
我国南方籼稻整精米率发展趋势及对策
2021-07-16 06:41邹禹钱宝云占新春郑乐娅张培江
安徽农业科学 2021年11期
邹禹 钱宝云 占新春 郑乐娅 张培江


摘要 统计2009—2018年我国南方籼稻整精米率检测数据,表明我国南方籼稻品种的整精米率呈逐年下降的发展趋势,整精米率已经成为优质籼稻发展的限制因素。从品质性状遗传、生态环境、栽培管理、稻米加工等方面分析整精米率的影响因素,提出了相应的对策。结合优质常规籼稻的品种选育和优质杂交籼稻配组的实践,对优质籼稻育种进行展望。提高我国南方稻区籼稻品种的整精米率,对提高籼稻种植效益和加工效益、保障我国粮食安全、提升我国稻米在国际市场中的竞争力均具有重要意义。
关键词 籼稻;整精米率;发展趋势;对策
中图分类号 S-511.2+1 文献标识码 A
文章编号 0517-6611(2021)11-0038-08
doi:10.3969/j.issn.0517-6611.2021.11.011
开放科学(资源服务)标识码(OSID):
The Development Trend and Countermeasures on Head Rice Rate of Indica Rice in Southern China
ZOU Yu,QIAN Bao-yun,ZHAN Xin-chun et al
(Rice Research Institute,Anhui Academy of Agricultural Sciences,Hefei,Anhui 230031)
Abstract Through the statistics of the detection data of Indica rice head rice rate in southern China from 2009 to 2018,we found out that the head rice rate of Indica rice varieties was decreasing year by year,which indicated that the head rice rate had restricted the development of high-quality Indica rice.We analyzed the influence factors of quality trait inheritance,ecological environment,cultivation mode,fertilizer and water management,rice processing,and put forward corresponding countermeasures.Combined with the practice of the breeding of good quality Indica conventional rice varieties and good quality Indica hybrid rice,the breeding of good quality Indica rice was prospected.It was of great significance to improve the head rice rate of Indica rice varieties in southern china,to enhance the benefit of Indica rice planting and processing,and to ensure the food security of China and enhance the competitiveness of Chinese rice in the international market.
Key words Indica rice;Head rice rate;Development trend;Countermeasure
我國水稻种植范围广,品种多且地区间差异较大。2010年以来,我国在水稻主产区实施稻谷最低收购价政策,调动了种粮积极性,全国水稻种植面积稳定在3 000万hm2以上,2018年面积最大达到3 018.9万hm2;单产和总产总体发展趋势为逐年增产,单产在2010年超过6.55 t/hm2,2018年达7.02 t/hm2;稻谷总产量在2010年突破20 100万t,2018年达到21 213.0万t[1]。近几年,我国水稻生产持续丰收,稻谷市场供给充足,但我国仍然持续大量进口泰国、越南和巴基斯坦等国家的低价大米,给国内籼米市场造成较大冲击,同时也造成国内稻谷库容紧张。分析原因,一方面是我国稻米生产成本稳定增加、国内外大米价格差距较大;另一方面我国南方稻区的籼稻的稻米品质,尤其是整精米率与泰国等国家的籼稻相比还有一定差距,造成了我国籼稻市场“稻强米弱”的格局。
