
放牧强度对荒漠草原一年生植物刺穗藜种群特征及空间分布的影响
2021-08-03 13:18郑佳华赵天启乔荠瑢赵萌莉张心玥吴建新
草地学报 2021年7期
张 峰, 郑佳华, 赵天启, 乔荠瑢, 赵萌莉*, 张心玥, 吴建新
(1.内蒙古农业大学草原与资源环境学院, 内蒙古 呼和浩特 010019; 2. 内蒙古农业大学草地资源教育部重点实验室, 内蒙古 呼和浩特 010019; 3.中建二局第一工程有限公司成都分公司, 四川 成都 610000; 4. 内蒙古自治区退耕还林和外援项目管理中心, 内蒙古 呼和浩特 010020)
荒漠草原是我国北方天然的生态屏障,其降水少、草层低矮且其生境脆弱[1],主要利用方式为放牧[2]。近年来,由于过度放牧造成草地不同程度的退化,草地群落结构发生变化,群落盖度、多样性及生产力降低[3],植被空间格局也随之发生变化[4-5]。种群空间分布指其在空间上的聚集或梯度分布,受到多个生态学过程综合作用[6-7],是形成空间格局的主要因素[8]。但以往的研究中主要通过幂函数、随机森林模型等常规方法进行分析[9-10],且主要集中在对土壤水分[11-12]、土壤有机碳[13-14]、土壤碳氮比[15]及土壤理化性质[16]等进行分析,借助地统计学分析自然生态系统的过程及格局和基于种群多度分析的研究相对较少[17-19],尤其是对一年生植物种群空间分布的研究较少。
一年生植物生长周期短,是典型的机会主义者,其种子萌发、生长及结实过程中高度依赖水分。虽然一年生植物生活周期短、植株矮小[20]、结构简单,但其在生态系统中作用不小,尤其是在荒漠植物群落中,具有较大的生物量和生产力[21-22]。而目前关于一年生植物或短命植物的研究主要集中在古尔班通古特沙漠,主要针对短命植物的群落特征[23]、叶片再吸收[24]、开花节律[25]、繁殖特性[26]及化学计量[27]等上,对其空间分布特征的研究较少。一年生植物刺穗藜(Chenopodiumaristatum),为荒漠草原常见种,在降水集中的6—8月份之间即可完成生长、开花、结果,在生长季高峰期时,其生物量较高,对群落生产力贡献较大。以往对刺穗藜的研究较少,本文以荒漠草原一年生植物刺穗藜种群为研究对象,结合常规分析和地统计学分析刺穗藜种群特征及其空间分布特征,探讨其种群特征及空间分布规律对不同载畜率的响应,为草地合理利用提供理论基础。
1 材料与方法
1.1 研究区概况
本试验区位于内蒙古农牧业科学院综合试验示范中心四子王基地(41°46′44″ N,111°53′42″ E),海拔约1 456 m;典型的中温带大陆性季风气候,年均温3.4℃,年均降水量为250 mm左右,降水集中在5—9月。该地建群种为短花针茅(Stipabreviflora),主要优势种为无芒隐子草(Cleistogenessongorica),冷蒿(Artemisiafrigida)和一、二年生植物有刺穗藜(Chenopodiumaristatum)等,植被稀疏低矮,群落平均高度约为8~10 cm,盖度低于20%,群落生产力约为110~120 g·m-2;土壤较贫瘠,主要为淡栗钙土,有机质含量较低,具有缺氮、少磷、钾有余的特点[17]。
1.2 试验设计及样品分析
