
鄱阳湖大型底栖动物时空演变特征及驱动因素
2021-07-22 12:11:48邹亮华张庆吉龚志军张艳杰鲁顺保蔡永久
中国环境科学 2021年6期
邹亮华,邹 伟,张庆吉,3,李 颖,4,龚志军,张艳杰,鲁顺保*,蔡永久**
鄱阳湖大型底栖动物时空演变特征及驱动因素
邹亮华1,2,邹 伟2,张庆吉2,3,李 颖2,4,龚志军2,张艳杰1,鲁顺保1*,蔡永久2**
(1.江西师范大学生命科学学院,江西 南昌 330027;2.中国科学院南京地理与湖泊研究所,中国科学院流域地理学重点实验室,江苏 南京 210008;3.南京大学地理与海洋科学学院,江苏 南京 210023;4.西南大学地理科学学院,重庆 400715)
于2019年7月对鄱阳湖133个样点大型底栖动物进行调查,分析其群落结构与环境因子之间的关系.结果表明,共记录底栖动物48种,隶属7纲16目22科38属,全湖底栖动物的平均密度和生物量分别为158.38ind/m²和173.76g/m²,现阶段优势种主要为河蚬、铜锈环棱螺和大沼螺.将全湖分为5个湖区:北部湖区、西北部湖区、中部湖区、东部湖区和南部湖区,单因素方差分析表明不同湖区水体理化指标差异显著(ANOVA,<0.05),沉积物环境指标差异不显著.相似性分析结果表明不同湖区底栖动物群落结构差异性显著,北部湖区生物多样性显著高于其它湖区.典范对应分析表明水深、溶解氧、浊度、总磷、叶绿素a、烧失量和底质类型与底栖动物群落结构显著相关.与历史研究相比,鄱阳湖底栖动物多样性下降明显,优势种由大型软体动物逐渐演变成小型软体动物和昆虫类.鄱阳湖采砂、水文情势变化和水生植被衰退是影响鄱阳湖大型底栖动物群落结构演替的主要环境因素.
鄱阳湖;大型底栖动物;群落结构;生物多样性;环境因子
底栖动物是指生活史的全部或大部分时间生活于水体底部的无脊椎动物,是维系水生生物多样性、水生态系统结构和功能的重要组成部分[1].底栖动物可加速水底碎屑的分解、调节泥水界面的物质交换及促进水体的自净等作用,同时还是水生态系统食物链的重要环节[2].底栖动物具有分布范围广、生命周期长和迁移能力弱、不同物种对环境变化的敏感程度不同等特点[3],因此被广泛运用到水生态评价中.随着我国对流域水生态管理的日益重视,水生态系统健康的研究也更加深入,底栖动物作为生态监测的关键类群,需要开展进一步的研究[4].
鄱阳湖位于长江中游,江西省北部,是中国第一大淡水湖,正常水位(14~15m)情况下面积达3914km2.鄱阳湖是典型的吞吐性、季节性的湖泊,与赣江、抚河、信江、饶河、修水相接,调蓄后经湖口注入长江,水位涨落变化较大[5].近年来受上游大型水利工程和流域来水影响,改变了长江和鄱阳湖之间的江湖关系,从而导致鄱阳湖生态系统格局出现巨大变化[6].此外,近年来鄱阳湖氮、磷污染越来越严重,水体富营养化也进一步加剧,水环境质量不断下降[7],而底栖动物对环境的变化十分敏感,其群落结构往往会随着水环境的变化发生改变[8].
对鄱阳湖底栖动物的调查与研究已有一些报道,20世纪80年代初,谢钦铭等[9]对鄱阳湖底栖动物有过定性定量调查,调查样点共20个,记录底栖动物95种;20世纪90年代,Wang等[10]对鄱阳湖底栖动物进行定量调查,调查样点共18个,记录底栖动物58种;欧阳珊等[11]于2007、2008年在鄱阳湖湖区分别调查了43个和32个样点,采集到大型底栖动物35种;Cai等[12]在2012年对鄱阳湖底栖动物进行4个季度调查,调查样点共15个,采集到大型底栖动物41种,并结合历史研究分析表明底栖动物优势种发生明显变化.然而以往研究均表现出空间分辨率低的特点,难以全面反映鄱阳湖底栖动物群落结构特征,为此本研究根据鄱阳湖丰水期水域面积布设了133个样点,系统调查鄱阳湖大型底栖动物群落及环境因子,探讨鄱阳湖底栖动物群落结构的空间异质性及其驱动因子.并与历史研究进行比较,揭示其底栖动物群落结构的长期变化,以期为评价鄱阳湖水生态系统健康状况及影响因素提供科学支持.
1 材料与方法
1.1 研究区域概括
鄱阳湖(28°22′~29°45′N和115°47′~116°45′E)是我国最大的淡水湖,也是目前长江中游仅存的2个大型自然通江湖泊之一,位于江西省北部长江南岸.鄱阳湖地处我国亚热带湿润气候区,气候温和,降雨充沛,日照充足,年均气温16.5~17.8℃,7月平均气温28.4~29.8℃,多年平均年降水量1570mm.鄱阳湖承载赣江、抚河、信江、饶河、修水及漳田河等来水,经调蓄后由湖口注入长江,是一个过水性、吞吐型、季节性的湖泊,其具有“高水湖相,低水河相”的独特自然地理景观,其丰、枯水位面积相差10倍[13].鄱阳湖的水位受到支流和湖盆、北部长江的影响,鄱阳湖有明显的季节性水位阶段,12月到次年3月鄱阳湖星子站水位低于10m(吴淞),湖区面积通常减少到不到1000km².4~6月,水位增加至14m,丰水期7~9月,水位高于14m,湖泊面积超过4000km2,10~11月,水位急剧下降至10m左右,并在12月至次年3月保持在低水位阶段.作为少数几个自由连接长江的湖泊之一,鄱阳湖在维持长江洪泛生态系统独特生物多样性方面发挥着重要作用[14].
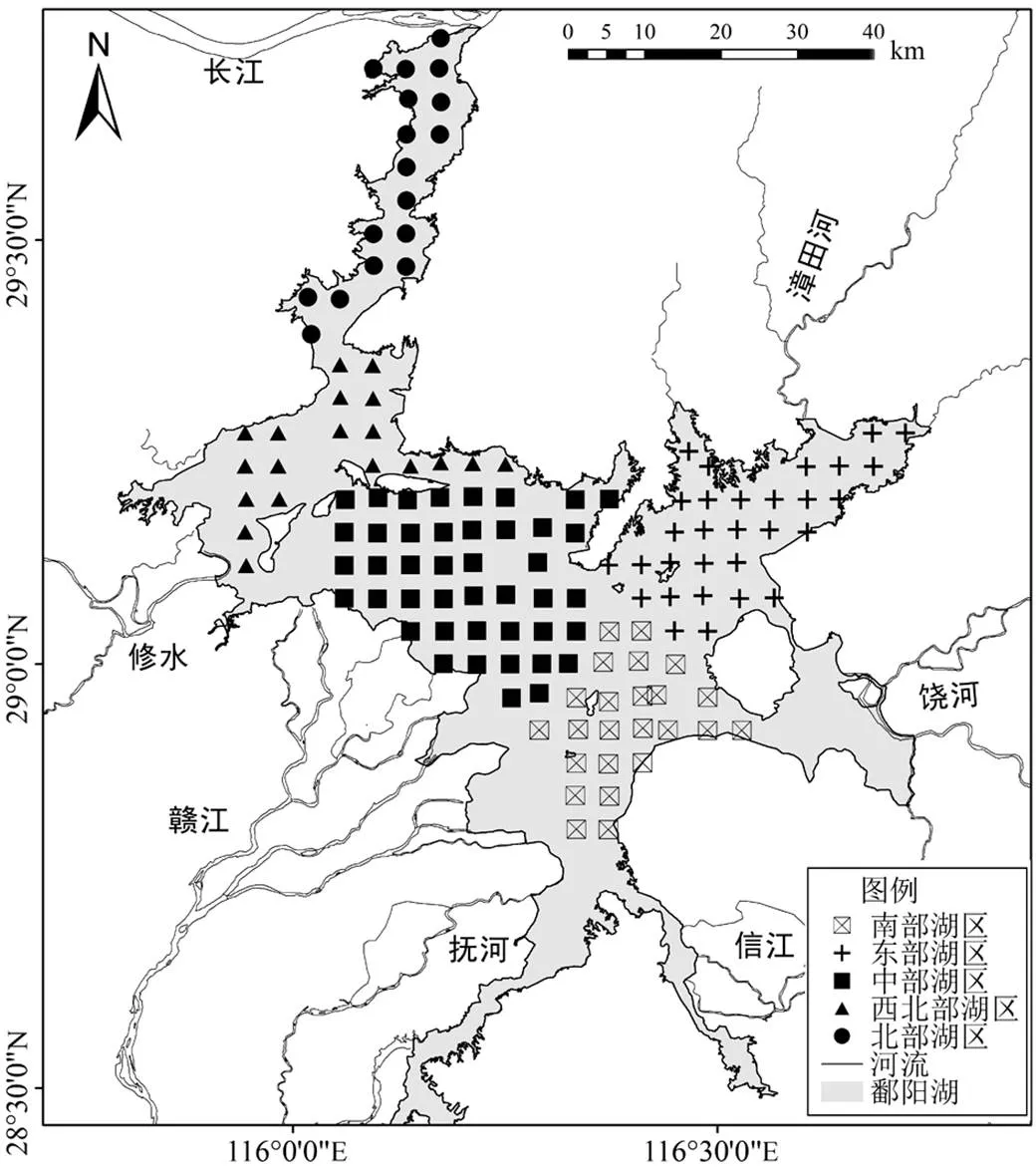
图1 鄱阳湖采样点分布
根据鄱阳湖的湖泊形态、水动力和生境条件等,把鄱阳湖分为5个湖区,共设置了133个样点(图1),分别为北部湖区(=17)、西北部湖区(=19)、中部湖区(=43)、东部湖区(=30)和南部湖区(=24).北部湖区为通江水道区域,湖面狭窄,流速较快,水较深,受采砂活动影响较多;西北部湖区为半封闭宽阔湖面,流速不均匀,河流入湖口,部分区域有采砂活动;中部湖区湖面宽阔,流速较快;东部湖区为湖湾区域,流速缓慢,水面相对静止,撮萁湖区域有大量的养殖活动;南部湖区流速缓慢,河流入湖口较多,河流带来的污染物较多.
