
葡萄树基活性炭的制备及其CO2吸附特性
2021-07-17 02:26:12吐尔逊吐尔洪苏比努尔吾麦尔江阿不都热依木卡德尔赛达尔帕尔哈提
环境科学研究 2021年7期
吐尔逊·吐尔洪, 苏比努尔·吾麦尔江, 阿不都热依木·卡德尔, 赛达尔·帕尔哈提
新疆农业大学, 新疆 乌鲁木齐 830052
CO2是主要的温室气体之一,为实现可持续发展需要为工业生产提供廉价易得的CO2捕集材料. 目前,膜分离、吸收和吸附等技术广泛应用于CO2捕获脱除领域,其中膜分离技术的制备工艺复杂,成本较高;吸收方法的捕获量较大,但是能耗及对设备的腐蚀性也较大,且吸收剂再生困难;吸附方法采用的固体吸附剂廉价易得,再生性好,是目前主要研究热点之一. 常见的CO2固体吸附材料有金属氧化物、碳材料、分子筛、金属-有机框架(MOFs)材料等,因吸附材料具有碱性位点较高、再生容易、腐蚀性较小等特点而被广泛应用于CO2的吸附脱除领域[1]. 碳基吸附材料中,活性炭因具有原料来源广、制备工艺简单、成本低、吸附能力强、化学性质稳定、再生容易等特点,成为一种经济环保的CO2吸附剂[2],在碳捕集、碳封存和其他环保领域有较大应用前景.
活性炭的吸附固碳技术是公认的解决气候变化问题的措施之一,因此环保领域对功能型活性炭的需求日益增加[3-4]. 活性炭是由木质和石油焦等含碳生物质原料经不同高温条件下热解、活化和各种改性加工制备而成的,因有发达的孔隙结构、较大的比表面积和丰富的表面化学基团,故具有较强的吸附能力. 活性炭的活化方法有物理活化法、化学活化法、物理化学活化法、微波化学活化法等. 常见的化学活化试剂有磷酸、氯化锌、氢氧化钾、氢氧化钠、硫酸、碳酸钾、多聚磷酸和磷酸酯等. 物理活化法又称气体活化法,是将已炭化的生物质原料在600~1 200 ℃下与水蒸气、CO2、CO或空气等气体反应,从而在生物炭表面扩大孔隙的一种方法[5]. 因物理法具有工艺流程简单、CO2和水蒸气等活化剂易得、制备过程对环境污染较小、产品比表面积大等特性,世界范围内70%以上的活性炭采用物理法生产[4]. CO2活化法能生产孔径小于1 nm的微孔活性炭,适合于无机气体的吸附分离. 目前,利用活性炭吸附去除CO2方面的应用研究较多. 在国内外研究中活性炭吸附剂的原料主要集中在玉米芯[6]、椰壳[7]、蔗渣[8]、棉花秸秆[9]、兰炭粉[10]、壳聚糖[11]、石油焦[12]、原料煤[13]、竹子、松子壳和瓜子壳[14]等材料上. 葡萄种植业每年产生大量的葡萄茎、葡萄榨渣、葡萄籽和葡萄皮等废物,其中葡萄茎是以木质纤维素为主,主要用于提取纤维素、半纤维素、酚类抗氧化和发酵性糖等[15]. 以葡萄生产为主导产业的新疆维吾尔自治区吐鲁番地区,每年在修剪老化葡萄树枝条的过程中也产生大量葡萄树枝废物. 葡萄树枝(葡萄茎)自身有一定多孔结构和吸附能力,故用于水中部分金属离子、有机污染物和多种气态污染物的吸附去除领域[15-18]. 此外,不同生物质固有的表面化学性质不同,被CO2活化后形成不同的活性结构,新形成的活性结构决定活性炭的吸附特性. 最近国外已有以葡萄籽[19]、葡萄穗[20]、葡萄蔓[21]、葡萄藤落皮层[22]和葡萄榨渣[23]为原料制备活性炭的研究,涉及ZnCl2、KOH、H3PO4、水蒸气和小流量CO2(0.1 mL/min)活化方法,以及表面特性的表征和镉吸附等[18,24-25]方面,而以葡萄树为原料制备活性炭及其碳捕集方面的应用研究较少. 国内葡萄树基活性炭的制备和应用方面的研究也较为鲜见. 活性炭的性能因原料自身性质的不同而存在较大差异. 不同生物质及其活性炭产物具有独特的孔隙结构和表面官能团类型. 因此,用CO2活化制备的葡萄树基活性炭的结构、化学特性及其吸附CO2的机制尚不明确. 此外,利用葡萄树枝制备活性炭可以高效利用葡萄产业的副产品、提高经济效益和促进循环经济的发展. 所以,为了发挥葡萄树生物质在吸附剂领域上的潜力,明确葡萄树基活性炭的结构特性、吸附性能以及为葡萄树枝寻找一种更加有效地资源化途径十分必要.
