
辽东山区落叶松-水曲柳混交林及其纯林生长与生物量分配特征1)
2021-07-30 08:06冯健高慧淋王骞春
东北林业大学学报 2021年7期
冯健 高慧淋 王骞春
(辽宁省林业科学研究院,沈阳,110032) (沈阳农业大学) (辽宁省林业科学研究院)
曹颖 卜鹏图 王占伟 陈东升
(辽宁省林业发展服务中心) (辽宁省林业科学研究院) (中国林业科学研究院林业研究所)
随着我国大力开展植树造林工作,我国林业建设取得了长足进展,森林面积已达2.08亿hm2,其中人工林保存面积6 933万hm2,规模居世界首位[1]。但目前我国的人工林大部分为单一树种的纯林,许多研究认为单一树种的纯林由于结构简单从而存在着养分周转效率低、林分稳定性差、生产力低、抵御自然灾害和病害能力弱等问题。从现在国内外的研究结果来看,营造混交林(同龄和异龄)是改善林分结构、增加生物多样性、提高林分生产力和增强林分稳定性的重要手段[2-4]。落叶松(Larixspp.)是我国重要的造林和用材树种,据我国第九次森林资源清查数据显示,现有落叶松人工林316.29万hm2,蓄积达2.37亿m3[5]。其林分质量的提升对于我国人工林建设有重要意义。目前,我国营造的落叶松人工林也多为纯林,Wang等[6]研究得出落叶松纯林在中、近熟林阶段存在地力退化的问题,陈立新[7]、刘世荣[8]的研究也证实落叶松纯林存在地力退化、土壤酸化等问题,为了提高落叶松纯林的生态稳定性和林分土壤环境,一些学者开展了落叶松与其他树种混交的研究,这些研究多集中在林分生长和生物多样性等方面,如落叶松与白桦、云杉和水曲柳(FraxinusmandshuricaRupr.)混交的研究,边丽宁比较了华北落叶松(Larixprincipis-rupprechtii)—白桦(BetulaplatyphyllaSuk.)、落叶松—云杉(PiceaasperataMast.)混交林和纯林在生长和多样性指标间的差异,均表现为混交林优于纯林[9]。另外,也有学者关注混交林生物量方面的情况,如夏成财等[10]研究16年生落叶松-白桦混交林认为混交林中乔木层生物量显著高于落叶松纯林和白桦纯林。从上述研究结果来看,与纯林相比,落叶松纯林通过混交合适的树种确实改善了养分周转效率、提高了生物多样性和林分生产力。辽东山区是辽宁省森林的主要分布区,分布着大面积的落叶松单一纯林,是否可以通过混交的方式改善其生长和生态稳定性是值得关注的问题。有关落叶松在生长和生物量方面的研究多选择纯林作为研究对象,郭书彬等[11]关于经营密度对华北落叶松人工林生长的影响研究,陈东升等[12]关于不同年龄日本落叶松人工林生物量、碳储量及养分特征的研究。有关水曲柳在生长方面的研究多集中于纯林,刘可欣[13]研究了修枝对水曲柳人工林生长和生理特性的影响,生物量方面的研究多集中于根生物量的研究,谷加存等[14]研究了造林密度对水曲柳人工林地上生长和细根生物量的影响。有关落叶松与水曲柳混交林生长及生物量在该地区的研究却鲜有报道。因此,本研究以落叶松-水曲柳混交林为研究对象,分析混交林及其对应纯林的生长特性和生物量分配比例,并探讨生长量、生物量与植被多样性、土壤养分的关系,为科学评估这两个树种混交后的林地生产力提供理论依据。