1.2 环境因子测定和样本采集
2019年鄱阳湖丰水期时,进行野外环境因子测定和样品采集.现场使用测深仪测定水深(WD),使用多参数水质监测仪(YSI 6600V2)测定pH值、电导率(Cond)、溶解氧(DO)、浊度(Tur),同时采集水样和表层沉积物样品并冷藏保存带回实验室.现场使用0.0625m2的改良彼得森采泥器,在每个采样点附近3个不同位置分别抓取1次合成1个样品,所采的泥样经60目尼龙筛网筛洗干净后,剩余物冷藏带回实验室进行分拣.在实验室中将剩余物放在白色解剖盘中,将底栖动物逐一挑出,样本用10%福尔马林保存.利用解剖镜和显微镜将样本都鉴定至尽可能低的分类单元[15-17],然后用滤纸吸去表面固定液,置于万分之一电子天平上称重,将结果换算成单位面积的密度(ind/m2)和生物量(g/m2).
在实验室内测定水体铵氮(NH4+-N)、硝酸盐氮(NO3--N)、正磷酸盐(PO43-P)、溶解性有机碳(DOC)、总氮(TN)、总磷(TP)、高锰酸盐指数(CODMn)和叶绿素a(Chl-a)浓度;测定沉积物总氮(TNs)、总磷(TPs)、烧失量(LOI)和底质类型组成,其中底质类型组成按黏土(<4µm)、细粉砂(4~16µm)、粗粉砂(16~63µm)、砂(>63µm)的所占质量百分比划分.营养盐参数的测定方法参考《水和废水监测分析方法》[18].
1.3 数据分析
1.3.1 优势种 优势种是指群落中占优势的且对整个群落具有控制性影响的种类[19],本文采用相对重要性指数(IRI)进行优势种的确定,其计算公式为:

式中:为某一种类的生物量占各点大型底栖动物总生物量的比例,%;为该种类的密度占各点总密度的比例,%;为该种类在各湖区出现的相对频率.
1.3.2 多样性指数 本文运用Simpson优势度指数()、Margalef丰富度指数(ma)、Shannon-Wiener多样性指数()及Pielou均匀度指数()这4个多样性指数计算各样点的底栖动物多样性.
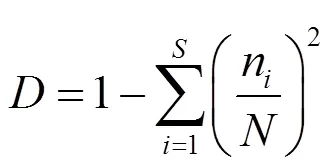

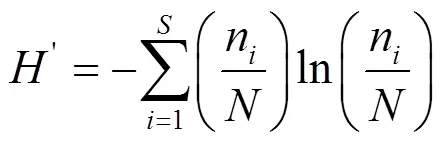
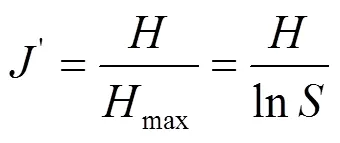
式中:为大型底栖动物群落总物种数;为大型底栖动物群落总个体数;n为第个物种的个体数
1.3.3 统计分析 为提高数据的正态性和方差齐性,将密度、生物量与所有水环境因子(除pH值外)数据进行log(+1)转换.使用单因素方差分析(one-way ANOVA)比较各湖区环境指标间是否存在显著差异.采用主成分分析(PCA)分析理化数据,将各环境因子重新进行降维,提取出可以尽可能多的反映原来变量信息的综合变量,寻找环境因子的潜在梯度.
利用相似性分析(ANOSIM)检验不同湖区的动物群落组成的差异显著性,利用相似百分比(SIMPER)找出对底栖动物群落差异贡献最大的物种,找出各湖区的特征种及其相应的贡献率.采用典范对应分析(CCA)分析底栖动物群落与环境因子之间的关系,首先使用向前引入法对鄱阳湖环境因子进行筛选,保留能通过蒙特卡罗置换检验的显著因子(<0.05).
单因素方差分析在SPSS 22中完成,ANOSIM分析和SIMPER在PRIMER 5.0中完成,PCA和CCA在R语言Vegan包中完成,底栖动物的空间分布图使用ArcGIS 10.7绘制.