该研究以新疆维吾尔自治区吐鲁番地区常见的枯竭的葡萄树枝和树皮为材料,用CO2活化法制备了活性炭,用比表面积和热重分析等方法进行了表征,并对其吸附CO2的特性进行了分析,以期为葡萄树废树枝和树皮的资源化利用、碳捕集和节能减排等提供新材料,为CO2的吸附处理提供参考数据.
1 材料与方法
1.1 活性炭制备
将自然风干的干净葡萄树皮和树枝条(直径为1.5~2.0 cm)粉碎至1~2 mm颗粒,过40目(0.5 mm)筛后烘干,称取一定质量的粉末样品放入石英坩埚,然后置于横式管式炉的石英管内,通入N2(流量200 mL/min)吹扫5 min以排除管内氧气. 开启管式炉,从25 ℃起以10 ℃/min加热至预定碳化温度500 ℃,在500 ℃碳化60 min后,以10 ℃/min继续升温30 min,升至800 ℃时通入99.99% CO2(1 000 mL/min),活化60 min,全过程保持N2保护,自然冷却后获得葡萄树皮和树枝基活性炭.
1.2 活性炭的孔结构表征
采用Micromeritics ASAP-2020(美国麦克公司,美国)全自动比表面及孔隙度分析仪测定吸附材料的比表面积、孔径分布和孔体积等,分析原理基于静态容量法等温吸附原理. 测量方法与条件:测试前将样品置于样品管中,在300 ℃下进行加热抽真空脱气预处理4 h; 77 K下,在0~1的相对压力(P/P0)范围内,用高纯N2静态容量法测定吸附脱附容量. 根据不同相对压力下N2吸附量的不同,可以得到N2吸附脱附等温线. 根据N2吸附等温线用BET方法计算比表面积,孔径分布采用二维非定域密度函数理论(2D-NLDFT)进行计算,2D-NLDFT的计算采用了SAIEUS软件. 微孔总孔体积采用t-Plot法获得[26].
1.3 CO2吸附试验和热重特性
使用热重分析仪(Metro-Toledo TGA/DSC 1/1100 SF,瑞士梅特勒-托利多公司)记录了被测样品在30 ℃ 下吸附CO2过程中的质量变化,最终通过质量变化计算得到材料的CO2吸附量. 具体操作步骤:称取微量样品(10~20 mg左右)装载到坩埚内后放入热重分析仪,待流量计和天平稳定,测量室被抽真空后通入CO2,流量控制为60 mL/min,吸附试验进行60 min;继续切换N2(100 mL/min),以10 ℃/min升温至1 000 ℃,测试被吸附CO2和活性炭样品的热稳定性; 然后按同样速率降温至30 ℃,记录冷却过程中的质量变化数据;同时用原始活性炭作对照. 分别用吸附量公式[27]计算吸附质量占样品质量的百分比以及单位质量活性炭所吸附的物质的量,单位分别为%和mmol/g.