2002—2009年全国稻米整精米率的统计结果显示,整精米率年平均值总体徘徊在55.2%上下,范围涵盖0.2%~879%[2],年度间全国稻米的整精米率处于总体平稳有所波动的状态,而从平均值看年度间波动不大于 5 百分点。由此可见,我国稻米的整精米率总体偏低,品种的整精米率更是参差不齐。笔者根据中国水稻产业发展报告,研究2009—2018年我国南方籼稻整精米率的变化趋势,分析南方籼稻整精米率变化的影响因素,并提出提高南方籼稻整精米率的策略,为提高我国南方籼稻品质遗传改良提供参考。
1 我国南方籼稻整精米率发展趋势
稻米品质一般由碾磨、外观、蒸煮食味和营养品质组成。在我国现行标准中采用的各项品质指标中,糙米率、整精米率、碱消值、胶稠度的数值越高,稻米品质越好;垩白率、垩白度与透明度的数值越低,稻米的品质越好;直链淀粉含量数值适中品质好;蛋白质含量数值越高其营养品质越好,但高蛋白质含量的稻米食味会受到影响。
整精米率是稻米碾磨品质的关键指标,直接影响出米率,无论何种类型的优质稻,均要求稻谷有较高的整精米率。整精米率的高低主要决定于稻谷对脱壳、碾磨过程中机械压力的承受能力,当机械压力超过米粒承受范围时,容易使米粒碎裂,造成整精米率下降。影响整精米率的因素很多,如直链淀粉含量、蛋白质含量、胶稠度等,而稻米本身的性质,如裂痕、垩白、不成熟粒率、粒形等也会对稻米的碎米率产生很大影响[3]。
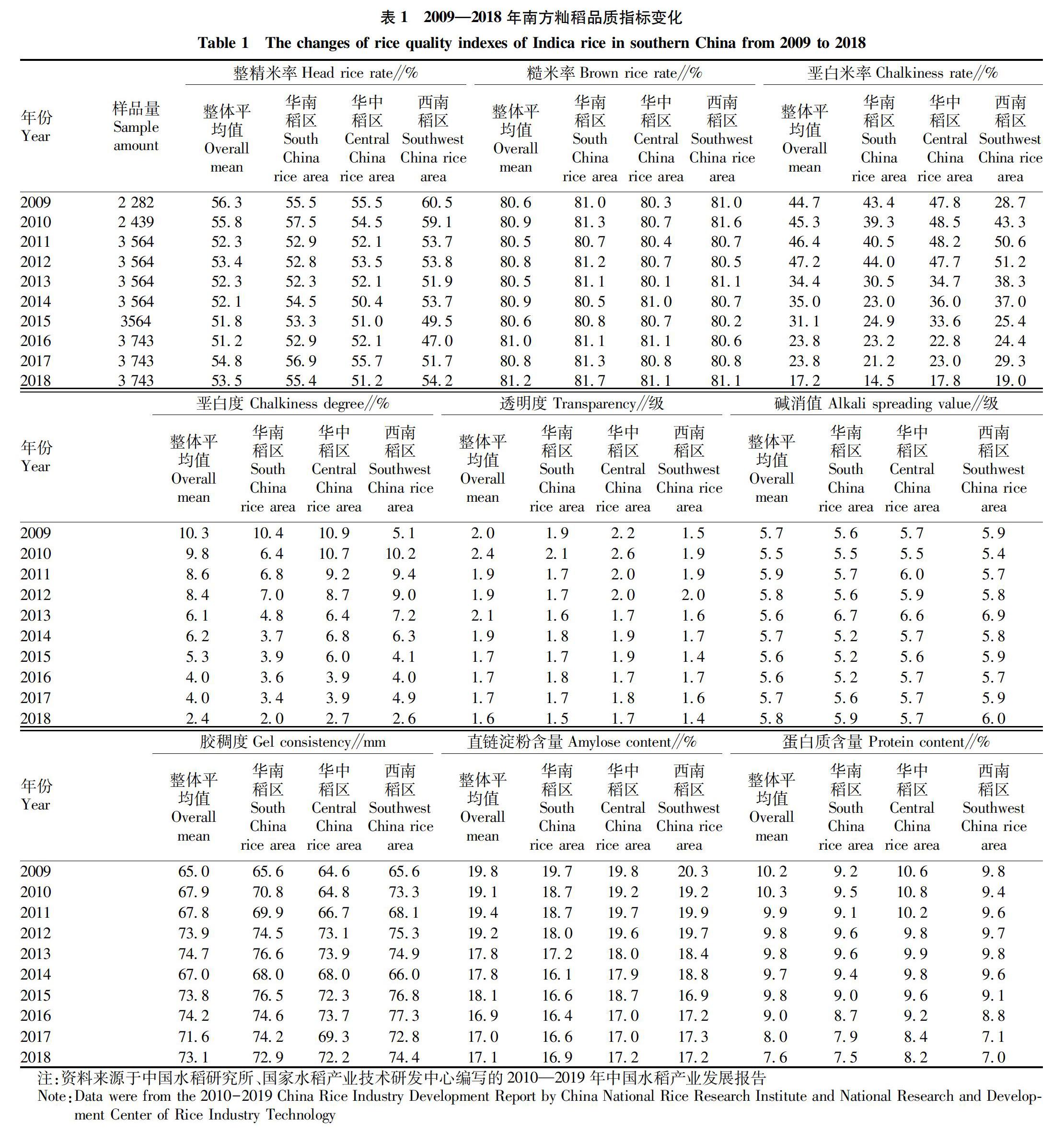
我国水稻育种家经过多年努力,在南方籼稻的品质改良上做了大量的工作,垩白米率、垩白度、透明度、胶稠度、直链淀粉和蛋白质含量等得到了较大改善,而整精米率却呈下降趋势,糙米率和碱消值变化不明显。从表1可以看出,垩白米率和垩白度总体为下降趋势,2018年分别平均为17.2%、2.4%,较2009年分别下降了27.5和7.9百分点;透明度总体向好态势,2010年平均为2.4级,2018年平均为1.6级;胶稠度呈提高趋势,从2009年的65.0 mm提高到2018年的73.1 mm;直链淀粉含量和蛋白质含量呈降低趋势,2018年分别平均为17.1%、7.6%,较2009年分别下降了2.7和2.6百分点。由此可见,2009—2018年,我国南方籼稻外观品质和蒸煮品质质量指数都得到了明显提升。
但是,2009—2018年我国南方籼稻的整精米率总体上呈下降趋势,与2009年相比,2016年下降了5.1百分点;2017—2018年有所回升,但仍未达到2009年水平。从华南、华中、西南3个稻区来看,也都呈下降趋势。华南稻区整精米率下降较少,2016年较2009下降了2.6百分点。西南稻区的整精米率下降幅度较大,2016年较2009年下降了13.5百分点,2017、2018年虽有回升,但与2009年相比还相差较大,分别相差8.8和6.3百分点。华中稻区是我国籼稻的主产区,该区域的籼稻整精米率也呈下降趋势,2014年的整精米率最低,仅为50.4%,较2009年下降了5.1百分点,此后有所回升但不稳定,至2018年达到51.2%,和2009年相比,仍差4.3百分点。
根据中国水稻研究所、国家水稻产业技术研发中心编写的2010—2019年中国水稻产业发展报告,2009—2018年籼稻整精米率的测试结果,得出整精米率的达标率较低,总体呈下降趋势,2016年籼稻整精米率达标率仅为56.7%,在所有测试的品质指标中达标率最低,而2009年籼稻的整精米率达标率为91.0%。尽管这10年来南方籼稻的外观品质和蒸煮品质都得到明显改良,但碾米品质的整精米率却显著下降。随着国内外稻米市场对长粒优质籼稻的青睐,整精米率将成为我国南方尤其是华中稻区优质籼稻发展的限制因素。
按应用类、区试类和选育类来看(图1),3种类型的整精米率均呈现下降趋势。应用类整精米率变化幅度较大,2009年为58.5%,而2011、2014年分别仅为48.4%、48.1%,较2009年分别下降10.1和10.4百分点,2016—2018年分别回升到52.1%、55.8%、55.6%。区试类整精米率变化幅度较应用类小,2009—2016年呈下降趋势,2016年较2009年下降了5.4百分点,2017、2018年向好变化,分别达55.8%、54.7%。
选育类整精米率在年度间差异较大,2009—2013年变化较小,在53.0%~56.5%,而2014—2018年,除2015年达到530%外,其他均较低,2016—2018年分别仅为46.9%、503%、49.7%。区试类和选育类的整精米率变化趋势将会影响未来我国南方籼稻市场的整精米率,由此也说明我国籼稻的整精米率改良状况不容乐观。
2 籼稻整精米率的影响因素