2 结果与分析
2.1 环境因子特征
由图2、表1可见,One-way ANOVA表明水体理化指标在各湖区均表现出显著差异(<0.05).北部湖区的平均水深为11.36m,浊度为20.04NTU, NO3--N浓度为0.66mg/L,均显著高于其它湖区(<0.05),但电导率和pH值显著低于其它湖区(<0.001).南部湖区DO平均值为9.83mg/L,显著高于其它湖区(<0.01).西北部湖区的PO43-P、NH4+-N、CODMn、DOC浓度显著高于其它湖区(<0.05).

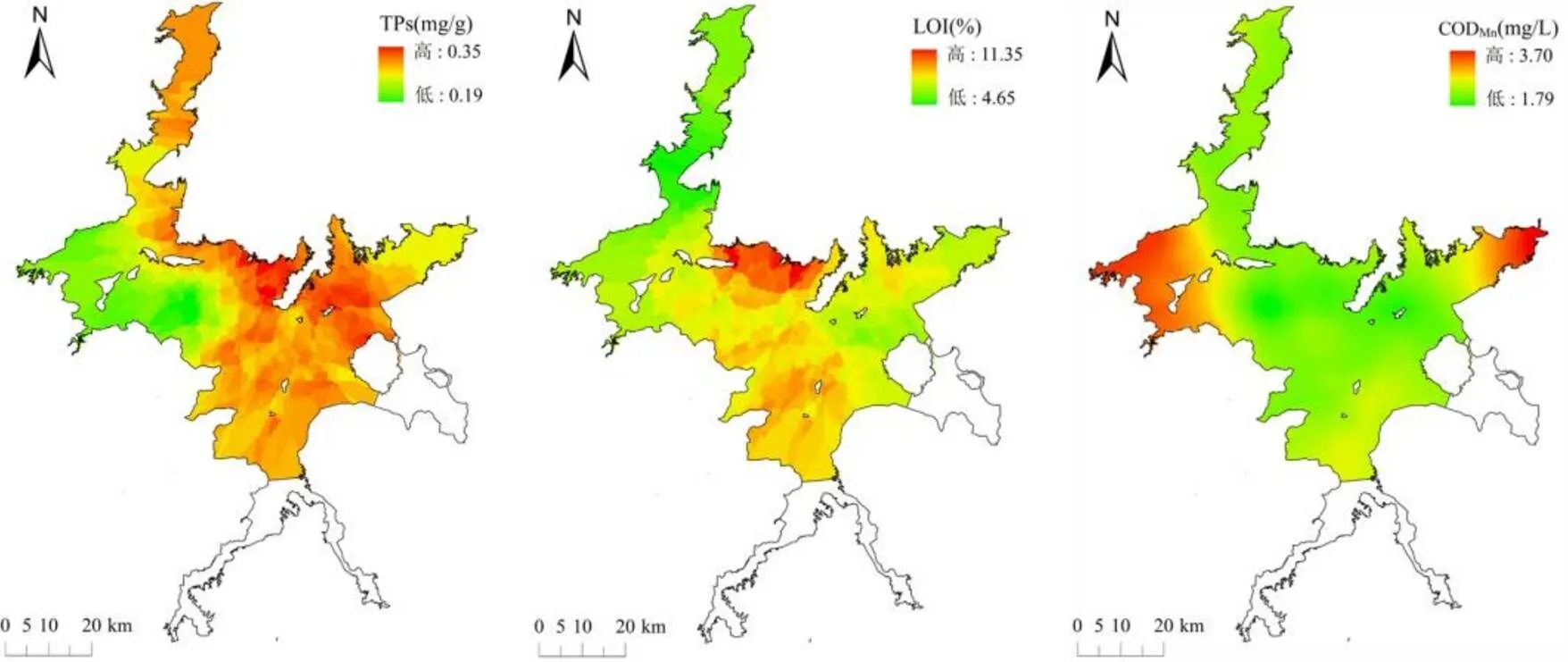
图2 鄱阳湖水体和沉积物理化参数空间格局
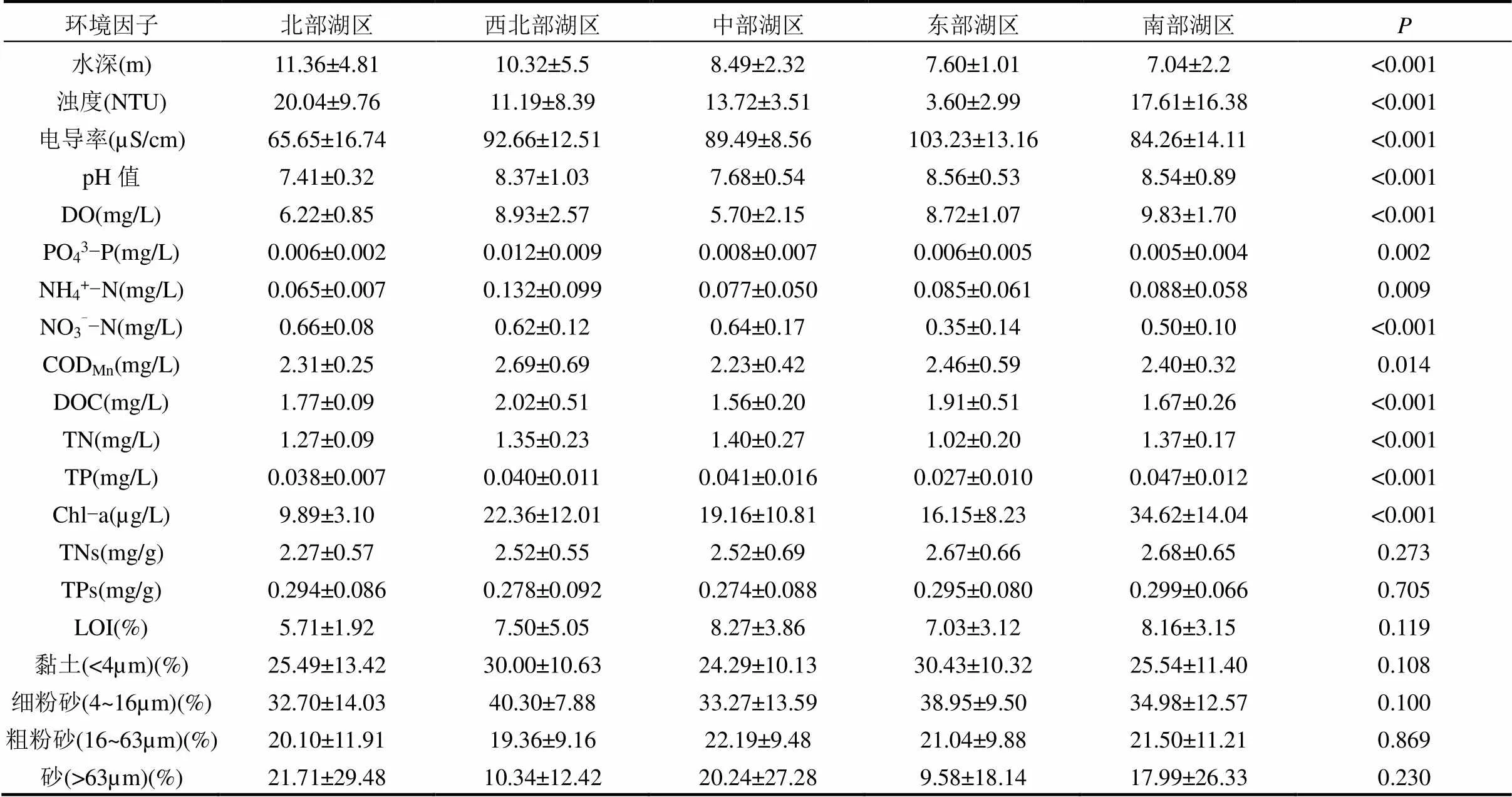
表1 鄱阳湖各湖区水体和沉积物理化参数
中部湖区的TN浓度为1.40mg/L,显著高于其它湖区(<0.05).南部湖区的TP浓度为0.047mg/L, Chl-a浓度为34.62µg/L,显著高于其它湖区(< 0.001).沉积物理化指标方面,不同湖区间无显著差异(>0.05),南部湖区的TNs、TPs平均值最高,中部湖区的LOI平均值最高.各湖区底质粒径组成中粗粉砂占比较为均匀,西北部湖区和东部湖区黏土、细粉砂占比较高,而北部湖区、中部湖区和南部湖区砂占比较高,接近于其它2个湖区的2倍(图3).
主成分分析显示第一主成分与电导率、DOC、CODMn、DO、pH值、TNs、黏土、细粉砂、粗粉砂呈负相关,与NO3--N、浊度、水深、砂呈正相关.第二主成分与Chl-a、TP、TN、PO43--P呈正相关,与LOI、TPs呈负相关(图4).不同湖区样点在主成分排序图上能较好地区分,东部湖区主要分布于第三象限,沉积物氮磷和有机质含量高,黏土和淤泥含量高.北部湖区的采样点主要分布于右侧,具有较高的水深和浊度,较低的电导率、有机污染和营养水平,沉积物砂含量高.西北湖区的洲滩区域样点集中分布在第二象限,水体中DOC、CODMn、NH4+-N浓度较高,表明有机质浓度较高.南部湖区样点主要分布于排序图的中间区域,表明各项指标值从全湖来看处于中等水平.中部湖区样点在排序图中分布较为分散,表明湖区内部理化因子变异较大.
2.2 种类组成与优势种

图3 鄱阳湖底质粒径组成空间格局
共鉴定到大型底栖动物48种,隶属7纲16目22科38属,其中软体动物腹足纲4科6属8种(16.7%),双壳纲5科10属12种(25%),环节动物寡毛纲1科2属2种(4.2%),多毛纲4科4属4种(8.3%),蛭纲2科2属2种(4.2%),节肢动物甲壳纲2科2属2种(4.2%),昆虫纲中摇蚊科幼虫9属15种(31.3%),其他类3种(6.3%).北部湖区物种丰富度相对较高,共采集到底栖动物38种,其次为中部湖区和南部湖区,为29种,东部湖区物种数为24种,而西北部湖区物种数最少,为21种.在所有底栖动物中,出现率最高的是铜锈环棱螺(42.65%),其次分别是河蚬(31.62%)、大沼螺(23.53%)、纹沼螺(16.91%)、长角涵螺(13.97%)、寡鳃齿吻沙蚕(13.24%).根据鄱阳湖大型底栖动物的IRI值表明,河蚬、铜锈环棱螺和大沼螺在各湖区均占据优势地位,为现阶段的优势种(表2).