试验区于2004年进行围建,设置围封对照区(Control,CK)、轻度放牧(Light grazing,LG)、中度放牧(Moderate grazing,MG)和重度放牧(Heavy grazing,HG) 4个放牧强度,3次重复,共12个小区,各小区面积均为4.4 hm2。放牧家畜为当地成年蒙古羯绵羊,载畜率分别为0(CK),0.15(LG),0.30(MG)和0.45(HG)羊单位·ha-1·月-1;自2004年起,放牧期于6月初开始并于11月底结束。
本试验于2020年8月在各小区均设置1个25 m×100 m的大样方,将其均等划分为50个5 m×10 m的中样方(东西宽5 m,南北长10 m),在各中样方西南角放置1个1 m×1 m的样方,以西南角作为起点(0,0),自西向东为X轴,自南向北为Y轴,最西南角坐标为(0,0),最东北角坐标为(20,90),记录各小样方中所有物种的密度;此外,在各小区分别采集100株刺穗藜植株用于单株重、植株高度、叶面积(Leaf area,LA)、比叶面积(Specific leaf area,SLA)、叶片/茎/根系干物质含量(Dry matter content,DMC)、叶片全碳(Leaf total carbon,LTC)、全氮(Leaf total nitrogen,LTN)含量等的测定与分析。测定方法如下:
(1)单株重(Biomass per individual):选取20株刺穗藜去跟后于65℃烘箱中48 h至恒重,使用万分之一天平称重,取其平均值为其单株重。
(2)比叶面积(SLA)=叶面积(LA)/叶干重(leaf dry weight,LDW)。各小区选取10株鲜活刺穗藜个体,对每株刺穗藜个体的所有叶片使用便捷式叶面积仪(YMJ-C)进行扫描测定叶面积,并将每株叶片烘干后使用万分之一天平称重为其叶干重。
(3)叶片/茎/根系干物质含量(DMC)=叶片/茎/根系的干重/鲜重,选择10株鲜活刺穗藜个体进行叶片、茎和根系分离后立即使用万分之一天平称量其鲜重,之后烘干称量其干重。
(4) 叶片全碳(LTC)、全氮(LTN)含量:通过凯氏定氮法使用元素分析仪(Elementar vario MACRO CUBE)进行样品碳、氮测定。
1.3 数据处理
对刺穗藜种群密度、单株重、植株高度、叶长、叶宽、叶面积、比叶面积,叶片/茎/根系干物质含量,叶片全碳、全氮含量及刺穗藜种群相对密度、变异性进行处理间单因素方差分析(One-way ANOVA)并进行Duncan检验,显著性水平为0.05。此外,在SPSS 19.0中进行正态化转换符合正态分布后使用地统计学软件GS+9.0通过克里格(Kriging)插值法绘制刺穗藜2D平面图及3D立体图,通过平面图空间分布及立体图峰谷之间的过渡及凹凸情况来判断刺穗藜种群分布情况。方差分析由SPSS 19.0处理,刺穗藜种群特征图表由SigmaPlot 14.0制作,2D及3D图由GS+9.0制作。变异系数(Coefficient of variation,CV)公式如下:
CV=STDi/Meani
式中:Meani为物种i的密度均值,STDi为物种i的密度标准差。
2 结果与分析
2.1 刺穗藜种群特征
由图1可知,与CK相比,各放牧处理下种群密度均显著增加(P<0.05),但其比叶面积无显著变化。此外,刺穗藜叶片及根系干物质含量无显著变化,叶片全碳、全氮含量也无显著变化。
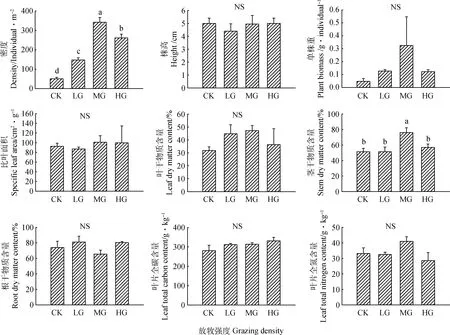
图1 刺穗藜种群特征