2.1 遗传因子对整精米率的影响
整精米率是多基因控制的数量性状,主要受水稻胚乳基因型控制或种子母体基因型控制,此外2者之间又有显著的细胞质效应[4]。目前通过QTL定位,已经发现35个关于整精米率的QTL。由表2可知,梅捍卫等[5]利用籼粳交组合特青/Lemont 进行QTL定位,共定位到4个影响整精米率的QTL(qHr2、qHr4、qHr6和qHr7);梅德勇等[6]利用籼籼交组合特青/IRBB衍生的重组自交系群体定位到2个整精米率QTL(qHRR3和qHRR5);Tan等[7]使用籼籼交组合珍汕97B/明恢63定位到1个整精米率QTL;Septiningsih等[8]利用种间杂交组合IR64/IRGC 定位到3个整精米率QTL。尽管发现众多的关于整精米率的QTLs,但均没有进行基因的精细定位和克隆,难以将这些QTL用于分子标记进行辅助育种。因此对稻米整精米率的基因精细定位及克隆还需进一步深入研究。
杂交稻的整精米率要偏低于常规稻,且受品种遗传影响[15]。在水稻杂交育种中,其品质的性状表现与产量性状相比,整体呈现相反的趋势[16]。进一步研究发现,杂交稻的整精米率与其亲本呈显著正相关,尤其与母本不育系相关性更大[17-18]。因此杂交水稻的高整精米率育种难度较大,要育成优质杂交稻,则必须选用高整精米率的双亲配组。
在与环境互作的研究中发现,稻米整精米率会明显受到各遗传效应与环境互作效应的影响[19]。徐斌等[20]研究发现,两加性效应QTL的总贡献率小于与环境互作的总贡献率。因此遗传研究需要在不同环境条件下进行,并在遗传效应和遗传×环境互作效应的遗传模型下进行双重分析。尽管稻米的整精米率是受多重因素影响,但却存在一些能在不同群体和环境下稳定表达的主效QTL。因此,研究者可以通過在不同遗传背景和环境下对整精米率相关基因进行精细定位,并在整精米率改良育种中加以利用,有望改善我国南方整精米率相对较低的状况。
2.2 品质性状间的相关性分析
基因具有一因多效和连锁特性,因此稻米品质性状之间存在着错综复杂的关系。粒重、米粒宽、精米率、垩白米率和糙米率与整精米率相关[21]。徐富贤等[22]在研究杂交中稻过程中也发现,长宽比和垩白米率是影响整精米率的关键因子,其中垩白米率与整精米率呈极显著负相关。然而不同类型籼稻品种之间也有差异性,王丹英等[23]研究发现,精米率在1.9%~52.7%的常规籼稻品种间垩白米率和垩白度的差别并不能以整精米率反映出来,其整精米率和垩白的相关性均不如杂交籼稻密切,常规籼稻整精米率与直链淀粉含量呈极显著正相关,而杂交籼稻整精米率与蛋白质含量呈极显著正相关。
分析2009—2018年南方籼稻稻米品质指标,包括整精米率、糙米率、垩白米率、垩白度、碱消值、胶稠度、直链淀粉含量和蛋白质含量,垩白米率、垩白度、直链淀粉含量和蛋白质含量从2009—2018年均有不同程度的降低(表1)。但通过Pearson 相关性分析,2009—2018年南方籼稻整精米率下降与其他品质性状的变化无显著相关性。按稻区看,华南、华中稻区的整精米率下降与其他品质性状的变化也无显著相关性;西南稻区的整精米率下降与直链淀粉含量降低显著正相关(表3)。
2.3 生态环境对整精米率的影响
我国地域广阔、地形复杂,南方稻区覆盖了热带、亚热带和温带区域。受区域环境特点的影响,我国籼稻整精米率也表现出区域性特点。
在各环境因子中,温度是影响稻米整精米率最显著的因子。研究发现水稻冠层温度差与稻米整精米率呈线性负相关[24],特别是灌浆结实期的温度,水稻灌浆结实期温度以21~26 ℃为宜[25]。目前研究发现环境温度尤其灌浆期的夜间高温对稻米的精米率、整精米率、垩白度、黏度曲线、糊化温度等都有非常重要的影响。Lyman等[26]
报道季节温度每增加1℃,稻米产量将减少6.2%;精米产量减少7.1%~80%,整精米减少9.0%~13.8%。这主要是由于高温使稻米产量减少、垩白和破损米粒增多综合作用引起的。Liu等[27]研究高温处理对不同品种稻米品质的影响,高温处理从抽穗期到成熟期,越光和IR72共2个品种均表现垩白米粒增多,整精米率、直链淀粉和总淀粉含量降低。高温或低温均可显著降低稻米的整精米率[28]。不同类型水稻的整精米率对温度也表现出不同的响应,粳稻的整精米率与温度呈线性负相关,而籼稻则与温度呈二次曲线、抛物线型[29]。马国辉[30]认为,在21~23 ℃是提高籼稻整精米率的最适温度。天气条件引起的不利于蒸发和蒸腾冷却的环境是稻米垩白产生的一个主要因素[31]。垩白是稻米重要的外观品质,且会直接影响稻米的整精米率。前人多将垩白归咎于遗传因素以及高温尤其是灌浆期的高温。全球变暖这一因素在干燥季节对稻米品质具有增强效应,在雨季则起负面作用,使得垩白增加、整精米率降低。但是,全球变暖并不是影响稻米垩白的主要因素。