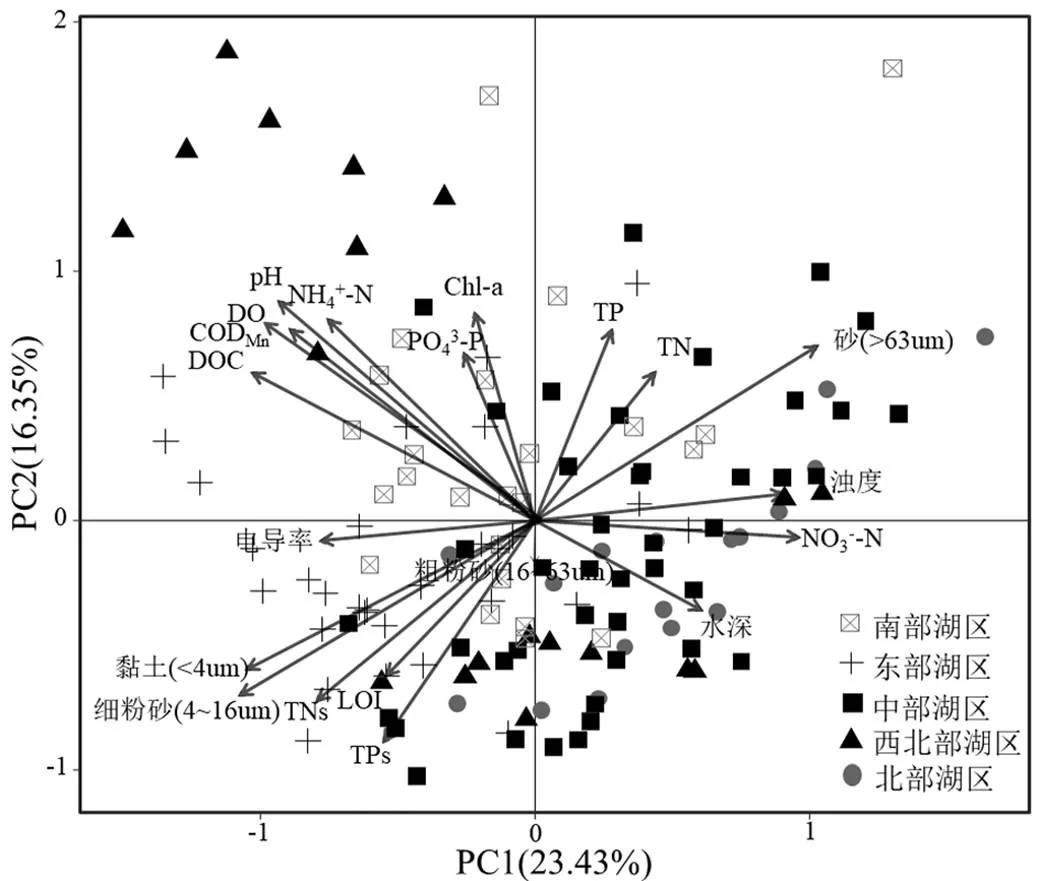
图4 鄱阳湖环境因子主成分分析
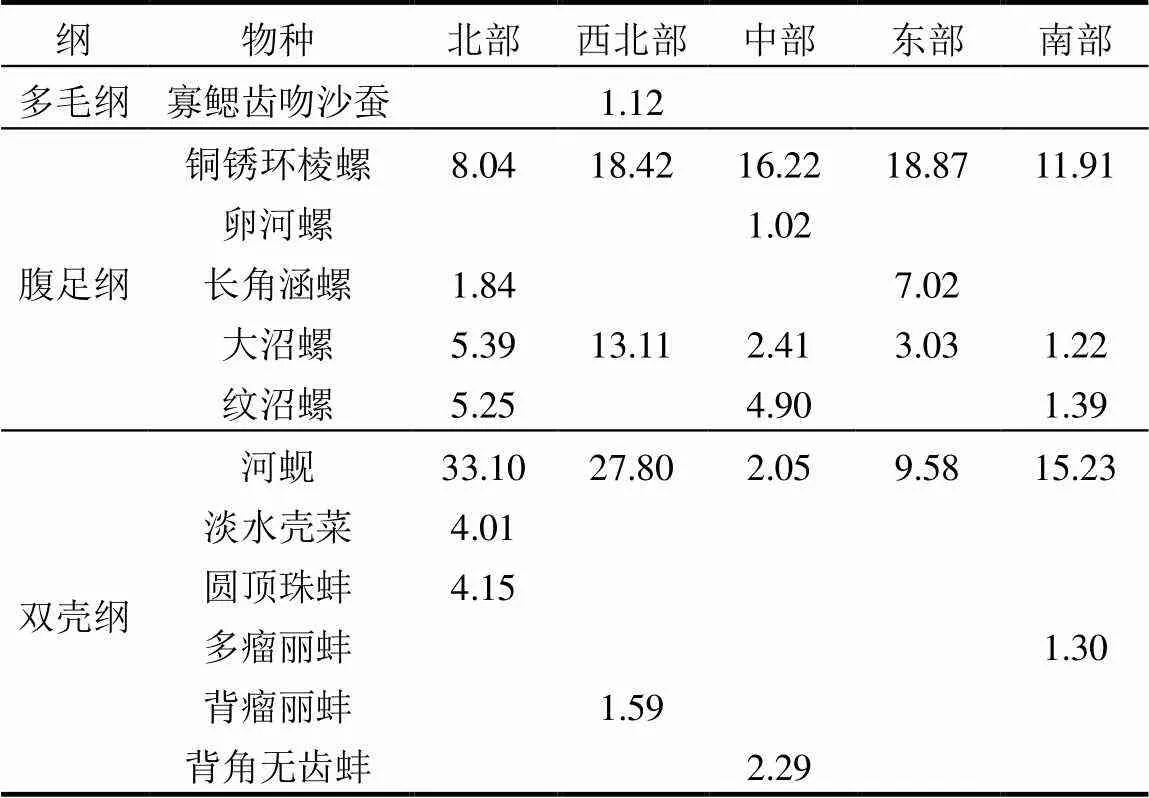
表2 鄱阳湖不同湖区大型底栖动物的优势种及其相对重要性指数得分
注:表中数值为相对重要性指数(IRI),且仅列出IRI>1的物种.
2.2 密度和生物量
全湖大型底栖动物的平均密度和平均生物量分别为158.38ind/m2和173.76g/m2.大型底栖动物密度空间差异较大(图5),最高值出现在西北湖区的样点,达到1897.14ind/m2,西北湖区的平均密度要高于其它湖区,为333.34ind/m2,其次为北部湖区,平均密度为288.21ind/m2,而中部湖区平均密度最低,仅为85.03ind/m2,东部湖区与南部湖区平均密度较为接近,分别为110.03和119.74ind/m2.大型底栖生物量空间变化也较大,最高值出现在南部湖区的一个样点,达到4678.11g/m2,与密度分布相同,西北湖区的平均生物量最高,为354.41g/m2,中部湖区最低,仅为65.67g/m2,而南部湖区、北部湖区、东部湖区分别为256.96,220.94,120.99g/m2.从不同类群所占比重可以看出,密度方面,腹足纲和双壳纲是优势类群,平均密度分别为78.29,38.53ind/m2,昆虫纲也占据一定比重,平均密度为23.07ind/m2.生物量方面,双壳纲和腹足纲主导了全湖生物量的空间变化,平均生物量分别为119.06g/m2和54.00g/m2,主要由于软体动物个体相对较大.
软体动物在各湖区均有出现,主要分布在湖区内部区域,而沿岸区域分布较少.铜锈环棱螺、大沼螺、河蚬和淡水壳菜是软体动物的优势种,占软体动物总密度和总生物量的80.14%和64.33%.其中铜锈环棱螺、大沼螺全湖区广泛分布,在西北湖区平均密度和平均生物量最高,中部湖区较低.河蚬与前二者分布相似,在中部湖区分布最低,平均密度和平均生物量仅分别为3.45ind/m2、7.06g/m2,在其它湖区则分布较高.淡水壳菜主要集中分布在北部湖区,其它湖区相对较少.
昆虫纲主要以摇蚊幼虫为主,全湖摇蚊幼虫平均密度为19.94ind/m2,其优势种为中国长足摇蚊、德永雕翅摇蚊和黄色羽摇蚊,平均密度为8.95ind/m2,占摇蚊幼虫总密度的44.87%.摇蚊幼虫空间分布上差异较大,其中中部湖区、南部湖区、北部湖区摇蚊幼虫种类较多,平均密度较高,分别为29.53,24.54, 22.24ind/m2,东部湖区发现种类较少,摇蚊幼虫平均密度为12.50ind/m²,西北部湖区仅发现中国长足摇蚊,平均密度为3.86ind/m2.除摇蚊幼虫外,昆虫纲其它种类较少,主要以杨氏桑嘎蜉为主,北部湖区和西北部湖区分布较高,平均密度分别为10.60和9.85ind/m2,而中部湖区和南部湖区未发现.

图5 鄱阳湖大型底栖动物各类群密度和生物量的空间格局
寡毛纲、多毛纲、蛭纲和甲壳纲在鄱阳湖大型底栖动物总密度中所占比例很低(9.95%).寡毛纲仅采集到苏氏尾鳃蚓和霍甫水丝蚓2种,全湖平均密度分别为2.39和2.68ind/m2,主要分布在西北部湖区、中部湖区和东部湖区,其他湖区密度较低.多毛纲主要分布在各湖区沿岸带部分样点中,其优势种为寡鳃齿吻沙蚕,主要分布在北部湖区和西北部湖区,其他湖区密度较低.蛭纲主要分布在中部湖区及北部湖区的沿岸带,东部湖区并未发现该类群.甲壳纲在全湖区出现率较低,优势种为秀丽白虾,北部湖区相对分布较多,其它湖区密度很低.