1 研究区概况
研究地位于辽宁省新宾满族自治县中南部的陡岭林场。处于长白山系龙岗山脉延伸部分,境内平均海拔约500 m、山势较缓,坡度在10°~25°,为低山丘陵区。属北温带季节性大陆气候,四季分明,气候宜人,雨量充沛,年平均降水量在800 mm。主要乔木树种有落叶松、樟子松(Pinussylvestrisvar.mongolicaLitv.)、黑松(PinusthunbergiiParl)、红松(PinuskoraiensisSieb. et Zucc.)、蒙古栎(QuercusmongolicaFisch. ex Ledeb.)、核桃楸(JuglansmandshuricaMaxim.)、水曲柳等;林下灌木主要有卫矛(Euonymusalatus(Thunb.) Sieb)、忍冬(LonicerajaponicaThunb.)、榛子(CorylusL.)、辽东楤木(Araliaelata(Miq.) Seem.)、刺五加(Acanthopanaxsenticosus(Rupr. et Maxim.) Harms.)等[15]。
2 研究方法
2.1 样地选取
2019年6—8月,在坡向、坡位等基本一致的立地条件下选取落叶松-水曲柳人工混交林、落叶松人工纯林、水曲柳人工纯林3个林型,其中落叶松-水曲柳混交林为行状混交(5行×3行)作为研究对象。采用典型样地法,在每个林型中分别设置3块20 m×30 m的标准地,共9个样地。样地具体情况见表1。
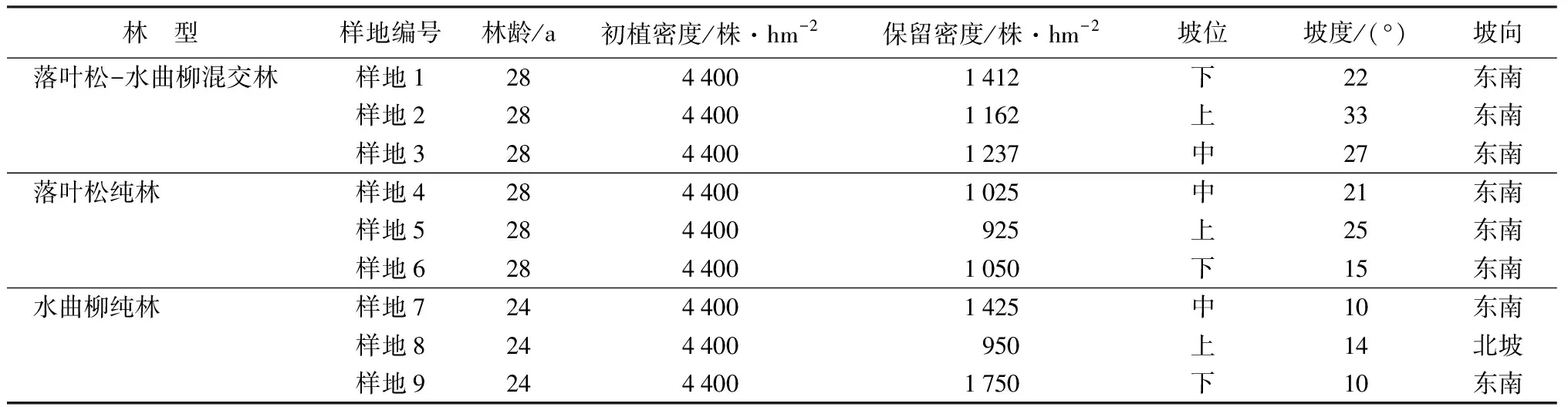
表1 样地基本概况
2.2 生长指标测定
对样地内的乔木进行每木检尺,测量其树高、活枝下高、胸径和冠幅等。树木高度、活枝下高采用超声波测高仪进行测量,精度0.1 m。胸径采用测树围尺测量,测量精度0.1 cm。落叶松和水曲柳的材积分别用下述公式计算:
落叶松材积:
V=0.000 051 269 179D1.816 998 7H1.103 414 6,
水曲柳材积采用平均实验形数法计算:
V=(H+3)·g1.3·f3。
式中:H为树高;g1.3为胸径的横截面积;f3为水曲柳平均形数为0.40[16]。
冠高率即冠高与树高的比值;高径比即树高与胸径的比值;冠径比即树冠与胸径的比值。
2.3 生物量指标测定
依据固定样地的每木检尺数据,采用等断面积标准木法将样地内所有树木分为五级,在第一、三、五级分别选取优势木、平均木、劣势木各1株。对于落叶松和水曲柳纯林,分别选取落叶松及水曲柳解析木各3株;对于混交林,在两个树种的交界处选取落叶松及水曲柳各3株,分别在两个树种的内部选取落叶松及水曲柳各3株。为此,共获取解析木18株,其中落叶松9株,水曲柳9株。
2.4 数据处理与分析
采用SPSS22.0软件中的比较平均值进行单因素方差、多重比较(LSD法)及相关性分析,F检验结果若P<0.05为差异显著,若P<0.01为差异极显著。
3 结果与分析
3.1 落叶松-水曲柳混交林生长特性
3.1.1 落叶松-水曲柳混交林单株树木生长特性