2 结果与讨论
2.1 孔隙特性
葡萄树枝基活性炭和葡萄树皮基活性炭的比表面积及孔径参数如表1所示. 由表1可见,以葡萄树皮为原料的活性炭的比表面积和孔体积比以葡萄树枝为原料的活性炭小,葡萄树枝基活性炭的比表面积为418.89 m2/g,比葡萄树皮基活性炭的比表面积大20%左右,且孔体积大1倍. 葡萄树皮基活性炭的孔径大于葡萄树枝基活性炭,说明葡萄树皮基活性炭的微孔结构少于葡萄树枝基活性炭,微孔结构发达的活性炭有利于吸附CO2等粒径较小的无机气体. 高温CO2活化法可以有效提高生物质炭的表面积,扩大其孔隙结构[28]. Ruiz-Fernández等[21]以葡萄蔓为原料,在750 ℃下用低流量CO2(10 mL/min)活化1 h得到的活性炭比表面积为5.0 m2/g,微孔体积也较小. Ozdemir等[24]用低流量CO2(0.1 L/min)活化法在700 ℃下活化120 min,获得的葡萄茎基活性炭的比表面积只有83.7 m2/g. 所以,笔者研究中两种活性炭比表面积的增大是CO2活化剂流量增加所致. Deiana等[18]在700 ℃下用水蒸气活化165 min制备了葡萄茎基活性炭,用酸去除部分灰分元素后,将比表面积和微孔体积从原来的412 m2/g和0.18 cm3/g分别提至906 m2/g和0.34 cm3/g. 采用H3PO4活化60 min制备的活性炭比表面积和微孔体积分别为1 676 m2/g和0.77 cm3/g,但这两种活性炭的DFT孔径集中在以0.6和1.5 nm为中心的2个区间,而孔径在1.5 nm附近的不利于吸附CO2的微孔体积在总孔体积中的占比近50%. 笔者研究中葡萄树枝基活性炭的孔结构特性非常接近于水蒸气活化葡萄茎基活性炭的特性. Shahraki等[25]用KOH活化法(活化温度400 ℃,活化时间120 min)制备的用于镉吸附的葡萄树茎基活性炭具有微孔-中孔结构,其比表面积为834 m2/g,微孔和中孔的孔径分别为1.73、2.41 nm,孔体积分别为0.302、0.228 cm3/g,比表面积和微孔体积是笔者研究中葡萄树枝基活性炭的2倍,但孔径较大也不利于吸附CO2,所以在保留小微孔的前提下,增加比表面积的制备和活化方法有待进一步探索.

表1 葡萄树基活性炭的孔隙结构特征及其CO2吸附量
由图1可见,两种活性炭等温线形状有明显区别. 葡萄树枝基活性炭的等温线属于I型等温线(国际纯粹与应用化学联合会IUPAC的分类),即在较低相对压力区域吸附容量呈缓慢垂直上升趋势,升至一个极限值后达到准平衡状态,这个特性符合I型吸附等温线,表示含有大量微孔结构. 所以,由此可判断葡萄树枝基活性炭为微孔结构. 葡萄树皮基活性炭的等温线呈S型〔见图1(a)〕,但中间段的迟滞环对应多孔吸附剂上出现的毛细凝聚现象,所以属于等温线的Ⅳ型H3类曲线,该类型等温线表示含有平板狭缝结构、裂缝和楔形结构的吸附剂上的物理吸附过程,即H3型迟滞环出现在含有片状颗粒或有狭缝状孔隙的吸附剂上,这说明由葡萄树皮制备的活性炭具有楔形孔隙结构. 虽然与Ⅱ型等温线类似,但在中等相对压力下毛细凝聚的发生使Ⅳ型等温线较Ⅱ型等温线上升得更快. 在等温线的中段,吸附等温线和脱附等温线明显不重合,脱附曲线高于吸附曲线,这种吸附滞后现象的出现对应Ⅳ型等温线的中孔结构,即葡萄树皮基活性炭也含有中孔结构. 吸脱附等温线在相对压力大于0.4的区域出现的“滞回”对应了中孔和大孔结构[29]. 中孔被毛细凝聚填满后,因吸附质分子相互作用强,继续吸附形成多分子层,吸附等温线继续上升. 在较高相对压力区域也没有表现出任何吸附平衡状态,进一步说明该材料表面结构上存在中孔结构. 因此,葡萄树皮基活性炭为微孔-中孔结构. 微孔在活性炭吸附污染物的过程中提供了大多数的吸附位点,中孔结构决定了吸附速度,既可以作为吸附质粒子进入微孔内的通道,又可以作为直接吸附粒径稍大且无法被微孔吸附的位点.