2.3 群落结构差异和多样性
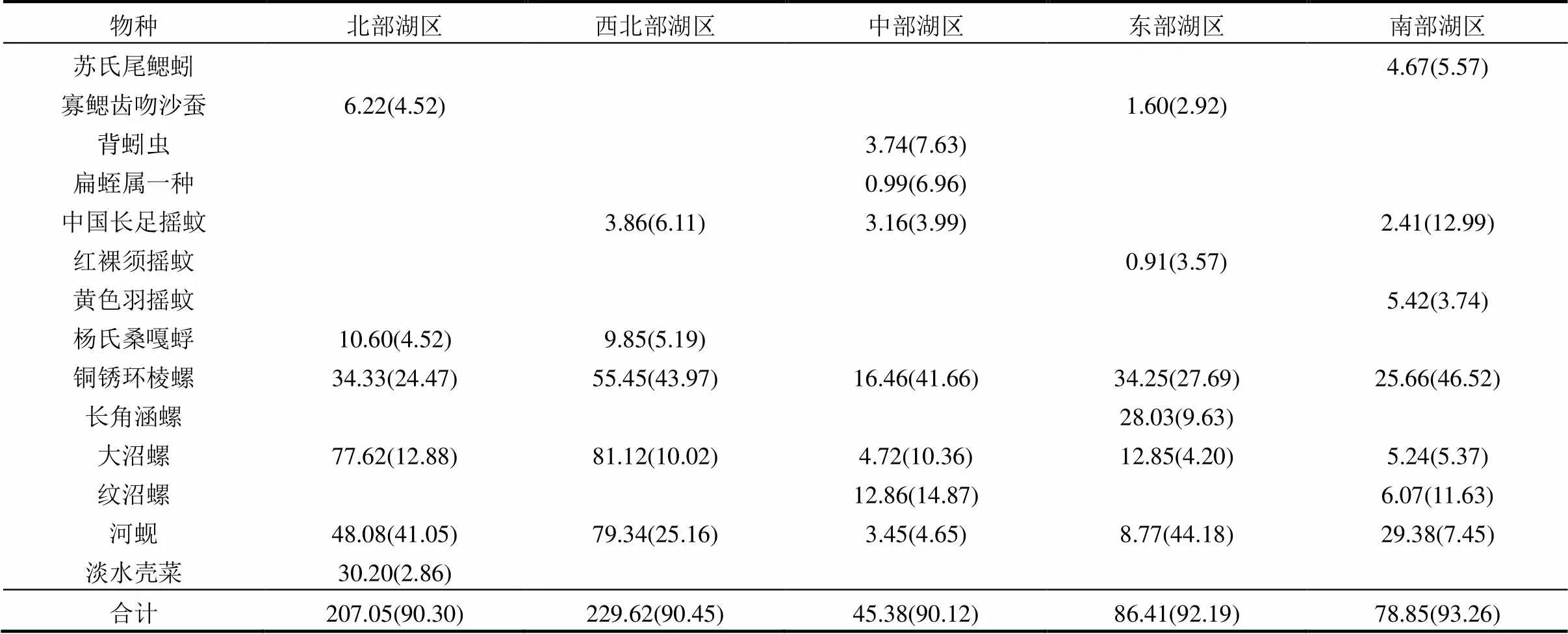
表3 不同湖区大型底栖动物特征种平均密度(ind/m²)及其对组内相似性贡献百分比(%)
ANOSIM检验结果表明各湖区之间的底栖动物群落存在显著差异(=0.002),各湖区的相似性百分比介于80.80%~89.62%之间(表3).北部湖区的特征种为河蚬、铜锈环棱螺、大沼螺、寡鳃齿吻沙蚕、杨氏桑嘎蜉、淡水壳菜,SIMPER分析表明这6个种类对组内相似性贡献率最大(累积达到90.30%).西北部湖区的特征种为铜锈环棱螺、河蚬、大沼螺、中国长足摇蚊、杨氏桑嘎蜉,其对组内相似性的累积贡献率为90.45%.中部湖区的特征种为铜锈环棱螺、纹沼螺、大沼螺、背蚓虫、扁蛭属一种、河蚬、中国长足摇蚊,其对组内相似性的累积贡献率为90.12%.东部湖区的特征种为河蚬、铜锈环棱螺、长角涵螺、大沼螺、红裸须摇蚊、寡鳃齿吻沙蚕,其对组内相似性的累积贡献率为92.19%.南部湖区的特征种为铜锈环棱螺、中国长足摇蚊、纹沼螺、河蚬、苏氏尾鳃蚓、大沼螺、黄色羽摇蚊,其对组内相似性的累积贡献率为93.26%.
4个多样性指数在不同湖区间均存在显著差异(<0.05,图6).Simpson指数全湖区平均值为0.43,北部湖区平均值最高(0.68),东部湖区平均值最低(0.32).Margalef指数全湖区平均值为1.03,北部湖区平均值最高(1.82),东部湖区平均值最低(0.68). Shannon-Wiener指数全湖区平均值为0.80,北部湖区平均值最高(1.39),东部湖区平均值最低(0.56). Pielou指数全湖区平均值为0.62,北部湖区平均值最高(0.88),西北部湖区平均值最低(0.50).总体而言,北部湖区相较于其它湖区的底栖动物多样性更高.
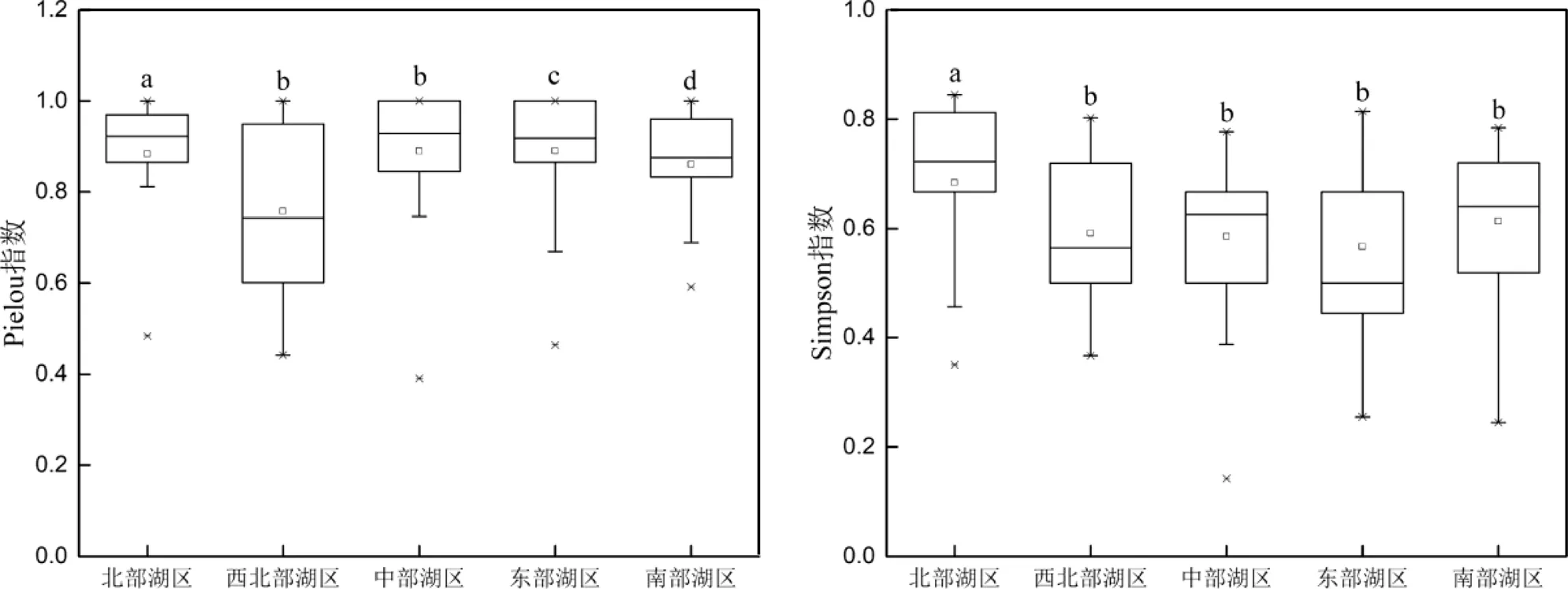
不同字母表示湖区间具有显著差异(<0.05)
2.4 关键环境影响因子识别
DCA分析结果表明第一轴的轴长5.24,因此选择单峰模型CCA排序分析.与沉积物参数作CCA分析筛选出2个环境因子,第一排序轴与LOI相关性较高,第二排序轴与砂(>63µm)相关性较高(图7),CCA分析的第1轴和第2轴的特征值分别为0.241和0.161,分别解释10.4%的物种数据方差变异和44.07%的物种-环境关系变异(表4).底栖动物密度与水体理化因子CCA分析最终筛选出5个环境因子.第1排序轴与水深、Chl-a、TP、DO相关性较高,第2排序轴与浊度相关性较高,CCA分析的第1轴和第2轴的特征值分别为0.318和0.293,分别解释11.2%的物种数据方差变异和33.95%的物种-环境关系变异.