落叶松-水曲柳混交林与落叶松、水曲柳纯林胸径、树高、材积、冠幅和冠高比较表明(表2),胸径、单株材积、冠高和冠幅表现为纯林大于混交林,其中,落叶松冠高和冠幅差异达到显著水平(P<0.05),水曲柳冠幅差异达到极显著水平(P<0.01),其他指标差异均不显著(P>0.05);树高和活枝下高表现为混交林大于纯林,但差异均不显著(P>0.05),其中,混交林中落叶松、水曲柳树高分别比落叶松、水曲柳纯林树高大2.06%和2.79%,混交林中落叶松、水曲柳活枝下高占树高比例分别比落叶松、水曲柳纯林提高了5.55%和2.01%。

表2 落叶松、水曲柳单木生物量统计 kg·株-1
落叶松和水曲柳间单株树木生长特性比较表明,胸径、树高、活枝下高和冠高均表现为落叶松大于水曲柳,其中,胸径和冠高差异达到极显著水平(P<0.01),树高差异达到显著水平(P<0.05),活枝下高差异不显著,混交林中落叶松胸径、树高、活枝下高和冠幅分别比水曲柳大3.48 cm、3.02 m、1.49 m和1.44 m;冠幅表现为水曲柳大于落叶松。
3.1.2 落叶松-水曲柳混交林单株树木形质特性
落叶松-水曲柳混交林中落叶松与落叶松纯林比较表明(表3),冠高率和冠径比表现为纯林大于混交林,差值均为0.05,其中,冠高率差异达到显著水平(P<0.05);高径比表现为混交林大于纯林,差值为0.09,但差异不显著(P>0.05)。落叶松-水曲柳混交林中水曲柳与水曲柳纯林比较表明(表3),冠高率和冠径比表现为纯林大于混交林,差值分别为0.02和0.09,其中,冠径比差异达到显著水平(P<0.05);高径比表现为混交林大于纯林,差值为0.17,差异达到极显著水平(P<0.01)。落叶松和水曲柳间树木形质比较表明,冠高率和冠径比表现为落叶松大于水曲柳;高径比表现为水曲柳大于落叶松,且差异达到极显著水平。
总体来看,混交促进了落叶松和水曲柳的高生长;促进了自然整枝,增加了活枝下高长度;使林木更加圆满通直;与落叶松纯林相比,增加了林分蓄积。落叶松树高和活枝下高均明显高于水曲柳,落叶松-水曲柳混交林复层林结构明显。
3.2 落叶松-水曲柳混交林及对应纯林生物量分配分析
对落叶松、水曲柳在混交林和纯林中各器官生物量所占比例进行分析表明(表3,4),落叶松、水曲柳m(地上生物量)∶m(地下生物量)、m(树干生物量)∶m(总生物量)表现为混交林大于纯林;m(叶生物量)∶m(总生物量)、m(根生物量)∶m(总生物量)表现为纯林大于混交林;落叶松m(枝生物量)∶m(总生物量)表现为纯林大于混交林,而水曲柳m(枝生物量)∶m(总生物量)表现为混交林大于纯林。不同林型、不同树种各器官生物量占总生物量的比例由大到小均为树干生物量、根生物量、枝生物量、叶生物量;其中,树干生物量占比较高,达到52.86%~62.37%;叶生物量占比最低,在1.10%~2.37%。

表3 落叶松-水曲柳混交林及其纯林生长特性
3.3 落叶松-水曲柳混交林及其纯林生长量与生物量相关分析