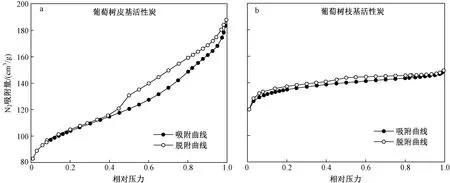
图1 葡萄树枝基活性炭和葡萄树皮基活性炭的N2吸附等温线
二维非定域密度函数理论(2D-NLDFT)适合于分析微孔和微孔-中孔活性炭的孔径[30],两种活性炭的2D-NLDFT孔径分布结果如图2所示.

图2 葡萄树枝基活性炭和葡萄树皮基活性炭的2D-NLDFT孔径分布
由图1(a)可见:葡萄树皮基活性炭的等温线在相对压力为0时N2吸附量为82.92 cm3/g,对应微孔吸附;相对压力升至1.0时,连续且快速增加的N2吸附量对应孔径为2.0~30 nm的中孔. 由图2可见:2D-NLDFT模型较好地拟合了葡萄树皮基活性炭的微孔-中孔分布,微孔孔径主要集中在0.36~1.9 nm;中孔孔径集中在3.0~7.0 nm,但其体积远少于微孔体积. 葡萄树枝基活性炭孔径主要集中在0.36~1.6 nm,孔径为0.66 nm的微孔体积最大,无中孔出现.
综上,两种活性炭不含大孔,微孔数量较多,葡萄树皮基活性炭为微孔-中孔结构,葡萄树枝基活性炭主要为微孔结构.
2.2 CO2吸附特性
吸附压力、温度、CO2初始浓度以及气体速率都对活性炭吸附CO2的性能产生影响. Yaumi等[31]发现,常压下椰壳基活性炭在30 ℃时CO2吸附量最大,当温度升高时CO2吸附量明显降低. Mohanad等[32]研究显示,花生壳基活性炭在0、25和50 ℃下CO2吸附量分别为5.3、4.2和0.90 mmol/g. 活性炭对CO2的吸附是放热过程,所以温度越高吸附量越低. 该研究选择了30 ℃和常压为吸附试验条件,在30 ℃下葡萄树基活性炭对纯CO2的吸附结果如图3和表1所示.
由图3可见,两种活性炭与CO2接触时,很快发生物理吸附. 葡萄树皮和葡萄树枝基活性炭达到吸附量最高值分别需要75和95 s,对应的最高吸附量分别为0.598和1.326 mmol/g. 被两种活性炭吸附的CO2分子在孔隙中的热稳定性有明显差异. 葡萄树皮基活性炭的吸附量达到最高值后开始快速释放CO2,达到平衡时吸附量减少了一半以上,而葡萄树枝基活性炭平衡时吸附量降低的比例较小. 葡萄树枝基活性炭的CO2平衡吸附容量约为葡萄树皮基活性炭的4倍.