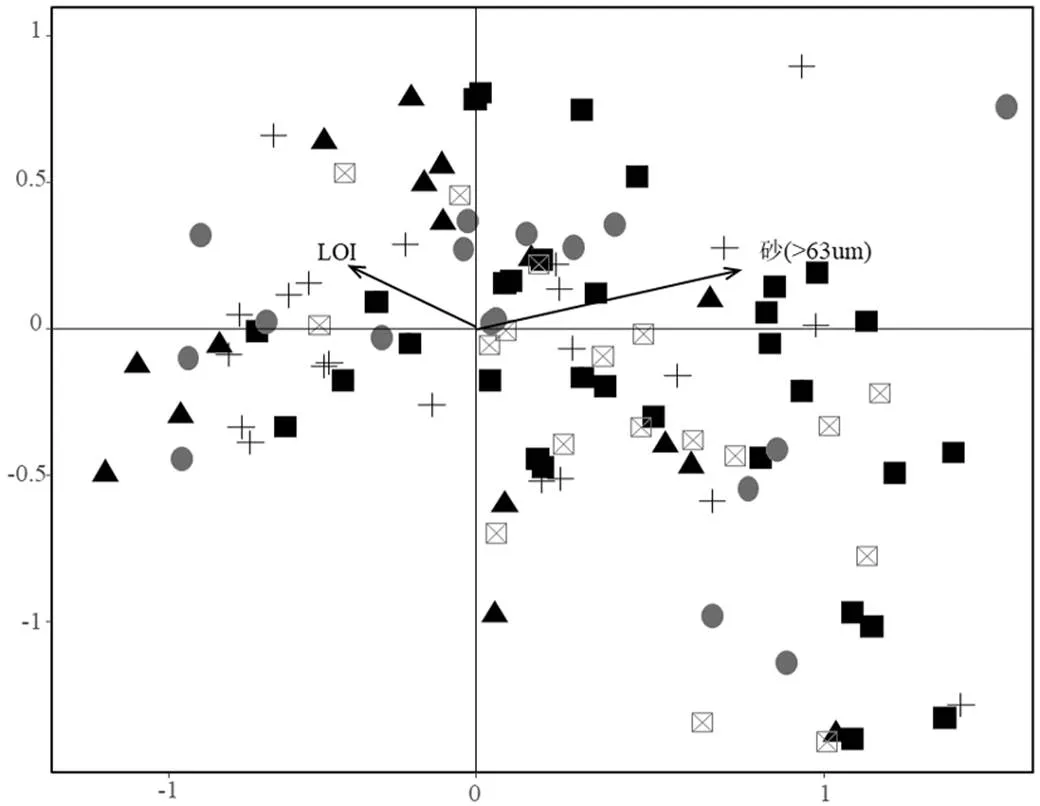
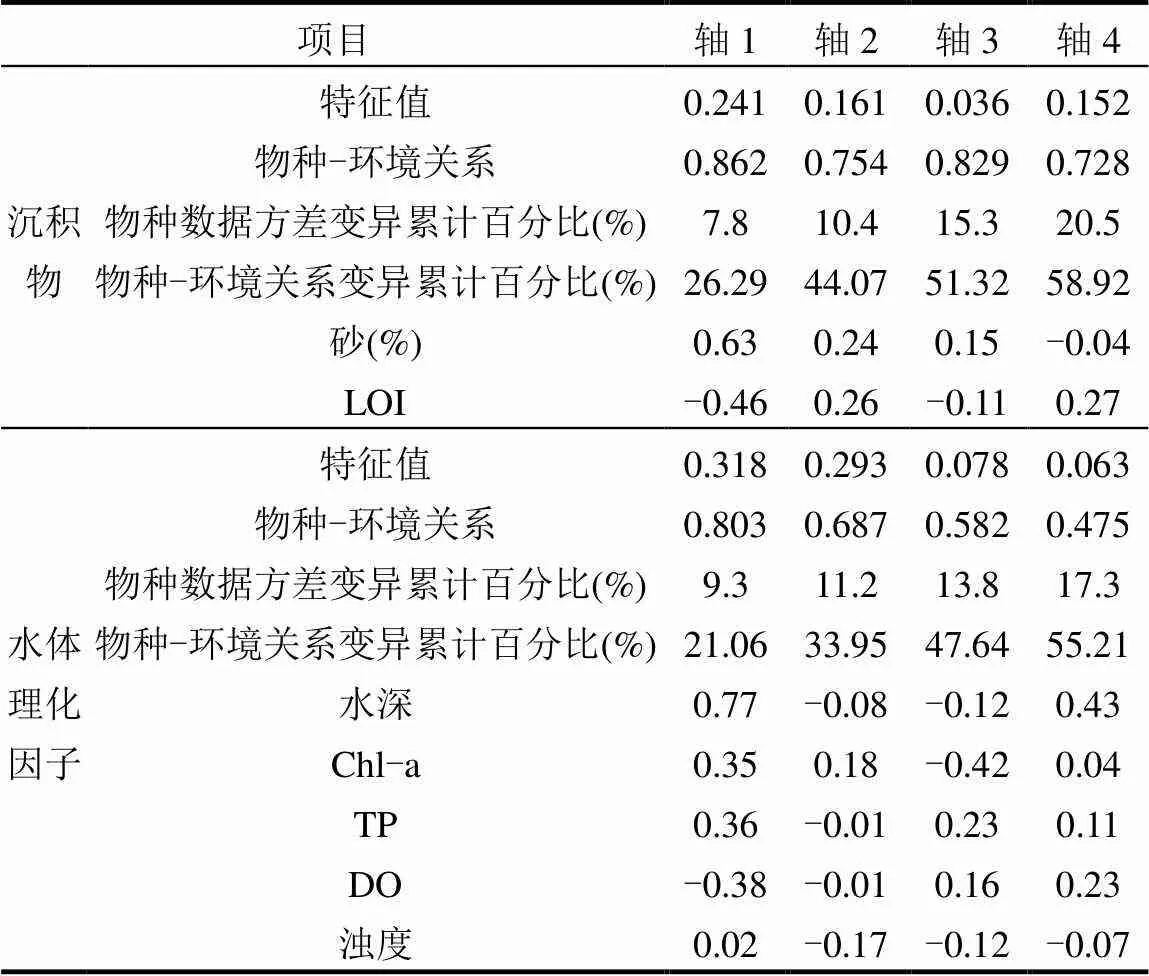
表4 底栖动物群落与环境因子的典范对应分析结果
3 讨论
3.1 底栖动物群落结构空间异质性及影响因素
不同的区域环境因素对底栖动物群落结构和现存量会产生显著影响.本文研究表明,底质类型、水深、溶解氧、浊度、总磷、叶绿素a是影响鄱阳湖大型底栖动物分布的主要影响因子.富营养化对湖泊底栖动物的影响前人已有充分的研究[20-21],此外,对于过水性湖泊而言,鄱阳湖平均换水周期约为30d[22],湖泊中的底栖藻类将会更容易被水流带走,因此对底栖动物而言,将可能产生食物限制.而CCA分析结果表明Chla、TP为驱动底栖动物群落结构的主要影响因子,这2个指标除了可以衡量水体的富营养水平外,也可以从一定程度上表征底栖动物食物量多少,这一现象在同为过水性湖泊洞庭湖的研究中也有报道[23].此外,LOI是表征沉积物中有机质含量的指标,也可以表征底栖动物食物数量和质量.由于LOI和Chla、TP均是驱动底栖动物群落结构的影响显著因子,可以推测食物资源可能主要影响着鄱阳湖底栖动物群落结构.鄱阳湖东部湖区底栖动物均匀度与多样性显著低于其它湖区,该区域底质粒径偏细,粒径范围较窄,底质异质性较低难以为多类底栖动物提供栖息场所.北部湖区与南部湖区底质粒径范围较广,生境多以淤泥+沙混合底质为主,为底栖动物提供了生存空间和庇护场所,适合软体动物栖息,同时也有利于寡毛类和摇蚊幼虫生存,从而表现出更高的多样性.底质粒径大小是底质类型的重要决定因素.一般而言,底栖动物多样性随底质粒径增大而发生显著变化,且一般不同类型底质中的优势类群不同[24].底质粒径大小与有机质和有机污染物的富集有关,粒径较小的底质与有机质更易结合,低有机质含量的底质无法支撑复杂的底栖动物食物网结构和较高生物量,造成湖泊底栖动物群落结构的改变和群落多样性的显著降低[25].不同类群底栖动物对底质类型的喜好差异较大,一般而言,粒径偏小的底质不利于滤食者的摄食,影响软体动物的栖息,如河蚬喜欢砂质淤泥底质,而颤蚓类和摇蚊幼虫喜好栖居于淤泥底质中,因此底质类型直接影响底栖动物的群落组成和多样性[26].一般认为,从沿岸带生境多样性较高,能够容纳更多的动物生存,因此底栖动物多样性会随着沿岸带到湖心逐渐降低.本研究中虽然水深也是驱动鄱阳湖底栖动物群落结构的主要影响因子,但结果表明,鄱阳湖深水区域相较于沿岸洲滩与湖湾浅水区域底栖动物丰富度较高,如软体动物主要分布在湖区内部深水区域,而沿岸区域分布较少.出现这种截然相反的情况是因为鄱阳湖不同季节水位波动较大,在干旱季节,沿岸带长期出露水面,从而导致其生境不稳定,抑制底栖动物生存.浊度和溶解氧能够直接影响底栖动物生存,尤其是双壳类,对浊度和溶解氧要求较高.
3.2 底栖动物群落结构历史演变
近年来,受人类活动、江湖关系逐渐改变等影响,鄱阳湖湖泊富营养化程度严重[27],底栖动物群落组成与结构也发生了显著变化.本次调查中,共采集底栖动物48种,平均密度和生物量为158.38ind/m2和173.76g/m2,优势种为河蚬、铜锈环棱螺和大沼螺.历史上对鄱阳湖底栖动物进行过多次调查(表5),与20世纪90年代相比,物种数、平均密度与生物量下降明显.另外大型底栖动物群落结构变化显著,优势种呈现小型化的趋势,从丰富的软体动物演变为小型软体动物河蚬和耐污能力较强的铜锈环棱螺,出现了蚌类资源衰退的现象.张屹等[28]对洞庭湖大型底栖动物群落演变的研究表明,近30a大型底栖动物的密度呈波动式下降趋势,相对较为耐污的种类成为优势种,这一演变特征与鄱阳湖较为相似.此外,由于本次调查在鄱阳湖布设了133个监测点,因此与历史研究相比,本次调查中沿岸带监测点位更多,而螺类在沿岸带分布较多,这也可能是本次调查中优势种多数为螺类的主要原因.