2.2 种群密度的空间变异性
由表1可知,与CK相比,放牧显著增加刺穗藜种群相对密度(P<0.05);此外,放牧降低其种群变异性(P<0.05)。
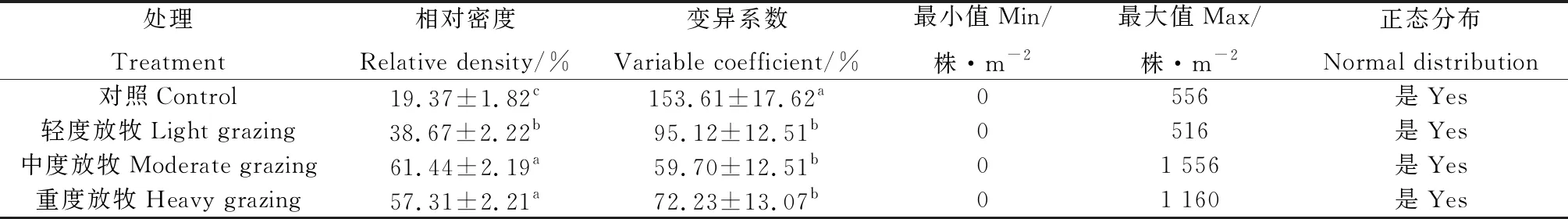
表1 刺穗藜种群密度的描述性统计
2.3 空间格局的2D平面图
如图2所示,CK为梯度分布,刺穗藜种群主要集中在右下方(东南角),以此为中心向左上方(西北角)梯度扩散;而在LG,MG和HG中,刺穗藜种群主要为斑块分布,存在多个小斑块,但刺穗藜种群整体分布较均匀。

图2 不同放牧强度下刺穗藜种群空间分布2D平面图
2.4 空间格局的3D立体图
从图3的起伏剧烈程度及凹凸情况并结合图2可知,CK中虽无剧烈起伏,但呈现出由高到低的梯度分布;而LG,MG和HG中刺穗藜种群虽存在起伏及凹凸现象,但整体起伏程度较小,变化平缓,整体差异较小,分布较均匀,空间异质性较小。

图3 不同放牧强度下刺穗藜种群空间分布3D立体图
3 讨论
本研究中,各放牧处理均使一年生植物刺穗藜种群密度增加。这可能是由于长期围封下的草地群落缺乏干扰,使得枯落物累积而阻碍植株繁殖更新[28-29],放牧处理下枯落物较少,当降水来临时一年生植物能迅速萌发生长,使其种群密度增加。其次,放牧会对植物株丛大小、分蘖数量、种群密度及种间关系等均有调节作用[30-31]。一年生植物生活周期短、植株矮小[20],家畜在采食过程中,主要以植株较高且占据优势的物种如短花针茅、无芒隐子草等为食,造成其株丛出现破碎化[32]和矮型化[33-34],从而释放较多空间与资源,使得一年生植物可以利用的空间和养分资源增加。
本研究表明,放牧造成刺穗藜种群密度变异性降低,其空间异质性也降低。在对种群异质性的研究中,吕世杰等[35]研究表明放牧造成无芒隐子草种群变异性降低;张峰等[17]对糙隐子草种群变异性的研究也如此。本研究表明围封中刺穗藜种群呈现出梯度分布,其空间异质性较大;而放牧使其呈现出斑块分布并使其异质性降低。也有研究表明[36]放牧增加了糙隐子草种群空间异质性,这主要是由于各研究立地条件、放牧强度不一致及家畜选择性采食及践踏作用,造成各种群空间分布格局不一致。本研究中一年生植物刺穗藜主要是依靠种子繁殖,由于种子成熟主要围绕母株扩散,造成其呈现出梯度分布;而在放牧干扰下家畜的选择性采食、践踏及粪尿归还等作用,使其从梯度分布变为斑块分布,由于其起伏变化及波峰波谷的凹凸程度较小,种群空间分布较均匀,故放牧下异质性较小。
4 结论
与围封相比,放牧增加了刺穗藜种群密度,但并未改变其性状特征;放牧降低了刺穗藜种群变异性及其空间异质性,使其分布更均匀。
猜你喜欢
数学物理学报(2022年1期)2022-03-16
数学物理学报(2021年6期)2021-12-21
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
应用数学(2020年2期)2020-06-24
新疆农业科学(2020年1期)2020-02-14
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
体育科研(2016年5期)2016-07-31
中国麻业科学(2015年5期)2015-12-28
中医研究(2014年8期)2014-03-11
中医研究(2014年8期)2014-03-11
