
不同海拔梯牧草生物型种群构件的生物量分配策略
2021-07-26 05:17张鲜花李江艳袁小强朱进忠
草原与草坪 2021年3期
张鲜花,李江艳,袁小强,朱进忠
(新疆农业大学草业与环境科学学院,新疆草地资源与生态实验室,新疆 乌鲁木齐 830052)
自Harper等[1]提出植物种群构件结构理论,植物构件间性状及其相关性的研究在草学、农学、林学、园艺学等领域种质资源的开发利用中被广泛应用[2-8]。植株构件的大小不仅能够反映植物个体及种群的生长发育节律,也能反映其对环境的适应能力,进而体现了植物个体及种群适应环境的生存策略。同时,研究者也能通过植物构件性状的变化,了解受损生态系统中群落功能群植物的演变过程与趋势[9]。例如,对不同湿地芦苇分株构件的生物量动态及分配规律研究表明,芦苇分株构件生物量积累具有遗传因素控制、稳定的生物学特性,同时也具有受环境因素影响的生态学特性[10]。再如对新疆巴音布鲁克高寒草原建群种紫花针茅(Stipapurpurea)的研究表明,随着退化程度的增加,紫花针茅种群高度和个体生物量显著降低,分蘖节和茎的直径呈现增加的趋势[11]。
梯牧草(Phleumpratense),又名猫尾草,系禾本科梯牧草属植物,是世界上广为栽培的一种优良牧草。其野生种主要分布在N 40°~50°寒冷潮湿地区,在我国仅分布于新疆天山中西段的山地[12],分布范围与数量十分有限,抢救性地对其开展一些基础研究迫在眉睫。野生梯牧草种群在不同生境条件下植株形态特征(如株高、茎粗、叶片的长与宽、花序大小等构件)差异较大,在生境条件较好地段,表现出植株高大、叶量丰富、花序较长等特点[13]。目前对梯牧草的研究,国内文献大多集中在引种及分子育种等方面[14-19],构件方面的研究较少。鉴于此,本研究以分布于不同海拔地段的野生梯牧草为试验材料,对比分析不同海拔梯度下种群的构件及生物量分配规律,为揭示梯牧草的生态适应性提供基础资料,也为合理开发利用与保护野生种质资源提供科学依据。
1 材料和方法
1.1 试验材料
试验材料取自天山西段伊犁哈萨克自治州境内的昭苏县,地处乌松山中西段和昭苏盆地,地理位置为E 80°40′~80°56′,N 43°04′~43°08′,海拔1 800~2 200 m。气候属温带大陆性山区湿润冷凉型气候,年均温2.9℃,年降水量500~700 mm,春、秋、冬降水较多,时有夏旱发生。梯牧草种群分布及草地类型为禾草+杂类草的山地草甸,与梯牧草共生的主要植物有鸭茅(Dactylisglomerata)、无芒雀麦(Bromusinermis)、曲芒鹅观草(Roegneriatschimganica)、圆柱披碱草(Elemusdahurius)、细叶早熟禾(Poaangusitifolia)、偃麦草(Elytrigiarepens)、小糠草(Agrostisgigantea)、红车轴草(Trifoliumpratense)、线叶野豌豆(Viciatenuifolia)、黄花委陵菜(Potentillachrysantha)、蓬子菜(Galiumverum)、直立老鹳草(Geraniumrectum)等。
1.2 方法