表5 鄱阳湖大型底栖动物的物种数、优势种、平均密度和平均生物量的年代变化
底栖动物群落结构的变化,与栖息环境变化和人为干扰程度的加剧密不可分.引起鄱阳湖底栖动物群落结构变化可能主要有以下方面原因:一方面是鄱阳湖存在大量采砂作业的现象,2001~2010年鄱阳湖采砂面积约为260.40km2,挖砂深度平均4.95m, 2005~2006年采砂量为2.36亿m3,占长江中下游地区消费量的40%[29-30].采砂造成的底质破坏以及水体浊度升高使底栖动物栖息地丧失和食源严重不足[31].刘振元等[32]对南洞庭湖软体动物多样性评估表明,采砂区物种组成最为单一,采砂是影响软体动物生存的重要制约因素.Meng等[33]对洞庭湖的研究表明采砂活动直接导致水深增加,透明度降低,沉积物粒径改变,软体动物类群丧失赖以生存的栖息地,进而导致湖泊底栖动物群落结构变得简单化.剧烈的采砂活动导致湖泊底质生境稳定性较低,部分底栖动物直接被清除或死亡.另一方面是鄱阳湖水位变化明显,研究表明,1990~2016年期间,长江干流水量减少导致鄱阳湖丰水期平均水位下降明显,而地形改变导致枯水期水位下降,水域面积低至500km2[34].2005年后鄱阳湖水位与之前相比降幅可达9.4%,枯水期提前且持续时间长,10月份水位异常偏低[35].高变幅的水位对底栖动物群落将产生变化,特别对湖泊沿岸带底栖动物产生显著影响[36]. Aroviita等[37]对芬兰23个湖泊的对比研究发现,沿岸带底栖动物在不同水位变幅情况下具有显著差异,物种丰富度随变幅的增大而降低.一般认为,水草是底栖动物分布的重要环境因子.大型沉水植物为底栖动物提供了更加多样的生境,利于腹足类、昆虫类等底栖动物生活、摄食和繁殖.茂盛水草的存在亦为底栖动物躲避鱼类及其它大型动物的掠食以及水流、风力的不利影响提供了避难场所.近30a来鄱阳湖长期持续低枯湖水位,水生植被生存空间被逐渐压缩,导致水生植被大幅缩减[38],可能也是鄱阳湖底栖动物群落结构变化的重要原因.
4 结论
4.1 本次鄱阳湖调查共采集大型底栖动物48种,平均密度为158.38ind/m2,平均生物量为173.76g/m2,底栖动物的主要优势种为河蚬、铜锈环棱螺、大沼螺,不同湖区的种类数、优势种均存在显著差异.
4.2 空间尺度上,水深、溶解氧、浊度、总磷、叶绿素a、烧失量和底质类型是鄱阳湖底栖动物群落结构的显著影响因子.
4.3 时间尺度上,与20世纪90年代相比鄱阳湖底栖动物的物种数呈下降的趋势,优势种从大型软体动物逐渐演变成小型软体动物,引起鄱阳湖底栖动物群落结构变化可能与采砂、水文情势变化、水生植被衰退等因素有关.
[1] 王备新,杨莲芳.大型底栖无脊椎动物水质快速生物评价的研究进展[J]. 南京农业大学学报, 2001,24(4):107-111.
Wang B X, Yang L F. Advances in rapid bio-assessment of water quality using benthic macroinvertebrates [J]. Journal of Nanjing Agricultural University, 2001,24(4):107-111.
[2] Covich A P, Palmer M A, Crowl T A, et al. The role of benthic invertebrate species in freshwater ecosystems: zoobenthic species influence energy flows and nutrient cycling [J].BioScience, 2004,54(8):767-775.
[3] 张 又,刘 凌,蔡永久,等.太湖流域河流及溪流大型底栖动物群落结构及影响因素[J]. 中国环境科学, 2015,35(5):1535-1546.
Zhang Y, Liu L, Cai Y J, et al. Benthic macroinvertebrate community structure in rivers and streams of Lake Taihu Basin and environmental determinants [J]. China Environmental Science, 2015,35(5):1535-1546.
[4] 金小伟,王业耀,王备新,等.我国流域水生态完整性评价方法构建[J]. 中国环境监测, 2017,33(1):75-81.
Jin X W, Wang Y Y, Wang B X, et al. Methods development for monitoring and assessment of ecological integrity of surface waters in China [J]. Environmental Monitoring in China, 2017,33(1):75-81.
[5] Li B, Yang G S, Wan R R, et al. Comparison of random forests and other statistical methods for the prediction of lake water level: a case study of the Poyang Lake in China [J]. Hydrology Research, 2016, 47(S1):69-83.
[6] Zhang Z X, Chen X, Xu C Y, et al. Examining the influence of river–lake interaction on the drought and water resources in the Poyang Lake basin [J]. Journal of Hydrology, 2015,522:510-521.
[7] 张 琍,陈晓玲,张 媛,等.水文地貌分区下鄱阳湖丰水期水质空间差异及影响机制[J]. 中国环境科学, 2014,34(10):2637-2645.
Zhang L, Chen X L, Zhang Y, et al. Spatial distribution of water quality and its impacting factor in the wet season of Poyang Lake using the hydro-geomorphological partitions [J]. China Environmental Science, 2014,34(10):2637-2645.
[8] 王伟莉,闫振广,何 丽,等.五种底栖动物对优控污染物的敏感性评价 [J]. 中国环境科学, 2013,33(10):1856-1862.
Wang W L, Yan Z G, He L, et al. Sensitivity evaluation of five zoobenthos to priority pollutants [J]. China Environmental Science, 2013,33(10):1856-1862.
[9] 谢钦铭,李 云,熊国根.鄱阳湖底栖动物生态研究及其底层鱼产力的估算[J]. 江西科学, 1995,13(3):161-170.
Xie Q M, Li Y, Xiong G G. The ecological study of macrozoobenthic and the estimation of demersal fish productivity in Poyang Lake [J]. Jiangxi Science, 1995,13(3):161-170.
[10] Wang H Z, Xu Q Q, Cui Y D, et al. Macrozoobenthic community of Poyang Lake, the largest freshwater lake of China, in the Yangtze floodplain. [J]. Limnology, 2007,8(1):65-71.
[11] 欧阳珊,詹 诚,陈堂华,等.鄱阳湖大型底栖动物物种多样性及资源现状评价[J]. 南昌大学学报(工科版), 2009,31(1):9-13.
Ou Y S, Zhan C, Chen T H, et al. Species diversity and resource assessment of macrozoobenthos in Poyang Lake [J]. Journal of Nanchang University(Engineering & Technology), 2009,31(1):9-13.
[12] Cai Y J, Lu Y J, Wu Z S, et al. Community structure and decadal changes in macrozoobenthic assemblages in Lake Poyang, the largest freshwater lake in China [J]. Knowledge and Management of Aquatic Ecosystems, 2014,414:1-18.
[13] 杨桂山,马荣华,张 路,等.中国湖泊现状及面临的重大问题与保护策略[J]. 湖泊科学, 2010,22(6):799-810.
Yang G S, Ma R H, Zhang L, et al. Lake status, major problems and protection strategy in China [J]. Journal of Lake Sciences, 2010,22(6): 799-810.
[14] 钱奎梅,刘宝贵,陈宇炜.鄱阳湖浮游植物功能群的长期变化特征(2009~2016年) [J]. 湖泊科学, 2019,31(4):1035-1044.
Qian K M, Liu B G, Chen Y W. Long term dynamics of phytoplankton functional groups in Lake Poyang during 2009~2016 [J]. Journal of Lake Sciences, 2019,31(4):1035-1044.
[15] 刘月英,张文珍,王跃先,等.中国经济动物志-淡水软体动物[M]. 北京:科学出版社, 1979.
Liu Y Y, Zhang W Z, Wang Y X, et al. Economic fauna of China (freshwater mollusk) [M]. Beijing:Science Press, 1979.
[16] Morse J C, Yang L F, Tian L X. Aquatic insects of China useful for monitoring water quality [M]. Nanjing: Hohai University Press, 1994.
[17] 王俊才,王新华.中国北方摇蚊幼虫[M]. 北京:中国言实出版社, 2011.
Wang J C, Wang X H. Northern China Chironomus larva [M]. Beijing:Yanshi Press in China, 2011.
[18] 国家环境保护总局《水和废水监测分析方法》编委会.水和废水监测分析方法[M]. 4版.北京:中国环境科学出版社, 2002.
“Water and wastewater monitoring and analysis method”editorial board of State Environmental Protection Administration of China. Monitoring and analysis methods of water and wastewater [M]. fourth edition.Beijing:China Environmental Science Press, 2002.
[19] 韩 洁,张志南,于子山.渤海中、南部大型底栖动物的群落结构[J]. 生态学报, 2004,24(3):531-537.
Han J, Zhang Z N, Yu Z S. Macrobethic community structure in the southern and central Bohai Sea, China [J]. Acta Ecologica Sinica, 2004,24(3):531-537.
[20] 龚志军,谢 平,唐汇涓,等.水体富营养化对大型底栖动物群落结构及多样性的影响[J]. 水生生物学报, 2001,25(3):210-216.
Gong Z J, Xie P, Tang H J, et al. The influence of eutrophycation upon community structure and biodiversity of macrozoobenthos [J]. Acta Hydrobiologica Sinica, 2001,25(3):210-216.
[21] 蔡永久,龚志军,秦伯强.太湖大型底栖动物群落结构及多样性[J]. 生物多样性, 2010,18(1):50-59.