湿度对水稻整精米率影响的研究较少,主要由于湿度测定准确性差,研究难度较大。但通过前人的研究[32],推测高湿环境可能在水稻的某个生育期直接或间接影响稻谷的整精米率,特别是籼稻,这需要进一步的试验验证。
光照对稻米整精米率也有较大的影响[25]。弱光会降低稻米的整精米率[33],这可能与弱光引起的稻米自身蛋白质含量降低、籽粒充实不良和胶稠度变硬等不良现象有关。
在种植、收获、干燥和储存过程中,由于米粒之间或者米粒与环境之间水分的差异,米粒会吸水或者失水,在此过程中米粒内部会形成一个吸湿应力,当应力超过米粒的机械强度时就会形成裂痕(多为内部裂痕),而米粒的裂痕或者内部裂痕都是米粒破裂的一个主要原因[3],米粒的破裂直接影响整精米率。
2.4 栽培方式及肥水管理的影响
Siebenmorgen等[34]对米粒形成过程至收获前影响稻米品质的因素进行了综述,指出稻米的品质与品种的农艺性状及种植时间、灌溉、施肥量、收获时的条件等均有一定的相关性。
栽培技术对稻米整精米率具有不可忽视的作用,合理的栽培和管理能够增加稻米的整精米率。大田密度过大会降低稻米的整精米率[35]。因此,在栽培过程中应兼顾产量和整精米率,通过合理密植达到双赢的目的。
研究认为,总施氮量的增加可以提高稻米的整精米率[36-37],尤其是在穗肥期。而过量施用氮肥会降低稻米的整精米率[38],同时,增加土壤的有机肥也可以提高稻米的整精米率[39]。
2.5 高整精米率优质长粒籼稻栽培与育种实践
选用广东等地育成的长宽比在3.4以上的优质常规籼稻品种11个,2019年在全椒育种基地种植,5月20日播种,抽穗期在8月上中旬。收获前保持土壤湿润,完熟时人工收获,采用小型脱粒机脱粒,晾干后在室内放置50 d左右进行品质分析,检测结果如表4。这11个品种的整精米率平均达66.0%,其中米粒长宽比3.4的4个品种的整精米率均在60.0%以上,平均66.9%,粤油丝苗整精米率最高为70.8%,平均垩白粒率和垩白度分别为7.2%和1.4%;米粒长宽比3.6的4个品种的整精米率均在60.0%以上,平均66.4%,齐华占整精米率最高为71.3%,平均垩白粒率和垩白度分别为9.9%和35%;米粒长宽比4.3以上的3个品种整精米率在63.4%~65.8%,平均64.4%,平均垩白粒率和垩白度分别仅为29%和1.2%,长宽比4.3以上的品种垩白粒率和垩白度明显低于长宽比为3.4和3.6的品种的垩白粒率和垩白度。这说明长宽比在4.3以上的优质常规籼稻品种在优化生长环境、晾晒条件下整精米率仍然可以达到60%以上,通过加大米粒长宽比,可以选育到低垩白粒率和垩白度的常规籼稻品种。
近年来,安徽省农业科学院水稻研究所致力于優质杂交籼稻育种,重点选用株型好整精米率高的不育系与优质恢复系配组。2019年配组576个优质杂交籼稻组合在全椒育种基地种植,5月8日播种,小区面积3.24 m2,每个小区种植90穴,每穴插单苗,初步筛选出10个优质高产杂交籼稻组合。这10个组合的株高在115 cm上下,株型好,叶片上举,抗倒伏能力强;产量在11.30~11.96 t/hm2,较对照丰两优4号增产4.8%~11.3%;始穗期在8月5—15日,全生育期135~140 d;灌浆结实期处于较高温度气候条件下,结实率仍达到85.2%~95.2%。
尽管在较高温度条件下灌浆,收获前保持土壤湿润,完熟时人工收获,采用小型脱粒机脱粒,晾干后在室内放置50 d 左右进行品质分析,检测结果如表5。
由表5可知,10个杂交籼稻组合的米质总体上表现较优良,主要表现在米粒长宽比3.8~4.1,垩白粒率在4.9%~173%,垩白度1.3%~5.8%,而整精米率高,在66.2%~705%。胶稠度仅1个为78.5 mm,其余9个组合的胶稠度在80.0 mm以上,最高达88.0 mm,直链淀粉含量14%左右,食味均较好。
这些组合的育成说明在杂交籼稻的双亲选育上,注重优质尤其是高整精米率的不育系和恢复系选育,杂交籼稻的优质和高产是可以结合的,也能够育成适宜长江中下游稻区较高温度条件下种植的垩白粒率和垩白度较低而整精米率较高的优质杂交籼稻组合。
3 提高南方籼稻整精米率的意义
3.1 提高整精米率可以提高水稻种植效益和加工效益
稻米品质主要包括碾磨品质、外观品质、蒸煮食味品质和营养品质,其中碾磨品质的优劣主要体现在糙米率、精米率和整精米率[40]。在国家《优质稻谷》标准和部颁《食用稻品种品质》标准中都将整精米率列为重要指标,无论何种类型的优质稻,均要求稻谷有较高的整精米率。因此整精米率作为稻米品质的重要性状之一,直接关系到稻米的商品价值和稻农的经济效益[41]。
3.2 提高籼稻整精米率可以提高我国粮食安全的保障能力
