
盐胁迫对匈牙利能源植物SZARVASI-1种子萌发和幼苗生长的影响
2021-07-26 02:06曾继娟朱强乔改霞
江苏农业科学 2021年11期
曾继娟 朱强 乔改霞

摘要:以银川森淼现代林业科技园2017年采收的匈牙利能源植物SZARVASI-1种子为材料,采用控制试验,通过对匈牙利能源植物SZARVASI-1种子及幼苗进行不同浓度的NaCl溶液处理,研究其萌发期的发芽率、幼苗期株高、根长、地上生物量、地下生物量、总生物量以及根冠比的变化,旨在丰富耐盐牧草种质资源,为耐盐植物新品种选育与开发利用提供基础。结果表明,随着NaCl胁迫的加剧,匈牙利能源植物SZARVASI-1的发芽率、生物量等指标呈先增加后减少的趋势,适宜其种子萌发的最适NaCl浓度为0.8%;适宜其幼苗生长的最佳NaCl浓度为0.6%,超过此浓度则产生抑制作用,植物通过增加地下部分的生长来适应逆境。
关键词:能源植物;盐胁迫;萌发;生物量
中图分类号:S184 文献标志码: A
文章编号:1002-1302(2021)11-0186-05
收稿日期:2020-10-12
基金项目:农业部引进国际先进农业科学技术项目(编号:2016-X14)。
作者简介:曾继娟(1990—),女,甘肃白银人,硕士,助理研究员,主要从事植物资源收集与抗逆生理研究。E-mail:981315800@qq.com。
通信作者:朱 强,硕士,副研究员,主要从事植物资源收集与开发利用研究。E-mail:qzhu2008@163.com。
化石能源逐渐短缺,环境污染日益加重,能源植物的开发利用成为热点。第一代能源植物如玉米(Zea mays)、小麦(Triticum aestivum)、大豆(Glycine max)等基于其大多为一年生的特性,每年需要耕地播种,投入高、产出低[1],已经很难满足发展需求。因此,多年生草本、速生树种等第二代能源植物的开发已成为当前的研究重点[2]。盐分是限制植被产量和品质最重要的环境因素之一[3],对种子萌发、幼苗生长及光合作用的影响较为显著。目前,盐碱地是人类面临的共同问题,主要集中分布在内陆的干旱和半干旱地区[4],宁夏回族自治区干旱、半干旱、大面积的盐碱地是制约当地农业和畜牧业发展的主要瓶颈。如何有效利用和改善盐碱地,已经引起人们的高度关注。近年来,人们在生物措施治理盐碱地方面开展了大量工作,有关种子萌发及生长发育的耐盐性研究多集中于豆科牧草[5]、草坪草[6]、禾本科牧草[7]和经济作物[8]。
牧草在改良和利用盐碱地方面起到重要作用,禾本科牧草的耐盐性通常优于豆科牧草和作物[9]。牧草种子能否在盐胁迫条件下正常萌发,是保证植物生长发育的前提[10],因此研究盐分胁迫对牧草种子的萌发及幼苗生长意义重大。本研究通过开展匈牙利能源植物SZARVASI-1在盐胁迫条件下的萌发和苗期生长情况分析,明确其耐盐能力,旨在丰富耐盐牧草种质资源,为宁夏乃至西北地区能源植物草地的建立、耐盐新品种选育开发和盐碱地改良利用提供理论依据。
1 材料与方法
1.1 物种概况