1.2.1 样地设置 在梯牧草集中分布区域,按照海拔梯度的不同,设置5个样地(表1)。
1.2.2 采样及测定方法 2019年8月牧草结实期,每隔100~200 m的海拔梯度设置1个调查样地,样地面积100 m×100 m,每个样地中设置3条100 m的样线,样线间隔30 m;在每条样线随机挖取健康、完整植株20株,每个样地总计取样60株,依次分株编号,测定株高后,带回实验室,将根、茎、叶、穗进行分离,并置于纸袋中,于105℃杀青30 min,80℃下烘干24 h,采用分析天平称其干重,记录每株植物各构件的重量,并计算:
(1)根冠比=地下生物量/地上生物量
(2)构件生物量分配(%)=(构件生物量/全株的总生物量)×100%;
(3)营养分配(%)=(茎重+叶重)/全株总生物量×100%;
(4)贮藏分配(%)=(根重/全株总生物量)×100%;
(5)繁殖分配(%)=(花序重/全株总生物量)×100%。
1.3 数据处理
采用SPSS 17.0对不同海拔种群构件生物量差异进行比较分析,对株高、营养分配和繁殖分配之间进行相关性分析;采用单因素方差比较分析,进行显著性检验、相关性检验及回归分析。各构件生物量与单株生物量大小之间关系采用幂函数进行曲线拟合,判断生长趋势。其中异速生长关系表示为Y=β·Xа,式中Y是梯牧草构件生物量大小,β是标准化常数,X是单株生物量,а为异速生长指数。若а=1为等速关系,а≠1为异速关系,若а<1,则为显著的异速生长关系。
2 结果与分析
2.1 不同海拔梯牧草的构件生物量
梯牧草的构件生物量大小依次为茎>根>叶>穗,且各海拔之间均存在显著差异(P<0.05)(表2)。各构件生物量随海拔升高呈现先降低后升高再降低的趋势,总体表现为海拔2 000 m处最大,海拔1 900 m处最小。根冠比随海拔升高,呈现下降的趋势。
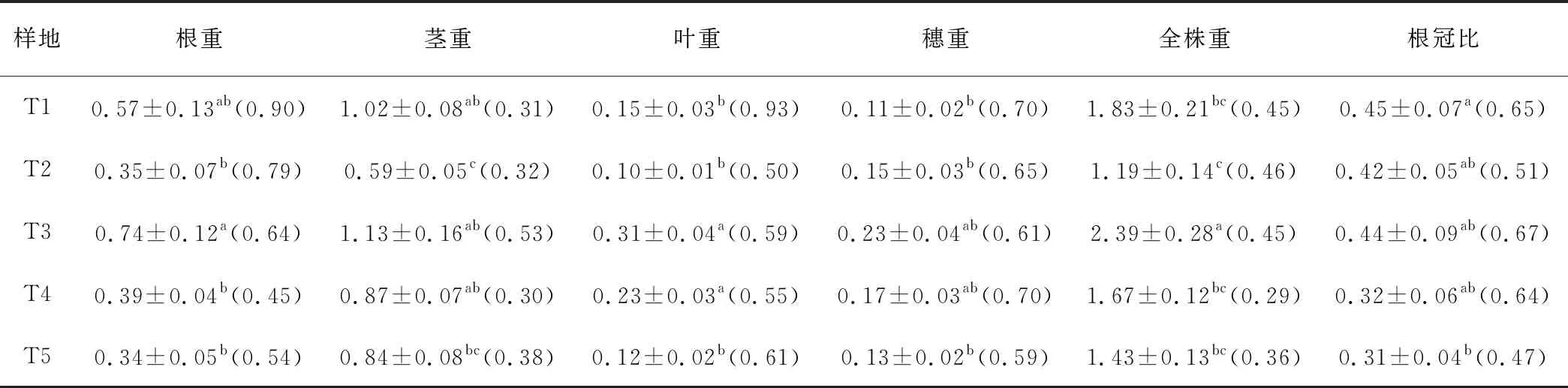
表2 梯牧草个体各构件生物量组成
根生物量海拔2 000 m处最大,其次为1 800 m,二者均显著高于其他海拔处(P<0.05), 2 200 m处最小;茎生物量在各海拔表现出与根生物量相同的趋势,海拔2 000 、1 800和2 100 m处均显著高于1 900 m和2 200 m处(P<0.05);叶生物量海拔2 000 m和2 100 m处显著高于其他海拔,1 900 m处最小(P<0.05);穗生物量表现出与叶生物量一致的趋势。全株生物量海拔2 000 m显著高于其他海拔(P<0.05),大小顺序为海拔2 000 m>1 800 m>2 100 m>2 200 m>1 900 m。
根变异系数介于0.45~0.90,茎变异系数介于0.30~0.53,叶变异系数介于0.50~0.93,穗变异系数介于0.59~0.70。不同海拔地段梯牧草各构件变异系数均大于0.20,有一定的变异性。
2.2 不同海拔梯牧草的构件生物量分配
梯牧草个体各构件生物量分配,依次表现为营养分配>储藏分配>繁殖分配,且在各海拔之间表现出一定的差异性(表3)。
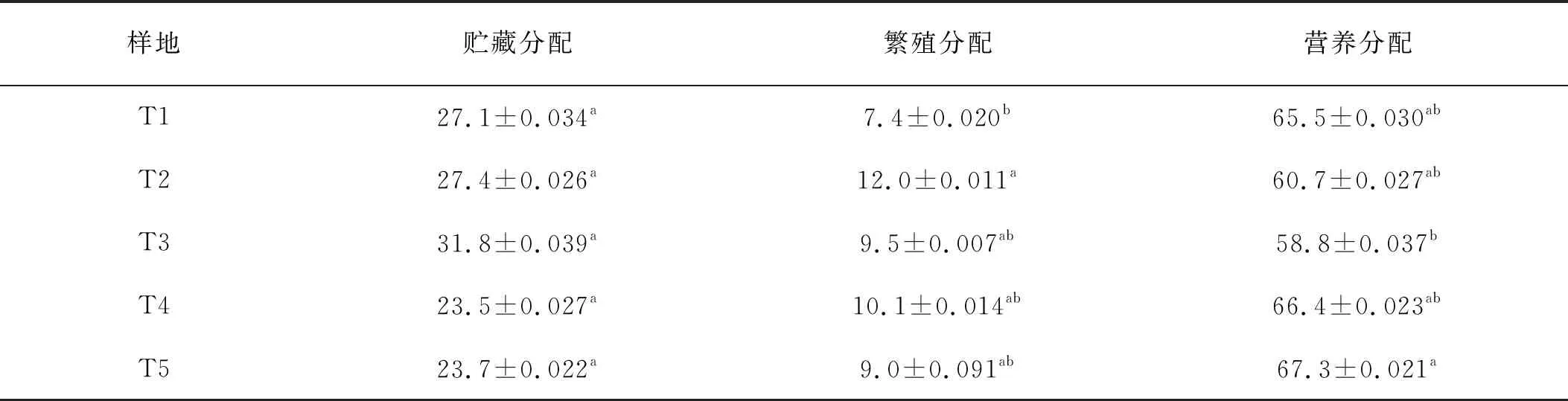