Cai Y J, Gong Z J, Qin B Q. Community structure and diversity of macrozoobenthos in Lake Taihu, a large shallow eutrophic lake in China [J]. Biodiversity Science, 2010,18(1):50-59.
[22] 李云良,姚 静,李梦凡,等.鄱阳湖换水周期与示踪剂传输时间特征的数值模拟[J]. 湖泊科学, 2017,29(1):32-42.
Li Y L, Yao J, Li M F, et al. Numerical simulations of residence time and tracer travel time in Lake Poyang [J]. Journal of Lake Sciences, 2017,29(1):32-42.
[23] 王小毛,欧伏平,王丑明,等.洞庭湖底栖动物长期演变特征及影响因素分析[J]. 农业环境科学学报, 2016,35(2):336-345.
Wang X M, Ou F P, Wang C M, et al. Long-term evolution and influencing factors of macrozoobenthos in Dongting Lake [J]. Journal of Agro-Environment Science, 2016,35(2):336-345.
[24] 段学花,王兆印,田世民.河床底质对大型底栖动物多样性影响的野外试验[J]. 清华大学学报(自然科学版), 2007,47(9):1553-1556.
Duan X H, Wang Z Y, Tian S M. Field experiment on the effect of streambed substrate on macroinvertebrate diversity [J]. Journal of Tsinghua University(Science and Technology), 2007,47(9):1553-1556.
[25] Thompson B, Lowe S. Assessment of macrobenthos response to sediment contamination in the San Francisco Estuary,California,USA [J]. Environmental Toxicology and Chemistry, 2004,23(9):2178-2187.
[26] 邹 伟,李太民,刘 利,等.苏北骆马湖大型底栖动物群落结构及水质评价[J]. 湖泊科学, 2017,29(5):1177-1187.
Zou W, Li T M, Liu L, et al. Macrozoobenthic community structure and water quality assessment of Lake Luoma, Jiangsu Province,China [J]. Journal of Lake Sciences, 2017,29(5):1177-1187.
[27] 胡春华,周文斌,王毛兰,等.鄱阳湖氮磷营养盐变化特征及潜在性富营养化评价[J]. 湖泊科学, 2010,22(5):723-728.
Hu C H, Zhou W B, Wang M L, et al. norganic nitrogen and phosphate and potential eutrophication assessment in Lake Poyang [J]. Journal of Lake Sciences, 2010,22(5):723-728.
[28] 张 屹,王丑明,石慧华,等.洞庭湖近30年大型底栖动物的群落演变[J]. 生态环境学报, 2015,24(8):1348-1353.
Zhang Y, Wang C M, Shi H H, et al. Macrobenthic community succession during last thirty years in Dongting Lake [J]. Ecology and Environmental Sciences, 2015,24(8):1348-1353.
[29] 江 丰,齐述华,廖富强,等.2001~2010年鄱阳湖采砂规模及其水文泥沙效应[J]. 地理学报, 2015,70(5):837-845.
Jiang F, Qi S H, Liao F Q, et al. Hydrological and sediment effects from sand mining in Poyang Lake during 2001~2010 [J]. Acta Geographica Sinica, 2015,70(5):837-845.
[30] Lai X J, David S, Claire H, et al. Sand mining and increasing Poyang Lake’s discharge ability: A reassessment of causes for lake decline in China [J]. Journal of Hydrology, 2014,519:1698-1706.
[31] Zou W, Tolonen K T, Zhu G W, et al. Catastrophic effects of sand mining on macroinvertebrates in a large shallow lake with implications for management [J]. Science of the Total Environment, 2019,695: 133706.DOI:10.1016/j.scitotenv.2019.133706.
[32] 刘振元,孟星亮,李正飞,等.南洞庭湖区软体动物物种多样性评估及保护对策[J]. 生物多样性, 2020,28(2):155-165.
Liu Z Y, Meng X L, Li F F, et al. Diversity assessment and protection strategies for the mollusk community in the southern Dongting Lake [J]. Biodiversity Science, 2020,28(2):155-165.
[33] Meng X L, Jiang X M, Li Z F,et al.Responses of macroinvertebrates and local environment to short-term commercial sand dredging practices in a flood-plain lake [J]. Science of the Total Environment, 2018,631-632:1350-1359.DOI:10.1016/j.scitotenv.2018.03.086.
[34] 孙芳蒂,马荣华.鄱阳湖水文特征动态变化遥感监测[J]. 地理学报, 2020,75(3):544-557.
Sun F D, Ma R H. Hydrologic changes of Poyang Lake based on radar altimeter and optical sensor [J]. Acta Geographica Sinica, 2020,75(3): 544-557.
[35] 邓志民,张 翔,肖 洋,等.鄱阳湖水位演变及其影响因子分析[J]. 武汉大学学报(工学版), 2015,48(5):615-621.
Deng Z M, Zhang X, Xiao Y, et al. Study of evolution of water level in Poyang Lake and impact factors [J]. Engineering Journal of Wuhan University, 2015,48(5):615-621.
[36] 彭建华,郑志伟,邹 曦,等.汉丰湖底栖动物与环境因子间相关性初步研究[J]. 环境影响评价, 2015,37(5):63-68.
Peng J H, Zheng Z W, Zou X, et al. Study of correlation between benthos animals and environmental factors in Hanfeng Lake [J]. Environmental Impact Assessment, 2015,37(5):63-68.
[37] Aroviita J, Hämäläinen H. The impact of water-level regulation on littoral macroinvertebrate assemblages in boreal lakes [J]. Hydrobiologia, 2008,613(1):45-56.
[38] 胡振鹏,林玉茹.鄱阳湖水生植被30年演变及其驱动因素分析[J]. 长江流域资源与环境, 2019,28(8):1947-1955.
Hu Z P, Lin Y R. Analysis of evolution process and driving factors for aquatic vegetations of Poyang Lake in 30years [J]. Resources and Environment in the Yangtze Basin, 2019,28(8):1947-1955.
Characteristics and driving factors of spatiotemporal succession of macrozoobenthos in Poyang Lake.
ZOU Liang-hua1,2, ZOU Wei2, ZHANG Qing-ji2,3, LI Ying2,4, GONG Zhi-jun2, ZHANG Yan-jie1, LU Shun-bao1*, CAI Yong-jiu2**
(1.College of Life Sciences, Jiangxi Normal University, Nanchang 330027, China;2.Key Laboratory of Watershed Geographic Sciences, Nanjing Institute of Geography & Limnology, Chinese Academy of Sciences, Nanjing 210008, China;3. School of Geography & Ocean Science, Nanjing University , Nanjing 210023, China;4.School of Geography science, Southwest University, Chongqing 400715, China)., 2021,41(6):2881~2892
Macrozoobenthos community and environmental variables were investigated at 133 sites in Poyang Lake in July 2019. The relationships between macrozoobenthic community structure and environmental factors were examined. A total of 48 species belonging to 7 classes, 16 orders, 22 families and 38 genera were recorded. The average density and biomass of macrozoobenthos were 158.38 ind/m² and 173.76g/m², respectively, mainly dominated by,and. In terms of spatial patterns, the whole lake was divided into five areas, including northern, northwest, central, eastern and southern lake areas. One-way ANOVA analysis detected significant differences of physicochemical variables among the five lake areas, while no significant differences in sediment parameters. The community structure of benthic animals among the five lake areas also varied significantly as indicated by one-way analysis of similarity (ANOSIM). Regarding to biodiversity indexes, significantly higher values in northern lake were found than those in other lake areas. Canonical correspondence analyses (CCA) showed that water depth, dissolved oxygen, turbidity, total phosphorus, chlorophyll a, loss on ignition, and sediment type were significantly related to the community structure of macrozoobenthos. In the long term, the biodiversity of macrozoobenthos in Poyang Lake has declined significantly compared with those in their historical condition, with the dominated species changed from large molluscs to small molluscs and insects. Sand mining, changes in hydrological conditions and decline of aquatic vegetation were the main environmental factors regulating the spatiotemporal succession of the macrozoobenthic community structure in Poyang Lake.
Poyang Lake;macrozoobenthos;community structure;biodiversity;environmental factors
X171
A
1000-6923(2021)06-2881-12
2020-09-20
中国科学院战略性先导科技专项(A类)(XDA23040203);国家自然科学基金资助项目(32071572,31660072,31760136);江西省自然科学基金资助项目(20201BAB204237);江西省教育厅基金(GJJ160312, GJJ170198)
* 责任作者, 教授, luxunbao8012@126.com; ** 副研究员, caiyj@niglas.ac.cn
邹亮华(1994-),男,江西赣州人,硕士研究生,主要从事湿地生态学研究.
猜你喜欢
水生生物学报(2022年6期)2022-07-08 09:31:56
海洋通报(2022年2期)2022-06-30 06:06:28
当代水产(2021年8期)2021-11-04 08:49:06
海洋通报(2021年1期)2021-07-23 01:55:24
云南畜牧兽医(2021年1期)2021-02-24 04:37:28
意林·少年版(2019年1期)2019-01-16 20:01:10
海峡旅游(2018年4期)2018-06-01 11:20:00
江西建材(2018年4期)2018-04-10 12:37:24
海洋渔业(2017年5期)2017-11-07 02:34:58
天津师范大学学报(自然科学版)(2016年6期)2016-12-14 08:23:08