SZARVASI-1能源草是匈牙利科学家用了15年时间成功培育出的新型草种,2004年获得匈牙利专利,2005年获得欧盟25个国家的专利,其各项性能指标处于国际领先地位。SZARVASI-1能源草是一种生态适应性强、生物产量高、产草期长、用途广泛、耐盐碱的优良生物质能源植物,可用来做固体燃料、造纸和工业纤维原料、饲料等。目前国内河北省沧州市于2013年开展了SZARVASI-1能源草的引种栽培相关研究,试验结果表明,SZARVASI-1能源草在抗盐碱、作为牛羊饲料方面有较好的应用前景。
1.2 试验材料
2015年9月18日,宁夏林业研究院种苗生物工程国家重点实验室通过国家外专局引进匈牙利能源草SZARVASI-1种子10 kg,放于4 ℃冰箱保存,于2016年4月在银川森淼现代林业科技园进行田间播种。
1.3 试验方法
1.3.1 种子胁迫处理 以2017年在森淼现代林业科技园采收的种子为试材,挑选饱满且无病虫害的种子用0.1% KMnO4消毒15 min,再用蒸馏水反复冲洗干净后置于垫有2层滤纸的无菌培养皿中,培养皿规格为直径15 cm,每皿50粒。設置NaCl质量分数分别为0、0.2%、0.4%、0.6%、0.8%、1.0%、1.2%,共7个处理,每个处理5次重复。每皿加入200 mL,分2次进行胁迫,对照用蒸馏水,24 h后观察并记录发芽情况,萌发以胚根长达到种子长度50%为标准。以后每隔24 h浇水观察1次,于17:00—18:00用称质量法确定每皿的失水量,并用等量蒸馏水补充水分。
1.3.2 幼苗胁迫处理 于2018年3月16日在宁夏银川森淼现代林业科技园(38°25′06″N,106°10′35″E)温室内开展试验。将沙土过筛,按照体积比沙土 ∶蛭石=1 ∶1 的比例混合均匀,装入无孔花盆(口径23 cm、底径18 cm、高22 cm)中,每盆装土2.0 kg,盆内土壤pH值为8.5,有机质含量24.6 g/kg,全氮含量1.36 g/kg,全磷含量1.07 g/kg,全钾含量18.1 g/kg。将种子均匀适量的播撒于盆内,每个花盆中施入5 g复合肥,浇水至蛭石完全润湿,在温室内培养。出苗后,待长出3~4张真叶时定苗35株,待植物具有4~5张真叶时进行NaCl胁迫处理。
采用完全区组试验设计,于2018年4月5日进行盐分胁迫处理,设置NaCl质量分数分别为0、0.2%、0.4%、0.6%、0.8%、1.0%、1.2%,共7个处理,每个处理5次重复。于16:00—18:00进行胁迫,每盆800 mL,分数次浇透花盆,对照浇灌蒸馏水,为避免盐分在土层积累,从次日起每天17:00—18:00用称质量法确定每盆的失水量,并用等量蒸馏水补充失水;同时,为防止盐分渗漏,在花盆底部加垫,定期将漏液倒入盆内。
1.3 指标测定及方法
发芽率、发芽势、发芽指数与发芽时间指标测定参照以下公式:
发芽率=(第14天所有发芽的种子数/供试种子数)×100%;发芽势=n/N×100%(式中:n为日发芽最高峰的种子数,N为种子总数);发芽时间(t):指发芽开始至发芽结束的时间;发芽指数(GI)=∑(Gt/Dt) (式中:Gt为第t天种子萌芽数,Dt为对应的种子发芽的天数)。