表3 梯牧草个体各构件生物量分配
营养分配,海拔2 200 m最大,与海拔2 000 m具有显著差异(P<0.05),但与其他海拔差异不显著。储藏分配,在各海拔之间未呈现出显著的差异,海拔2 000 m处最大,2 100 m处最小;繁殖分配,海拔1 900 m处显著高于1 800 m处,但与其他海拔之间差异不显著。
2.3 不同海拔梯牧草构件生物量与株高的关系
株高与茎生物量在各海拔的关联性均呈极显著正相关(P<0.01),相关系数介于0.567~0.772。根、叶、穗生物量与株高关联性在不同海拔地段的强弱不同,在海拔1 800 m处,株高除与茎生物量呈极显著正相关,和根生物量呈显著相关外,与叶和穗关联性较弱;海拔1 900 m处,株高与茎呈极显著正相关,与根和穗生物量呈显著正相关,与叶生物量关联性不显著;而海拔2 000 m和2 100 m处,与株高关联性最强的为茎、其次为穗,叶和根关联性不强;海拔2 200 m处,呈极显著正关联的为茎生物量,根、叶和穗生物量关联性不显著(表4)。

表4 梯牧草个体各构件生物量与株高的关系
2.4 不同海拔梯牧草构件生物量分配与总生物量的关系
不同海拔根、茎、叶及穗生物量与全株生物量呈幂函数的正相关关系,表现出异速生长的规律(图1-5)。其中海拔1 800 m处,根、茎及叶生物量与全株生物量之间呈现极显著正相关,而穗部对全株生物量影响较小;海拔1 900 m与2 000 m处,根、茎、穗与叶生物量与全株生物量之间均呈极显著正相关或显著相关,说明各构件大小均在不同程度上影响全株生物量的大小;在海拔2 100 m处,茎和叶生物量均与全株生物量呈极显著正相关,而穗生物量和全株生物量呈显著正相关,根对全株生物量的影响较小;海拔2 200 m处,除茎与全株生物量呈极显著的正相关外,其他构件与全株生物量无显著相关性。

图1 海拔1 800 m梯牧草个体构件生物量分配与全株总生物量的关系

图2 海拔1 900 m梯牧草个体构件生物量分配与全株总生物量的关系

图3 海拔2 000 m梯牧草个体构件生物量分配与全株总生物量的关系

图4 海拔2 100 m梯牧草个体构件生物量分配与全株总生物量的关系

图5 海拔2 200 m梯牧草个体构件生物量分配与单株总生物量的关系
3 讨论
植物种群构件生物量分配策略是优异种质选择与评价的主要依据[20]。影响植物构件生物量的因素较多,如植物自身的特性,环境因子等[21-22]。本研究重点探讨不同海拔梯度对梯牧草各构件生物量形成的影响以及对全株生物量的贡献问题。研究结果表明,海拔与梯牧草种群各构件生物量之间的关系,没有表现出随着生长地海拔的升高或降低各构件生物量相应的递增或递减,各构件生物量的大小主要取决于生长地的水热条件。在调查区域内,海拔2 000~2 100 m山地是降水最大的地段,年均降水量可达到650~700 mm,植物生长旺季的年均温度基本在25℃以上,其水热条件十分有利于梯牧草的生长,种群构件生物量也达到最大;随着海拔的升高,水、热条件均有所变化,种群各构件的生物量也随之降低;海拔1 800 m地段处于草地垂直带谱的最底端,位于山间盆地最低处,年降水量虽然只有500 mm,但在植物生长期除了可接纳自然降水外,还伴有地下水的补给,形成了局地潮湿和高温的生境,对促进梯牧草的生长发育有一定作用,与地带性分布规律有较大的差异。