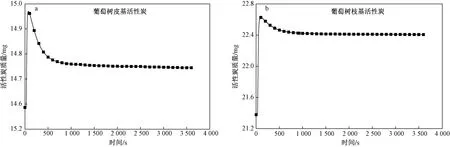
图3 葡萄树枝基活性炭和葡萄树皮基活性炭的CO2吸附曲线
活性炭吸附剂对CO2的吸附特性由比表面积、孔径、孔体积和表面官能团等因素决定. 一般来说,孔隙大小与吸附质分子大小接近时容易发生孔隙填充,而不容易发生毛细管凝聚,二者越接近吸附性能越强,CO2分子直径为0.33 nm,因此孔径越接近0.33 nm,吸附就越强. 袁翠翠[33]在920 ℃制备的煤基活性炭的比表面积和总孔体积分别为 1 071 m2/g和 0.447 0 mL/g,CO2吸附量高达59.44 mL/g (2.651 mmol/g),吸附过程中0.93 nm以内的孔隙对CO2吸附起关键作用. Ismail等[23]研究显示,用不同比例KOH(800 ℃)活化制备的葡萄榨渣基活性炭的比表面积范围为1 640~2 473 m2/g(孔径在0.52~0.76 nm),KOH:碳(质量比)越高,比表面积和孔径越大,但在常温常压(25 ℃,100 kPa)下CO2吸附量最高(3.9 mmol/g)的活性炭的比表面积、孔径分别为 1 727 m2/g、0.52 nm,并不是比表面积最大的活性炭. 孔径是活性炭CO2吸附效果的重要影响因子之一,根据IUPAC分类,微孔还可以细分为小微孔(<0.7 nm)和大微孔(0.7~1.0 nm)[34],孔径小于0.7 nm的孔体积和CO2吸附量之间的相关系数大于0.90[35]. 综上,在CO2分子被葡萄树枝基活性炭吸附的过程中,孔径接近0.66 nm的微孔起主要作用. 表2为用CO2物理活化法制备的活性炭在常温常压下的CO2吸附量,结果显示比表面积大小和CO2吸附量之间没有明显的线性关系,葡萄蔓基活性炭的比表面积较小,微孔体积占比只有51.7%,但孔径最小的葡萄蔓基活性炭的CO2吸附量最大. 综上,活性炭同时具备接近CO2分子直径的微孔孔径和较大的比表面积时,才能有效提高对CO2的物理吸附能力.
孔隙中的表面官能团与CO2分子之间的作用和化学反应也是活性炭捕获CO2的主要途径. 含氧官能团在孔表面通过产生负电荷来增强酸性CO2分子和孔表面之间的作用力[23]. 但表面官能团和CO2分子之间的作用有一定的选择性,如孔径较适宜的超微孔活性炭吸附剂的表面官能团(碳酰、醌、吡喃酮、苯并吡喃、羧基、内酯等含氧官能团)对活性炭的CO2吸附不产生影响[40]. 国内外关于氮掺杂活性炭的制备及其CO2吸附特性方面的研究受到越来越多的关注,因为表面含氮官能团能通过增强活性炭表面对CO2的亲和力从而显著提高活性炭的CO2捕集量[35,40-41]. 孔径为2.3 nm的有序介孔活性炭负载40%聚乙稀亚胺(PEI的最佳负载量)后,孔径缩小,孔隙表面上的氨基官能团增加,使其在35 ℃下CO2的吸附量从0.689 mmol/g升至1.234 mmol/g[27]. Singh等[42]研究显示,氮掺杂后多数活性炭在常温常压下的CO2吸附量提高了3~4倍. 此外,郭慧娴等[43]研究显示,在CO2吸附过程中孔隙中的NaOH分子通过形成Na2CO3将活性炭的CO2吸附量提高了4倍.
由表2可见:葡萄树枝基活性炭与CO2活化制备的其他未改性活性炭相比,其CO2吸附量相对较低;橄榄油渣、扁桃壳和橄榄石基活性炭的CO2吸附量较好,可能与较好的微孔体积比例及比表面积有关;孔径、比表面积、微孔体积和微孔体积比例较小的椰子壳活性炭的CO2吸附量较葡萄树枝基活性炭大. 但以上活性炭表面的官能团类型和数量都没有被表征,故无法进行综合对比. 葡萄树枝基活性炭虽然有较合理的微孔孔径,但比表面积较小,表面上有助于吸附CO2的活性官能团可能较少,所以达不到理想的吸附量值. 综上,还需深入研究葡萄树基活性炭孔隙表面上有利于吸附CO2的其他活性成分及其表征方法,以及在缩小孔径的同时,提高比表面积的制备条件.

