在一定程度上提高群落盖度(P<0.01,图5),第二年对盖度的影响消失(图6)。豆科重要值增加会导致群落地上生物量(P<0.05)和高度略微下降(P<0.01,图6),对群落盖度和非播种物种数没有显著影响(图5,图6)。杂类草重要值增加会显著降低群落地上生物量(P<0.01),高度(P<0.01)和第一年群落盖度(P<0.01),但可以显著提高非播种物种数(P<0.01,图5,图6)。先锋种重要值增加对群落性状影响较弱,具体为在建植第二年略微降低群落地上生物量(P<0.01)和高度(P<0.05,图6),在建植第一年小幅增加群落盖度(P<0.05)、减少非播种物种数(P<0.01,图5),第二年小幅降低群落盖度(P<0.05)、增加非播种物种数(P<0.05,图6)。
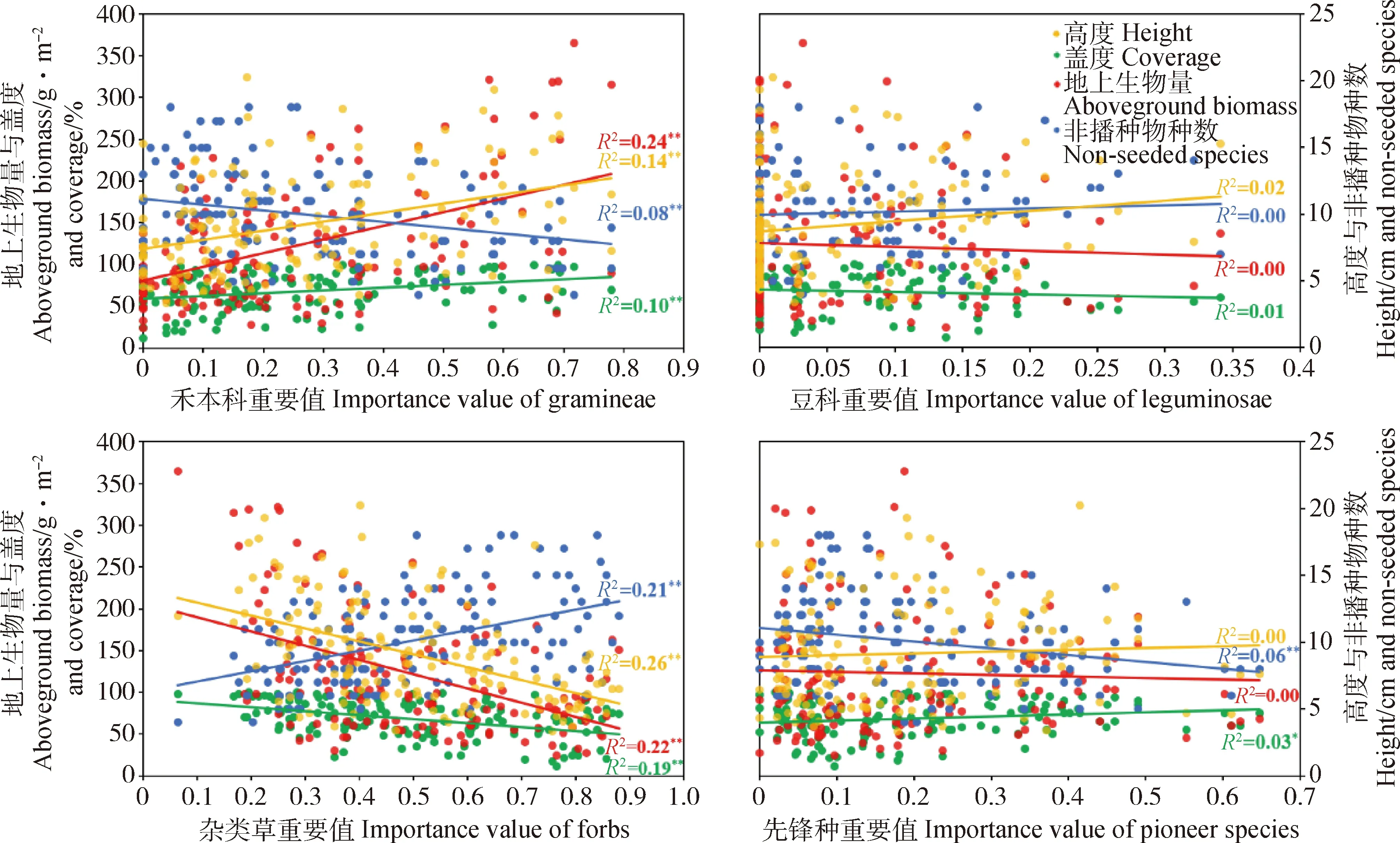
图5 2018年各功能群和先锋种重要值与群落性状回归分析结果
2.4 禾本科、豆科、杂类草和先锋种种子播种量与其重要值的关系
2018年各功能群种子播种量和先锋种种子播种量与其重要值呈显著正相关关系(P<0.01,表2)。在建植第二年,各功能群种子播种量与其重要值的正相关关系强度变弱(P<0.01),先锋种种子播种量与其重要值相关性不显著(表3)。

表2 2018年各功能群、先锋种种子播种量与其重要值相关性

表3 2019年各功能群、先锋种种子播种量与其重要值相关性
3 讨论
总体而言,不同功能群及先锋种植物在人工草地建植中的作用并不一致,其作用随建植年限有一定的变化。禾本科与杂类草在高寒人工草地建植初期对植物群落性状的影响较大,其中禾本科植物可形成较高的群落盖度、高度和地上生物量,同时显著减少非播种物种的出现,这与禾本科植物在人工草地建植初期提高群落地上生物量、抑制一二年生植物和多年生杂类草生长以及草地生产力主要取决于禾本科植物的结论一致[23-24]。杂类草的主要影响是降低群落高度和地上生物量,但可显著增加非播种物种数,这可能是由于试验所选用的大部分杂类草植物生长发育相对缓慢。豆科和先锋种在高寒人工草地建植初期的作用相对较弱,虽然对群落盖度、高度、地上生物量和非播种物种数有一定影响,但该影响非常有限。
增加混播组合中禾本科种子所占比例对人工草地建植初期植物群落有较大影响,增加杂类草与豆科种子比例对群落的影响次之,先锋种种子所占比例未表现出对群落的影响。其中,增加禾本科植物种子所占比例能进一步提高群落盖度、高度和地上生物量,减少非播种物种数。在建植第二年,虽然包含禾本科物种的组合与未包含禾本科物种的组合在群落盖度上没有显著差异,但是包含禾本科物种的组合具有更高的群落高度,因而也获得了更高的地上生物量。包含禾本科物种的组合在建植第二年依然能够显著减少群落中的非播种物种。因此,禾本科植物对于非播种物种的抑制作用是持续的,该抑制作用可能会持续到禾本科植物重要值下降到一定程度之后才会消失[22]。增加杂类草与豆科植物种子所占比例则会降低群落盖度或高度,进而导致群落地上生物量的下降。但是,增加杂类草植物种子播种量能提高群落中非播种物种的数量,有助于提高人工草地植物群落的物种多样性。将禾本科与杂类草进行组合是同时兼顾高寒人工草地生产力和物种多样性的方案。该方案在目前人工草地往往会导致草地物种多样性大幅下降以及人们对草地生态系统多功能性关注度持续提高的背景下具有一定的实践意义[14-15,25]。因此,在建设以生产为主要目的的人工草地时,应优先考虑禾本科植物。主要考虑草地多功能性时,可选择禾本科植物和杂类草植物进行组合。