如表5所示,落叶松-水曲柳混交林及其纯林生长量与生物量相关性较强,其中,胸径与m(地上生物量)∶m(地下生物量)、m(树干生物量)∶m(总生物量)呈显著的负相关,与m(根生物量)∶m(总生物量)呈显著的正相关;单株材积与m(叶生物量)∶m(总生物量)呈显著正相关。此外,树高与m(地上生物量)∶m(地下生物量)、m(树干生物量)∶m(总生物量),枝下高与m(叶生物量)∶m(总生物量),冠高与m(地上生物量)∶m(地下生物量)、m(树干生物量)∶m(总生物量)、m(根生物量)∶m(总生物量)的相关性也较强,但未达到显著及以上水平(P>0.05)。
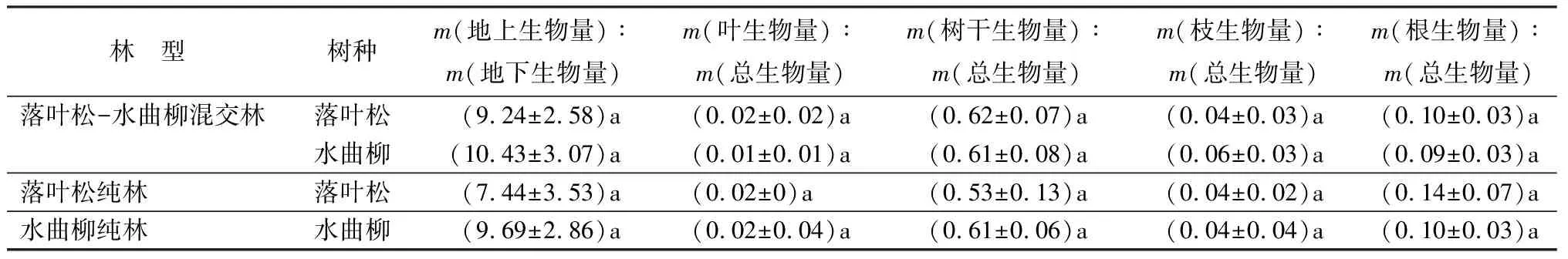
表4 不同林型落叶松、水曲柳不同器官单株生物量占比

表5 各生长量与纯林生长量间相关系数
3.4 落叶松-水曲柳混交林及其纯林生长量、生物量与植被多样性相关分析
如表6所示生长指标与植被多样性指标相关性较弱,其中,单株材积与草本层在丰富度指数(S)上呈显著正相关,林地蓄积与灌木层Simpson多样性指数(D)、群落均匀度指数(JD)和Shannon-Wiener多样性指数(H)呈显著的负相关。另外,树高与灌木层S,冠高与灌木层S、D、JD和JH呈较强的正相关,而蓄积与灌木层S和JH呈较强的负相关。

表6 生长量与植被多样性指数相关系数
如表7所示,生物量指标与植被多样性呈一定的相关性。各器官生物量占总生物量比值与植被多样性相关性较低,只有m(叶生物量)∶m(总生物量)与草本层S指数成显著正相关。另外,m(树干生物量)∶m(总生物量)与草本层H指数呈较强负相关;m(枝生物量)∶m(总生物量)与草本层S指数呈较强负相关;m(根生物量)∶m(总生物量)与草本层S和H指数呈较强正相关;m(地上生物量)∶m(地下生物量)与草本层S和H各指数呈较强的负相关。

表7 生物量与植被多样性指数相关系数
4 讨论
本研究表明,落叶松-水曲柳混交林的树高和活枝下高分别比对应的纯林高,其中,混交林中落叶松树高和活枝下高分别比纯林中落叶松提高了2.06%和5.55%,混交林中水曲柳树高和活枝下高分别比纯林中水曲柳提高了2.79%和2.01%。夏成财等研究落叶松—白桦混交林生长特性得出结论,混交林中落叶松、白桦的平均树高比各自纯林分别高出15.1%和20.5%,且差异显著[10]。盛伟彤等[17]研究福建柏(Fokieniahodginsii(Dunn) Henry et Thomas)和杉木(Cunninghamialanceolata(Lamb.) Hook.)混交林也得出结论,混交后杉木生长得到促进,树高比纯林提高5.5%,分析原因在于混交林中杉木占据林冠上层,福建柏位于林冠下层,给杉木提供侧方遮荫。另外,其研究还认为混交林促进了自然整枝[18-20]。这些结论均与本研究结果相一致,表明通过落叶松与水曲柳混交,可以促进落叶松、水曲柳树高生长,提高自然整枝能力。本研究中混交林虽然有促进树木高生长和自然整枝的表现,但这种表现在混交林和纯林中的差异没有达到显著水平,分析可能的原因在于林龄,有研究表明,混交对树高生长方面的作用会因林龄的不同而有所不同[21],本研究中的混交林处于中龄林到近熟林的过度阶段,正是混交开始对树高生长起作用的阶段,因此,本研究中混交林与纯林在树高表现为混交林大于纯林,而其差异却不显著,后续应持续观测,为最终结论提供数据支撑。