梯牧草在1 800 m处的生长表现,是草地非地带性分布水分因子发生根本改变的结果,而其他海拔间产生的差异,是梯牧草适应环境的体现。同时也说明,在地带性草地生境条件下,海拔2 000~2 100 m的地段最适宜梯牧草生长,至于某些构件上表现出的差异应与梯牧草着生的微地形有关,如小尺度的地形起伏,局部水土富集形成的微环境,都会在同一海拔地段使梯牧草形成不同的长势,这与在人工栽培条件下观察的结果有较大的不同。本研究与李江艳等[23]对该地区不同海拔下鸭茅形态特征变化的研究,张丽辉等[24]对三裂叶豚草在两种生境条件下种群差异的研究结论一致。
梯牧草生物量分配均表现为营养分配大于繁殖分配,表明在籽实成熟期,各生境下梯牧草表现出相同的营养分配与繁殖分配策略,即梯牧草仍然需要分配较多的资源给光合器官,使植物在停止生长前产生更多的有机物,以保证植物体能量与物质的继续积累。研究结果说明,生境的差异导致植株个体对种群各构件进行能量分配时是按照不同构件的贡献率进行的,这与刘秀香和杨允菲[25]、张德魁等[26]的研究结果一致。
异速生长关系是生物量分配和器官功能关系研究的重要指标。从异速生长变化上来看,各海拔地段幂函数的指数均不等于1,表明不同海拔地段各构件生物量大小与全株生物量之间存在异速生长关系。在不同海拔地段,有呈显著或不显著的异速生长变化。由此说明,梯牧草在生长过程中,各构件呈现从加速到缓速的异速生长规律,这是植物个体适应生境调节策略的一个表现。本研究结果与陶冶[27]的研究结果一致,说明植物个体构件生物量的分配受植物的遗传特性、环境条件和个体大小的影响。
4 结论
(1)在不同海拔地段,梯牧草各构件生物量大小均表现为茎>根>穗>叶,生物量分配上营养分配>储藏分配>繁殖分配,且在各海拔之间表现出差异性。
(2)不同海拔地段,各构件对梯牧草生物量的贡献率不同,并受局地生境变化的影响;海拔2 000 m地段是梯牧草生长发育的最适地段,各构件生物量对株体生物量的贡献率均最大。
(3)各海拔条件下,梯牧草种群根、茎、叶及穗生物量与单株生物量呈幂函数的正相关关系,各构件生物量均随单株大小的增加而增加。
(4)梯牧草在生长过程中,构件大小与单株个体大小呈异速生长关系,在不同海拔地段,呈显著或不显著的异速生长变化。
猜你喜欢
散文诗(2021年22期)2022-01-12
今日农业(2021年10期)2021-07-28
今日农业(2020年19期)2020-12-14
环球时报(2019-05-23)2019-05-23
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20
中国科技信息(2015年2期)2015-11-16
西藏科技(2015年11期)2015-09-26
植物营养与肥料学报(2014年1期)2014-03-11
郑州大学学报(工学版)(2014年6期)2014-03-01
当代畜禽养殖业(2014年1期)2014-02-27