于盐分胁迫30、60 d后,分别采集样品带回室内测定生物量相关指标。对各处理每株幼苗的分蘖数进行统计,分蘖数指地面以下或近地面处所发生的分枝。分别收集茎、叶,用水清洗每株幼苗的地下部分,同时将收集好的茎、叶、根置于烘箱,105 ℃ 处理10 min,然后在65 ℃下烘干至恒质量[11]。计算根冠比(RSR),即:地下干物质与地上干物质生物量的比值[12],其中,株高、根长用直尺测定。
1.4 数据分析
采用SPSS16.0软件对数据进行统计分析,用Duncans新复极差法进行差异显著性多重比较。
2 结果与分析
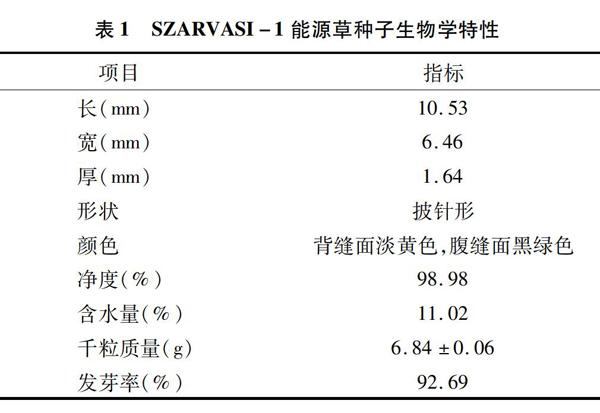
2.1 种子形态特征
SZARVASI-1能源草为禾本科披碱草属植物,果实是典型的颖果,种子生物学特性见表1,种子形态见图1。
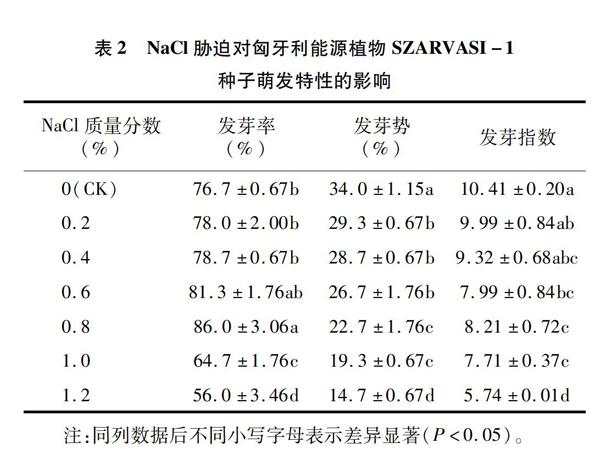
2.2 NaCl胁迫对匈牙利能源植物SZARVASI-1种子萌发特性的影响
匈牙利能源植物SZARVASI-1种子的发芽率随着盐浓度的增加呈先增大后减小的趋势(表2),在NaCl浓度为0.8%时有最大值,发芽率与对照差异显著,且分别较NaCl浓度1.0%、1.2%显著高出32.9%、53.6%;发芽势、发芽指数的变化规律相一致,均随着盐浓度的增加而减小,在CK时有最大值,且发芽势在对照时显著高于其他处理(P<0.05);但是发芽率、发芽指数在CK、0.2%、0.4%处理间差异不显著。表2表明匈牙利能源植物SZARVASI-1种子的萌发受盐胁迫环境的影响非常明显,适当的NaCl溶液有助于其种子萌发,当NaCl浓度超过1.0%时,则抑制其萌发。
2.3 NaCl胁迫对匈牙利能源植物SZARVASI-1苗期株高的影响
NaCl胁迫30 d时,随着盐浓度的增大,匈牙利能源植物SZARVASI-1的株高呈先升高后降低的趋势(图2),在NaCl浓度为0.6%时达到峰值47.4 cm,较对照(CK)显著增加29.51%(P<0.05);随着胁迫时间的延长,即胁迫60 d时,其株高在NaCl浓度为0.6%时有最大值,但是与CK之间没有显著性差异(P>0.05),随着盐浓度的增大,株高呈直线下降趋势,并且在NaCl浓度为0.8%、1.0%、1.2%三者之间均存在显著性差异(P<0.05),说明适宜浓度的NaCl溶液(0.6%)能够促进SZARVASI-1株高生长,浓度过大则产生抑制作用。
2.4 NaCl胁迫对匈牙利能源植物SZARVASI-1苗期根长的影响
胁迫30 d时,随着盐浓度的增大,匈牙利能源植物SZARVASI-1的根长呈先升高后降低的趋势(图3),在NaCl浓度为0.6%时达到峰值13.1 cm,较CK显著增加47.19%(P<0.05),但是与NaCl浓度为0.4%、0.8%时差异不显著(P>0.05);随着胁迫时间的延长, 即胁迫60 d时, 其根长变化规律基本一致,同样在NaCl浓度为0.6%时有最大值,较CK显著增加21.64%(P<0.05),与NaCl为浓度0.4%、0.8%时差异不显著,随着盐浓度的增大,其根长明显降低,说明该植物生长对NaCl胁迫较为敏感。
2.5 NaCl胁迫对匈牙利能源植物SZARVASI-1苗期生物量的影响