表2 CO2物理活化活性炭在常温常压(25 ℃、101 kPa)下的CO2吸附量比较
2.3 热重特性
热重分析(TGA)是根据样品在不同温度下的质量变化来分析样品热解动力学的一种方法. 该研究中通过热解和活化制备的两种活性炭质量是原料的25%左右,仍有大量碳元素存在. 热重分析仪分析的活性炭在氮气中的热解特性如图4所示. 由图4可见,在氮气气氛下两种葡萄树基活性炭及其CO2饱和产物的热解过程可分为4个阶段. 第一阶段为30~45 ℃,在此温度区间内原始活性炭和CO2饱和活性炭的曲线不重合,两种饱和活性炭失去的质量略大于其CO2平衡吸附量,此过程主要是CO2从孔隙表面脱附的过程. 葡萄树皮基活性炭及其饱和产物的质量损失均约为3.23%,二者相差不大,说明孔隙中的大部分CO2分子迅速被切换的N2吹走,且CO2吸附量也较少,故此温度区间内没有显示出明显的质量变化. 葡萄树枝基活性炭及其吸附CO2产物在该阶段中的质量损失分别为1.72%和5.45%,差值(3.73%)小于葡萄树枝基活性炭的CO2平衡吸附量(4.82%),说明CO2分子相对牢固地附着在孔表面,温度的升高才能使其脱附;其余1.09%的质量损失包括少量热不稳定物质和少量水分的质量.

图4 葡萄树枝基活性炭和葡萄树皮基活性炭的热重曲线(TGA)
第二阶段为挥发性有机物的挥发或热稳定性较好的有机物的分解阶段,两种活性炭的温度范围有区别. 葡萄树皮基活性炭的温度范围为45~620 ℃,而葡萄树枝基活性炭分两段,分别为45~570 ℃和 570~650 ℃. 在该阶段失重曲线相对平缓,两种原始活性炭失重率较小,其CO2饱和活性炭失去的质量略大于原始活性炭失去的质量. 葡萄树枝基活性炭及其饱和产物在570~650 ℃范围内的质量损失均为0.40%.
第三阶段为葡萄树基活性炭热解的主要阶段. 葡萄树皮基活性炭的主要分解范围为620~730 ℃,在该区间的质量损失为16.72%,其CO2饱和产物在620~740 ℃的质量损失为17.50%,吸附CO2使活性炭的热解温度范围拓宽10 ℃;葡萄树枝基活性炭在该阶段内质量降低缓慢,没有明显转折点.
第四阶段为灰化阶段(740~1 000 ℃). 当热解温度达 1 000 ℃时,仍没有被热解的部分是剩余的灰分和固定碳,葡萄树皮基活性炭的灰分和固定碳分别为66.65%和67.61%,葡萄树枝基活性炭的灰分和固定碳分别为83.57%和87.78%.
活性炭在不同温度范围内的质量变化是不同有机物分子热解所致. 在31~290 ℃区间内的失重由水分子蒸发和易热解有机物的分解所致[44]. 半纤维素分解温度较低,一般低于350 ℃时就会发生大量分解,纤维素大部分在315~400 ℃发生分解[45-46]. 小分子量有机物随着温度的升高而缓慢释放,这也会引起样品质量的降低. Ncibi等[47]研究显示,半纤维素、纤维素、木质素裂解温度分别为290、330、400 ℃. 因此,第二阶段主要是半纤维素和纤维素的热解阶段. 木质素热裂解的温度范围较宽(200~700 ℃),木质素热解过程中会生成较多的焦炭,失重速率随之降低,一般大部分木质素热解在400 ℃后才出现[48]. 240~670 ℃区间内的重量损失是由蛋白质、酸类及碳水化合物等有机物的分解所致[44]. 葡萄树枝和树皮基活性炭的热解制备过程包括500 ℃的碳化(60 min)和800 ℃的活化过程(60 min),因热解时间短样品中还有很多没有被热解的木质素和木质纤维,所以不同温度范围内的质量损失是由制备时未完全热解的蛋白质、酸类、碳水化合物、木质素以及纤维素等的热解导致.