各功能群与先锋种重要值对于群落地上生物量、盖度、高度和非播种物种数的影响与其种子播种量的影响基本一致。对群落影响最大的依然是禾本科的重要值,杂类草重要值的影响次之。豆科与先锋种重要值对于群落性状的影响较小。各功能群与先锋种重要值对于群落地上生物量、盖度、高度和非播种物种数的变异较其种子播种量具有更好的解释力度。这是因为各功能群和先锋种植物可通过自生性状直接影响群落性状,而种子最终不一定能发育成为植株。但是在人工草地建植初期,各功能群重要值与其种子播种量高度相关,各功能群种子播种量可通过间接作用影响人工草地群落。因此,控制各功能群种子播种量可能是调控包括地上生物量在内的人工草地植物群落性状的有效手段。
与前人研究相比,本试验并未发现豆科植物对人工草地生产力的促进作用[3,26],这可能是由于所选用的豆科植物不同以及试验地点海拔过高等原因导致的差异。考虑到豆科植物对牧草品质的提升作用和对土壤的改善作用[2-3],豆科植物依然可以作为高寒人工草地建植的候选物种,但是对于适应高寒地区种植的豆科种质资源需加强筛选和挖掘。本试验发现,先锋种在高寒人工草地建植初期的作用不明显。先锋种可能仅仅在裸地而不是混播草地中才能发挥出较大的作用,同时无纺布也可能不利于先锋种的生长和发育,先锋种植物在人工草地建植中的作用尚需进行更多的研究。
4 结论
禾本科与杂类草植物在高寒人工草地建植初期的作用较大,其中禾本科植物可以形成较高的群落盖度、高度和地上生物量,同时显著减少非播种物种的出现。杂类草的主要影响是降低群落高度和地上生物量,但可显著增加非播种物种数。豆科和先锋种植物在高寒人工草地建植初期的作用相对较弱,对群落性状的影响有限。在建设以生产为主要目的的高寒人工草地时,应优先考虑禾本科植物。主要考虑草地多功能性时,可选择禾本科植物和杂类草植物组合。
猜你喜欢
河南农业·综合版(2021年2期)2021-03-24 11:27:38
河南农业(2021年2期)2021-03-14 10:06:34
湖北农业科学(2020年22期)2020-12-18 07:32:08
黑龙江工程学院学报(2020年5期)2020-10-21 05:37:10
水土保持研究(2019年6期)2019-10-19 03:33:32
河南农业(2018年12期)2018-12-05 08:06:22
湖南城市学院学报(自然科学版)(2016年4期)2016-02-27 14:02:41
农产品市场周刊(2015年39期)2015-10-31 19:12:27
防护林科技(2015年5期)2015-06-10 11:42:52
海南热带海洋学院学报(2015年5期)2015-03-14 09:51:07