胁迫30 d时,随着盐浓度的增大,匈牙利能源植物SZARVASI-1的地上生物量呈先增加后减少的趋势(图4),在NaCl浓度为0.6%时有最大值,较CK显著增加14.47%(P<0.05),除NaCl浓度为0.4%外,与其他处理间均存在显著性差异;随着胁迫时间的延长,其地上生物量的变化规律愈为明显,并且在NaCl浓度1.2%时,较CK顯著减少10.45%,说明NaCl浓度等于或大于1.2%时显著限制了SZARVASI-1地上生物量。
胁迫30 d时,随着盐浓度的增大,匈牙利能源植物SZARVASI-1的地下生物量变化趋势与地上生物量相一致(图5),在NaCl浓度为0.6%时达到峰值2.879 g,较CK显著增加12.02%(P<0.05),且NaCl浓度为1.2%时较CK显著降低9.8%,其他各处理间均不存在显著性差异(P>0.05);随着胁迫时间的延长,其变化规律更为明显,在NaCl浓度为0.6%时较CK显著增加19.85%(P<0.05),但是NaCl浓度为1.2%时也较CK显著增加7.59%(P<0.05),与胁迫30 d呈相反的格局,说明该植物可以通过地下生物量的增加来调节自身对于盐胁迫的耐受性,逐渐适应环境。
胁迫30 d时,随着盐浓度的增大,匈牙利能源植物SZARVASI-1的总生物量呈先增加后减少的变化趋势(图6),在NaCl浓度为0.6%时达到峰值6.87 g,较CK显著增加13.55%(P<0.05),且与其他各处理之间均存在显著性差异(P<0.05),但是CK与处理NaCl浓度为0.2%、0.8%、1.0%之间差异不显著(P>0.05);随着胁迫时间的延长,其总生物量的变化规律更加明显,并且各处理间均存在显著性差异(P<0.05),当NaCl浓度为1.2%时,总生物量较CK显著降低4.5%,表明该浓度下,总生物量的降低主要由地上生物量的减少引起。
2.6 NaCl胁迫对匈牙利能源植物SZARVASI-1苗期根冠比的影响
胁迫30 d时,随着盐浓度的增大,匈牙利能源植物SZARVASI-1的根冠比变化趋势不明显(图7),在NaCl浓度为1.0%时达到峰值,较CK显著增加8.11%(P<0.05),与处理NaCl浓度为0.2%、0.8%之间不存在显著性差异(P>0.05);随着胁迫时间的延长,处理60 d时其根冠比在NaCl浓度为1.2%时有最大值,较CK显著增加20.93%,与其他各处理间均存在显著性差异(P<0.05),说明该浓度下,植物可能通过减少地上生物量或增加地下生物量来适应环境。
3 讨论与结论
盐是影响植物种子萌发和幼苗生长的重要因子之一[13]。罗志娜等通过对燕麦 (Avena sativa) 种子萌发的耐盐性研究发现,燕麦品种的发芽率、芽长和根长均随着盐浓度的增加表现出先升高后降低的趋势,低浓度盐胁迫可以促进种子萌发,高浓度盐胁迫明显抑制种子萌发[14-15]。也有研究指出,在盐生植物中,轻度盐分溶液促进种子萌发,当盐浓度达到一定值时,增加的盐浓度对植物的种子萌发和幼苗生长形成抑制效应[16-17]。本研究结果表明,匈牙利能源植物SZARVASI-1种子的发芽率在NaCl浓度为0.8%时有最大值,超过此浓度则抑制其萌发。史燕山等的研究表明,低盐浓度下,某些植物种子的萌发率显著高于对照,而高盐胁迫下,种子的萌发率显著降低[18-19],本研究结果与之相一致。由此可见,高浓度盐胁迫对种子萌发有抑制作用,抑制作用的大小因材料耐盐性强弱不同而有差异。