根據中国水稻产业发展报告,2009—2018年间,我国水稻年种植面积3 000万hm2以上,总产超过2亿t,平均单产6.75 t/hm2左右,单产总体呈上升趋势,但很缓慢,平均年增产仅为0.73%。我国南方籼稻种植面积约占全国水稻总面积的67%,而南方籼稻的整精米率总体不高,2009年平均为56.3%,此后呈递减趋势,到2016年平均仅为51.2%。尽管水稻单产增产幅度有限,而整精米率还有较大的上升空间。如果通过遗传改良,将籼稻的整精米率恢复到2009年水平,即为56.3%,以单产6.75 t/hm2计算,所产的整精米量相当于整精米率51.2%的籼稻单产7.42 t/hm2的量,与单产6.75 t/hm2相比增产9.96%。整精米率的4百分点之差,对于加工企业效益也是非常可观的。因此,提高籼稻整精米率不仅可提高加工效益,还可以显著提高我国稻米产量,从而提高我国粮食安全的保障能力。
3.3 提高整精米率可以提升我国稻米在国际市场中的竞争力
我国稻米分级标准的各级别碎米率含量都较高,其中籼稻米碎米率含量为30%~35%,而泰国出口大米含碎率只有25%[42],仅从整精米率来看,我国籼米在国际市场中质量不如泰国米。整精米在蒸煮过程中,一般饭粒开裂少,饭粒表面光滑圆润,韧性较好,适口性好。整精米率低的稻谷在加工过程中,产生的碎米较多,直接降低了稻谷的商品价值。含碎米较多的商品米,蒸煮的米饭外观也较差,也影响米饭的食味。因此,提高整精米率可以提升我国稻米在国际市场中的竞争力。
3.4 提高整精米率有利于提高种子质量
我国南方籼稻种子生产收获前常会遇到持续的高温多雨天气,水稻穗发芽现象很常见,穗发芽不仅影响种子质量,也会降低合格种子产量。容易发生穗芽的种子收获时遇雨,在储藏过程中发芽率下降也会快些。Buggenhout等[3]认为米粒破裂的一个主要原因是米粒具有裂痕或者内部有裂痕,裂痕对稻米的整精米率影响很大。整精米率高的米粒机械强度要大于米粒与环境水分交换过程中形成的吸湿应力,米粒机械强度大的一般致密度要好,吸水慢而放缓穗发芽。整精米率高的水稻种子在收获、干燥、精选、包装等种子加工过程中,形成的米粒裂痕少且种子发芽势好,有利于提高秧苗素质和植株生长发育一致性。因此,整精米率高有利于提高种子质量和秧苗素质,从而有利于构建高产群体。
4 提高南方籼稻整精米率的策略
4.1 优质遗传资源筛选与利用
决定整精米率高低的关键因素是自身整精米率基因的表达,目前还没有克隆到与整精米率直接相关的基因。因此,为了减少对整精米率选育的盲目性,还需加强整精米率基因的挖掘与利用。杂交籼稻的整精米率要普遍低于常规籼稻[15],且与亲本的整精米率极显著相关,因此在选择亲本组配时要优先选择高整精米率的亲本,尤其要注重选择高整精米率的母本[18]。
加大耐热种质资源的利用,在遇到高温天气时,耐热品种不仅可以降低产量的损失,还可以一定程度上减少对稻谷整精米率的影响。
抗病资源筛选与利用,稻瘟病尤其谷粒瘟、白叶枯病、纹枯病等病害都会影响稻谷的整精米率。因此,加强抗稻瘟病基因Pi-2、Pi-9、Pigm和抗白叶枯病基因Xa23在籼稻育种中应用,增强籼稻品种的抗病性,降低病害对整精米率的影响。
4.2 通过株型改良,改善田间群体小气候
温度和CO2会影响稻米品质形成。Figueiredo等[43]利用开放式顶部腔室,评估温度升高的影响或温度升高和CO2浓度升高的综合效果,研究表明温度和CO2浓度升高对水稻产量有明显负效应。Wang等[44]分析了CO2浓度和温度对籼稻、粳稻、野生稻的影响。结果显示,在高CO2浓度下生长的水稻比其他条件下水稻对高温胁迫更敏感。Usui等[45]研究了CO2富集、土壤和水变暖对稻米产量和品质的影响。结果表明,随着大气CO2浓度升高、土壤和水的升温均能够降低稻米外观品质,CO2浓度的影响最大。CO2浓度升高能够降低稻米蛋白质含量,也增加了垩白粒率。由于垩白粒率的增加导致整精米率显著降低。垩白粒率对蛋白质含量的依赖性在不同年份表现不同。稻米外观质量对年际变化的响应,表明CO2浓度升高与空气温度协同作用会降低稻米的外观品质。我国南方籼稻主产区的长江中下游稻区的中籼稻抽穗扬花期经常处于高温、高湿条件下,而且大部分中籼稻品种叶片较大,田间密闭,群体内通风透光条件较差,田间CO2浓度要高于群体通风透光条件好的,高CO2浓度不利于提高水稻耐热性,同时影响稻米外观品质和整精米率。
不同株型的水稻群体可以形成不同的田间小气候,主要表现在温度、湿度和光照的差异,因此不同株型水稻群体的田间小气候也会影响稻谷的整精米率[32]。通过改良水稻株型来优化田间群体内的小气候,提高水稻抗倒伏能力,以达到提高整精米率的目的。株型的改良涉及基因较多,主要包括:①优质水稻品种茎秆往往较软,抗倒伏能力弱。利用株型基因IPA1-2d、粗秆基因SCM2和半矮秆基因sd-1,降低水稻植株高度、增加茎秆粗度,提高抗倒伏能力。②利用分蘖角度控制基因tac1和半卷叶基因SRL1等,使植株株型内松外紧,剑叶内卷上挺,调节群体结构,改善群体内部通风透光条件。③利用直立穗基因DEP1,选育穗型中等偏大,一次枝梗长而多,着粒均匀,灌浆一致性好。④利用长粒基因GL7,选育籽粒长,长宽比大的品种,特别要注意籽粒的厚度,稻米横截面越圆越好,稻米加工品质好,整精米率高,商品性好。