本研究还表明,落叶松和水曲柳在单株树木生长特性方面的比较表现为胸径、树高、活枝下高和冠高均为落叶松大于水曲柳,形成了落叶松-水曲柳混交复层林结构。这一结构有利于林分生长,邓海燕等认为混交林形成明显的桉树(EucalyptusrobustaSmith)在上层而混交树种在下层的复层林分结构,这可使各树种充分利用林地空间环境资源,减少种间竞争,提高林分生长量,故团花(Neolamarckiacadamba(Roxb.) Bosser)、马占相思(AcaciamangiumWilld.)或湿加松(Pinuselliottii)混交均能促进桉树树高、胸径、材积生长。复层林的形成不仅可增强林木的光能利用率,提高生产力,还可调节林下气候环境,为适应不同生存空间的动物提供更多层次的场所,提高生态系统的物种多样性[22-23]。
本研究中混交林与纯林比较表明,通过落叶松与水曲柳的混交,落叶松提高了地上生物量和树干生物量占总生物量的比例,m(地上生物量)∶m(地下生物量)由纯林的7.44提高到混交林的9.24,m(树干生物量)∶m(总生物量)由纯林的0.53提高到混交林的0.62,水曲柳提高了地上生物量和枝生物量占总生物量的比例,m(地上生物量)∶m(地下生物量)由纯林的9.69提高到混交林的10.43,m(枝生物量)∶m(总生物量)由纯林的0.04提高到混交林的0.06。这表明通过营造混交林,改变了落叶松和水曲柳各器官生物量的分配。这一结果与其他学者研究结果一致[24]。覃林等[25]研究南亚热带红椎(Castanopsishvstrix)—马尾松(PinusmassonianaLamb.)纯林及其混交林生物量分配格局认为,林分乔木层生物量各组分的空间分配格局因林型树种组成不同而存在差异,红椎—马尾松混交林、红椎纯林和马尾松纯林乔木层生物量的空间分配分别为:树干材(56.01%)>枝(18.39%)>根(14.48%)>干皮(5.57%)>叶(5.55%)、干材(62.25%)>根(17.16%)>枝(11.76%)>干皮(6.84%)>叶(1.99%)和干材(49.31%)>枝(19.98%)>根(17.72%)>叶(8.80%)>干皮(4.19%)。本研究表明,不同林型、不同树种各器官生物量占总生物量的比由大到小为树干生物量、根生物量、枝生物量、叶生物量,其中,树干生物量占比较高,达到52.86%~62.37%;叶生物量占比最低,在1.10%~2.37%。这一结论与其他学者关于落叶松生物量分配格局的研究相一致[12,26-27]。陈东升等[21]研究落叶松纯林认为,从幼龄林到成熟林,落叶松树干生物量占比在39.7%~66.2%,而叶生物量占比在1.7%~11.8%。本研究表明,通过混交,落叶松和水曲柳树干生物量所占比例均有所提高,与本研究中生长量的结果相一致,即混交林提高了落叶松和水曲柳树高的生长,树高生长量提高了,自然就提高了树干的生物量,从而提高了树干生物量占总生物量的比例,这对于培育用材林是有利的。