吸附CO2前、后的活性炭的质量损失曲线非常相似,尤其是45 ℃后的曲线形状基本一致,曲线在垂直方向上的错位接近活性炭的CO2平衡吸附量,这表明饱和活性炭成分和结构与原始活性炭相比没有明显区别,但饱和活性炭和原始活性炭的热重曲线在45 ℃后不完全重合. 失重曲线上的差别间接反映了活性炭表面含氧官能团性质和数量上发生的变化. 活性炭表面含氧官能团的热分解温度研究[49-50]显示,羧酸在低于400 ℃、内酯基在600~650 ℃的温度区间热分解释放出CO2,羧酸酐在600~650 ℃热分解时释放出CO2与CO,酚和醚在700 ℃以上、羰基和醌基含氧官能团在900 ℃以上的温度区间热解释放出CO. 由图4(a)可见,葡萄树皮基活性炭及其CO2饱和产物的热重曲线垂直平移后,在620~730 ℃及高于900 ℃的温度区间内不重合的部分较明显,内酯基、羧酸酐、酚、醚、羰基和醌基等含氧官能团均可能发生变化. 图4(b)中不重合的曲线温度范围为650~1 000 ℃,在该温度范围内酚、醚、羰基和醌基等含氧官能均可能发生变化;接近1 000 ℃时曲线仍未呈稳定趋势,表明固定碳的热解反应仍未结束. 活性炭吸附CO2前、后表面官能团的变化需要用红外光谱及其他表征方法进一步研究.
3 结论
a) 两种葡萄树基活性炭的微孔体积占比均较大,其中孔径接近0.66 nm的微孔体积占比较大,其中葡萄树枝基活性炭主要以微孔为主,其30 ℃常压下CO2吸附容量为1.096 mmol/g. 葡萄树皮基活性炭具有大量微孔和少量中孔,其CO2吸附容量为0.247 mmol/g. 葡萄树枝基活性炭的CO2吸附容量明显高于葡萄树皮基活性炭,与其他CO2物理活化制备的未改性活性炭在常温常压下的CO2吸附容量相比,这两种葡萄树基活性炭CO2吸附容量均较低,CO2吸附量较高的葡萄树基活性炭的制备条件和改性方法需要进一步研究.
b) 随着温度的升高,两种葡萄树基活性炭及其CO2饱和产物失去的质量不完全相等,饱和活性炭在45 ℃之前释放全部CO2,活性炭对CO2的吸附主要以物理吸附为主,两种活性炭与其CO2饱和产物的热重曲线不重合的现象说明二者在表面官能团上的差异. 葡萄树枝基活性炭的CO2吸附性能比葡萄树皮基活性炭稳定.
猜你喜欢
快乐作文(3.4年级)(2023年8期)2023-05-01 09:04:00
今日农业(2022年2期)2022-11-16 12:29:47
童话王国·文学大师班(2022年3期)2022-04-19 14:01:36
中国特种设备安全(2021年4期)2021-10-13 06:42:14
小哥白尼(野生动物)(2018年4期)2018-08-01 03:41:14
小溪流(画刊)(2017年7期)2017-08-09 19:49:55
创新作文(小学版)(2017年33期)2017-04-04 02:02:10
快乐作文·高年级(2017年3期)2017-03-24 14:08:35
童话王国·文学大师班(2016年6期)2016-11-10 06:10:33
中学政史地·教学指导版(2014年10期)2015-02-02 08:59:38