4.3 通过栽培措施,优化群体结构
Haefele等[46]分析了全球水稻生产的土壤质量,种植水稻的土壤被分类为好、差、极差或问题土壤等,在全球范围内,1/3的水稻土壤较为贫瘠。因此,有条件的地方种植紫云英,增施有机肥,增加土壤有机质,改善土壤结构和通透性。
我国南方稻区土壤的有效态硼、硅和交换性镁含量較低[47],而这些微量元素的缺失会影响稻米的品质。因此适当增施硼、锌、硅等微量元素肥料能提高稻米的整精米率。
根据各地气候特点、水利条件、茬口以及品种生育期,兼顾水稻产量和品质,安排好播种期。田间栽插行向也是不可忽视的,根据太阳光照方向和风向确定,一般栽插行向为东西向,群体内部可以充分接收阳光和通风。通过施用多效唑或烯效唑等植物生长调节剂,化学控制水稻植株高度,缩短节间长度和叶片长度,优化群体结构,提高抗倒伏性。通过肥水调控,控制水稻植株高度和叶片长度,尤其是群体冠层叶片长度,调节田间群体结构,改善群体内部小气候,收获前保持田间土壤湿润对提高整精米率是非常重要,优化水稻植株生长环境,降低病虫危害,提高稻米品质,尤其稻米整精米率。
4.4 通过订单生产或分品种收购,提高加工质量
我国南方稻区生产上应用的籼稻品种较多,而在品种多途径审定后,生产上应用的品种将会更多。不同品种的籽粒粒型存在较大差异,如千粒重,大小相差可能达10 g以上,谷粒长宽比和厚度也不尽相同,尤其是不同厚度的稻谷混在一起进行糙米加工,对整精米率影响较大。为了降低种植风险,每个家庭农户可能都会种植多个品种,种植大户或合作社也是如此。在稻谷收购过程中,每个乡镇参与粮食经营的企业也较多,多为私营企业,而私营企业由于受到资金和库容的限制,很难做到分品种收购。水稻生产、收获、收购过程中,都会造成稻谷的混杂,均不利于整精米率的提高。因此,通过订单生产,分片种植,按品种单打单收,杜绝不同粒型品种混收。
米粒具有裂痕或者内部有裂痕是导致米粒破裂,从而影响整精米率。为了降低米粒产生的裂痕,收获时最好选用半喂入的收割机进行收获。稻米水分对糙米品质特性也有影响。Kim等[28]报道,破裂糙米比例随着水分的增加而增加,而糙米硬度随着水分的增加而减小。水分15%时的糙米率和异物最少,17%时不完善率和破损率最小。脂肪以及灰分含量随水分减小而增加,综合考虑认为水分为15%~17%时糙米的综合品质最好。因此,烘干时将温度控制在40 ℃以内进行低温慢烘,降低米粒与环境水分交换时产生的吸湿应力,减少裂痕,将稻谷水分含量控制在16%左右,有利于稻谷储存和稻米加工。从收获到加工尽量减少中间环节,防止混杂,加工时要根据加工要求更新设备,尤其是要添加使稻谷有序排列进入进行脱壳的设备,降低糙米的破损率和裂纹米率,可以显著提高整精米率。
5 展望
整精米率作为稻米品质的重要性状之一,有效提高稻米整精米率,可提高其出米率、食用品质和市场经济价值。多数籼稻品种存在整精米率偏低的问题,尤其是杂交籼稻更加突出,这主要是由于我国长期以来注重选育高产的水稻品种,而对品质的要求较低。如我国食用稻品质指标中,籼稻的整精米率一级在58%、二级在55%以上,即使整精米率达到58%,这与国内外稻米市场和加工企业对籼稻整精米率的要求仍有较大差距。因此,通过遗传改良,籼稻的整精米率提升空间较大[48]。
目前籼稻的整精米率已经成为优质籼稻育种的限制因素,这可能与整精米率易受环境影响和自身遗传复杂性有关。虽然对整精米率研究,也已经发掘多个QTL,但这些QTL还没有得到精细定位和克隆,甚至在不同的遗传背景和环境下定位也有差别。80%整精米率QTL,集中在第1、2、3、5、6、7号共6条染色体上,位置还是相对分散。因此希望利用分子标记辅助育种技术,定向选育高整精米率品种是有很大难度的,但是利用传统育种方法,加大选择压力,可以有效选育高整精米率的籼稻品种。广东等地已经育成不少优质高整精米率的常规籼稻品种,安徽等也育成高整精米率的优质杂交籼稻组合。
参考文献
[1]
徐春春,纪龙,陈中督,等.2018年我国水稻产业形势分析及2019年展望[J].中国稻米,2019,25(2):1-3,9.
[2] 卢林,朱智伟,段彬伍,等.我国稻米整精米率特点及环境影响因素分析[J].核农学报,2012,26(5):770-774.
[3] BUGGENHOUT J,BRIJS K,CELUS I,et al.The breakage susceptibility of raw and parboiled rice:A review[J].Journal of food engineering,2013,117(3):304-315.