本研究中落叶松和水曲柳地上生物量与地下生物量比例均表现为混交林较纯林有所提高,这一点应引起重视,应采取合理的经营措施进行人工干预和调整,如果地上部分比例过大,则可能会造成根茎比下降,对林分后期的生长发育构成障碍,对获得林分后期较高的生长速度不利[28]。本研究中混交林中落叶松、水曲柳叶生物量占比均比纯林低,其原因可能是因为纯林中树木个体均处于同一林层,为了得到更多的光照,会通过增加叶面积提高光利用率,而混交林中落叶松和水曲柳树高存在差异,形成了上下不同的林层,因而光利用效率更高,根据最优分配理论反推,落叶松和水曲柳不需要把更多生物量分配到叶中,因而叶生物量所占比例降低[29-31]。上述研究结果说明,在相同的立地条件下,由于造林模式的不同,使落叶松、水曲柳各器官生物量分配比例发生了明显的改变。
本研究结果表明,胸径、树高和冠高与落叶松、水曲柳各器官生物量分配格局的相关性较强,其中,胸径与m(地上生物量)∶m(地下生物量)、m(树干生物量)∶m(总生物量)呈显著的负相关,与m(根生物量)∶m(总生物量)呈显著的正相关。何怀江等[32]研究针阔混交林12个树种生物量分配规律认为,叶和树干生物量分配与胸径呈显著的负相关关系,即随着胸径的增大,叶和树干的生物量降低。这一结论与本研究结果相一致。生长指标与灌木层植被多样性关系较紧密,而生物量分配与草本层关系较紧密,其中,林地蓄积与灌木层D、JD和H各指数达到显著负相关,m(叶生物量)∶m(总生物量)与草本层S指数达到显著正相关。这一结果表明,通过落叶松与水曲柳的混交,促进了林地草本和灌木的物种多样性,草本和灌木的物种多样性的增加改变了林地环境,加速了林地养分循环,进而促进了落叶松和水曲柳的生长,也改变了各器官生物量分配比例。
5 结论
落叶松和水曲柳混交促进了落叶松、水曲柳树高生长,分别较对应纯林提高2.06%和2.79%;增加了活枝下高,分别比对应纯林提高5.55%和2.01%;乔木层形成了落叶松-水曲柳混交复层林结构,增强林木的光能利用率,提高了生产力。
落叶松和水曲柳混交提高了地上生物量、树干生物量和枝生物量占总生物量的比例,其中,落叶松m(地上生物量)∶m(地下生物量)提高了1.8,m(树干生物量)∶m(总生物量)提高了0.09,水曲柳m(地上生物量)∶m(地下生物量)提高了0.74,m(枝生物量)∶m(总生物量)提高了0.02。
落叶松-水曲柳混交林在一定程度上促进了树高生长、加速了自然整枝,提高了树木圆满度,复层林结构明显。落叶松-水曲柳混交林在林分生长和生物量分配比例上优于纯林,这种针阔混交模式可用于辽东山区培育用材林。
猜你喜欢
林业科技情报(2021年1期)2021-04-14
今日中国·西班牙文版(2021年4期)2021-04-09
农民致富之友(2019年11期)2019-05-23
森林工程(2018年4期)2018-08-04
现代园艺(2018年2期)2018-03-15
方圆(2018年23期)2018-01-07
农民致富之友(2017年23期)2018-01-02
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