植物在不同环境条件下的资源分配格局反映了植物生长对环境的响应规律和资源分配策略。中国西北地区盐渍化蔓延,盐碱地上的盐度成为影响植物生长的重要因子[20],植物对于盐分的适应决定了其生物量的分配,盐分格局能够改变牧草各器官之间固有的物质与能量分配模式[21]。本试验中,匈牙利能源植物SZARVASI-1的株高、根长均随着NaCl胁迫的增加先升高后下降,在NaCl浓度为0.6%时有最大值,并且胁迫时间的延长不影响其总体变化规律,说明该植物对于浓度为0.6%的盐胁迫较为敏感。同時,其地上生物量、地下生物量、总生物量均随着盐浓度的上升先增加后减少,在NaCl浓度为0.6%时有最大值;胁迫60 d,在NaCl浓度为1.2%时,地上生物量较CK显著降低,但是地下生物量较CK显著增加,这与胁迫初期呈现相反的格局;同时根冠比达到了最大值,幼苗将更多的同化物分配给根系生长以适应环境,从而提高其耐盐性,这与前人在其他植物中的研究[22-23]相一致。该植物有可能通过增加地下部分或者减少地上部分的生长来调节自身对于外界胁迫的适应能力。地上部生物量、地下部生物量分配格局不仅受环境因子,如盐分、光照、温度等的影响,而且与植物自身的适应机制有关,根冠关系对环境因子的响应以遗传特性为基础[24]。当少量根系能维持环境因子间的供需平衡时,植物根系通过启动自我调控能力将更多的能量用于地上部分的生长;当外界胁迫加剧时,植物会增加根系数量、根系活力以及根冠比来应对环境,但是不能弥补高盐造成的产量下降。在盐碱地上种植牧草不但能减少土壤水分蒸发,某些耐盐碱牧草庞大的根系还可以改善土壤结构[25],因此牧草对盐碱地的改良具有重要作用。
总之,低浓度的盐胁迫对禾草的生长具有促进作用,但是超过一定浓度时将抑制禾草幼苗生长。本研究中,适宜匈牙利能源植物SZARVASI-1种子萌发的最适NaCl浓度为0.8%;适宜其幼苗生长的最佳NaCl浓度为0.6%,超过此浓度则产生抑制作用,植物通过增加地下部分的生长来适应逆境。当然,植物对逆境的响应是受多因素、多途径调控的复杂过程,基于该植物是多年生牧草,其耐盐碱能力经过长期适应可以逐渐训练,而本试验仅从种子萌发、幼苗生长方面对匈牙利能源植物SZARVASI-1的耐盐性进行了初步探讨,有关不同生长时期的生理反应及其耐盐机制等还有待于进一步深入研究。
参考文献:
[1]王银柱,王 冬,匡彦蓓,等. 不同水分梯度下能源植物芒草和柳枝稷生物量分配规律[J]. 草业科学,2015,32(2):236-240.
[2]桑 涛. 能源植物秀——芒草[J]. 生命世界,2011,225(1):38-43.
[3]Bartels D,Sunkar R. Drought and salt tolerance in plants[J]. Critical Reviews in Plant Sciences,2005,24(1):23-58.
[4]乔 旭,黄爱军,褚贵新. 植物对盐分胁迫的响应及其耐盐机理研究进展[J]. 新疆农业科学,2011,48(11):2089-2094.