[4] 石春海,何慈信,朱军.稻米碾磨品质性状遗传主效应及其与环境互作的遗传分析[J].遗传学报,1998,25(1):46-53.
[5] 梅捍卫,罗利军,郭龙彪,等.水稻加工品质数量性状基因座(QTLs)分子定位研究[J].遗传学报,2002,29(9):791-797.
[6] 梅德勇,朱玉君,樊叶杨.籼稻稻米碾磨与外观品质性状的QTL定位[J].遗传,2012,34(12):1591-1598.
[7] TAN Y F,XING Y Z,LI J X,et al.Genetic bases of appearance quality of rice grains in Shanyou 63,an elite rice hybrid[J].Theoretical & applied genetics,2000,101(5/6):823-829.
[8] SEPTININGSIH E M,PRASETIYONO J,LUBIS E,et al.Identification of quantitative trait loci for yield and yield components in an advanced backcross population derived from the Oryza sativa variety IR64 and the wild relative O.rufipogon[J].Theoretical & applied genetics,2003,107(8):1419-1432.
[9] 胡霞,石瑜敏,賈倩,等.影响水稻穗部性状及籽粒碾磨品质的QTL及其环境互作分析[J].作物学报,2011,37(7):1175-1185.
[10] 刘家富,奎丽梅,朱作峰,等.普通野生稻稻米加工品质和外观品质性状QTL定位[J].农业生物技术学报,2007,15(1):90-96.
[11] ZHENG T Q,XU J L,LI Z K,et al.Genomic regions associated with milling quality and grain shape identified in a set of random introgression lines of rice (Oryza sativa L.)[J].Plant breeding,2007,126(2):158-163.
[12] ALUKO G,MARTINEZ C,TOHME J,et al.QTL mapping of grain quality traits from the interspecific cross Oryza sativa × O.glaberrima[J].Theoretical & applied genetics,2004,109(3):630-639.
[13] 翁建峰,万向元,郭涛,等.利用CSSL群体研究稻米加工品质相关QTL表达的稳定性[J].中国农业科学,2007,40(10):2128-2135.
[14] LI J M,XIAO J H,GRANDILLO S,et al.QTL detection for rice grain quality traits using an interspecific backcross population derived from cultivated Asian (O.sativa L.) and African (O.glaberrima S.) rice[J].Genome,2004,47(4):697-704.
[15] 汪莲爱.环境条件与遗传因子对稻米整精米率的影响[J].湖北农业科学,2006,45(5):558-560.
[16] 廖伏明,周坤炉,阳和华,等.籼型杂交水稻杂种与亲本的稻米品质比较[J].中国水稻科学,2003,17(2):134-140.
[17] 张利华,王林友,王建军.籼型杂交稻稻米碾磨品质与外观品质的配合力及遗传力研究[J].核农学报,2003,17(6):417-422.
[18] 闵捷,朱智伟,章林平,等.中国超级杂交稻组合的稻米品质分析[J].中国水稻科学,2014,28(2):206-210.
[19] CHEN J G,ZHU J.Genetic effects and genotype×environment interactions for cooking quality traits in Indica-japonica crosses of rice (Oryza sativa L.)[J].Euphytica,1999,109(1):9-15.
[20] 徐斌,黄育民,黄建勋,等.籼稻整精米率QTLs的检测及上位性和环境互作效应分析[J].厦门大学学报(自然科学版),2007,46(4):578-582.
[21] 李耘,左永树,张长伟,等.籼型杂交稻整精米率的作用因子及改良研究[J].西南农业学报,2004,17(1):22-25.
[22] 徐富贤,熊洪,朱永川,等.杂交中稻整精米率的影响因子及其间接鉴定方法研究[J].西南农业学报,2013,26(1):1-6.
[23] 王丹英,章秀福,朱智伟,等.食用稻米品质性状间的相关性分析[J].作物学报,2005,31(8):1086-1091.
[24] 高继平,隋阳辉,张文忠,等.水稻灌浆期冠层温度对植株生理性状及稻米品质的影响[J].中国水稻科学,2015,29(5):501-510.
[25] 游晴如,黄庭旭,马宏敏.环境生态因子对稻米品质影响的研究进展[J].江西农业学报,2006,18(3):155-158.
[26] LYMAN N B,JAGADISH K S V,NALLEY L L,et al.Neglecting rice milling yield and quality underestimates economic losses from high-temperature stress[J].PLoS One,2013,8(8):1-9.
[27] LIU Q H,WU X,MA J Q,et al.Effects of high air temperature on rice grain quality and yield under field condition[J].Agronomy journal,2013,105(2):446-454.
[28] KIM S Y,LEE H.Effect of quality characteristics on brown rice produced from paddy rice with different moisture contents[J].Journal of the Korean society for applied biological chemistry,2013,56(3):289-293.
[29] 张嵩午,周德翼.温度对水稻整精米率的影响[J].中国水稻科学,1993,7(4):211-216.
[30] 马国辉.环境生态对中国稻米品质的影响[J].农业现代化研究,1998,19(3):146-149.
[31] ZHAO X Q,FITZGERALD M.Climate change:Implications for the yield of edible rice[J].PLoS One,2013,8(6):1-9.
[32] 周立宏,李秀芬,王伯伦,等.稻田小气候特征及其对水稻产量和品质的影响[J].江苏农业科学,2016,44(5):81-85.
[33] 李林.气候环境对稻米品质的影响研究进展[J].生命科学信息,1988,1(2):9-11.
[34] SIEBENMORGEN T J,GRIGG B C,LANNING S B.Impacts of preharvest factors during kernel development on rice quality and functionality[J].Annual review of food science & technology,2013,4:101-115.
[35] 张初阳.稻谷整精米率影响因素浅谈[J].中国稻米,2005,11(3):40-42.
[36] 嚴奉君,孙永健,马均,等.不同土壤肥力条件下麦秆还田与氮肥运筹对杂交稻氮素利用、产量及米质的影响[J].中国水稻科学,2015,29(1):56-64.
[37] 张自常,李鸿伟,曹转勤,等.施氮量和灌溉方式的交互作用对水稻产量和品质影响[J].作物学报,2013,39(1):84-92.
[38] 陶进,钱希旸,剧成欣,等.不同年代中籼水稻品种的米质及其对氮肥的响应[J].作物学报,2016,42(9):1352-1362.
[39] 陈帅君,边嘉宾,丁得亮,等.不同有机肥处理对水稻品质和食味的影响[J].中国稻米,2016,22(4):42-45.
[40] FITZGERALD M A,MCCOUCH S R,HALL R D.Not just a grain of rice:The quest for quality[J].Trends in plant science,2009,14(3):133-139.
[41] WANG Q,HUANG J L,HE F,et al.Head rice yield of “super” hybrid rice Liangyoupeijiu grown under different nitrogen rates[J].Field crops research,2012,134:71-79.
[42] 中国水稻研究所,国家水稻产业技术研发中心.2010年中国水稻产业发展报告[M].北京:中国农业出版社,2010.
[43] FIGUEIREDO N,CARRANCA C,TRINDADE H,et al.Elevated carbon dioxide and temperature effects on rice yield,leaf greenness,and phenological stages duration[J].Paddy & water environment,2015,13(4):313-324.
[44] WANG D R,BUNCE J A,TOMECEK M B,et al.Evidence for divergence of response in Indica,Japonica,and wild rice to high CO2 × temperature interaction[J].Global change biology,2016,22(7):2620-2632.
[45] USUI Y,SAKAI H,TOKIDA T,et al.Rice grain yield and quality responses to free-air CO2 enrichment combined with soil and water warming[J].Global change biology,2016,22(3):1256-1270.
[46] HAEFELE S M,BHATTACHAN B K,ADHIKARI B B,et al.Spatial variability of fertilizer management and response in rainfed rice of Nepal[J].Agriculture ecosystems & environment,2014,189:190-198.
[47] 刘国群,李飞.南方典型水稻种植区土壤有效态中微量元素分析[J].浙江农业科学,2015,56(11):1814-1816.
[48] 中华人民共和国农业部.食用稻品种品质:NY/T 593—2013[S].北京:中国农业出版社,2013.
猜你喜欢
古今农业(2022年2期)2022-08-15
中学生数理化·七年级数学人教版(2021年11期)2021-12-06
江苏安全生产(2020年3期)2020-04-21
云南教育·中学教师(2019年6期)2019-08-13
基层中医药(2018年11期)2019-01-31
西南农业学报(2016年4期)2016-05-17
世界热带农业信息(2014年3期)2014-08-12