[5]张俊叶,张力君,赵青山,等. 几种豆科牧草种子萌发期的耐盐性[J]. 中国草地学报,2012,34(4):116-120.
[6]李 珊,陈静波,郭海林,等. 结缕草属草坪草种质资源的耐盐性评价[J]. 草业学报,2012,21(4):43-51.
[7]王晓龙,米福贵,郭跃武,等. 不同禾本科牧草种子萌发及幼苗耐盐性鉴定[J]. 草原与草坪,2014,34(1):23-28.
[8]慈敦伟,丁 红,张智猛,等. 花生耐盐性评价方法的比较与应用[J]. 花生学报,2013,42(2):28-35.
[9]余 玲,王彦荣,孙建华. 野大麦种子萌发条件及抗逆性研究[J]. 草业学报,1999,8(1):50-57.
[10]王晓龙,李 红,闫利军,等. 5种禾本科牧草种子萌发及幼苗耐盐性鉴定[J]. 种子,2016,35(8):27-31.
[11]徐炳山,山 仑,黄 瑾,等. 柳枝稷和白羊草苗期水分利用与根冠比的比较[J]. 草业学报,2003,12(4):73-77.
[12]龚吉蕊,黄永梅,葛之葳,等. 4种杂交杨对土壤水分变化的生态学响应[J]. 植物生态学报,2009,33(2):387-396.
[13]乔 枫,罗桂花,耿贵工. 蚕豆幼苗对NaCl和NaHCO3胁迫的生理响应[J]. 安徽农业大学学报,2011,38(5):783-787.
[14]罗志娜,赵桂琴,刘 欢. 24个燕麦品种种子萌发耐盐性综合评价[J]. 草原与草坪,2012,32(1):34-38.
[15]王晓龙. 五种禾本科牧草生物学特性、农艺性状及抗逆性研究[D]. 呼和浩特:内蒙古农业大学,2014.
[16]武俊英,刘景辉,翟利剑,等. 不同品种燕麦种子萌发和幼苗生长的耐盐性[J]. 生态学杂志,2009,28(10):1960-1965.
[17]Baskin C C,Baskin J M. Seeds:ecology,biogeography,and,evolution of dormancy and germination[M]. Amsterdam:Elsevier,1998.
[18]史燕山,駱建霞,黄家珍,等. 盐胁迫对7种草本地被植物种子萌发的影响[J]. 天津农学院学报,2007,14(4):1-4.
[19]田晓艳,刘延吉,张 蕾,等. 盐胁迫对景天三七保护酶系统、MDA、Pro及可溶性糖的影响[J]. 草原与草坪,2009(6):11-14.
[20]肖 强,郑海雷,陈 瑶,等. 盐度对互花米草生长及脯氨酸、可溶性糖和蛋白质含量的影响[J]. 生态学杂志,2005,24(4):373-376.
[21]李旭东,张春平,傅 华. 黄土高原典型草原草地根冠比的季节动态及其影响因素[J]. 草业学报,2012,21(4):307-312.
[22]郭慧娟,胡 涛,傅金民,等. 苏打碱胁迫对多年生黑麦草的生理影响[J]. 草业学报,2012,21(1):118-125.
[23]孟 林,尚春艳,毛培春,等. 偃麦草属植物种质材料苗期耐盐性综合评价[J]. 草业学报,2009,18(4):67-74.
[24]徐炳成,山 仑,黄 瑾,等. 柳枝稷和白羊草苗期水分利用与根冠比的比较[J]. 草业学报,2003,12(4):73-77.
[25]刘延吉,张珊珊,田晓艳,等. 盐胁迫对NHC牧草叶片保护酶系统,MDA含量及膜透性的影响[J]. 草原与草坪,2008(2):30-34.
猜你喜欢
今日农业(2020年19期)2020-12-14
山东农业科学(2016年12期)2017-01-21
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
中国烟草学报(2012年3期)2012-04-10
