
利用InDel标记进行啤酒大麦黄花叶病抗性鉴定
2021-07-16 08:53:52栾海业杨红燕黄煜韬张英虎乔海龙沈会权
大麦与谷类科学 2021年3期
徐 肖,栾海业,杨红燕,黄煜韬,张英虎,臧 慧,陶 红,陈 和,陈 健,乔海龙,沈会权*
(1.江苏沿海地区农业科学研究所,江苏 盐城 224001;2.江苏天赋生物科技有限责任公司,江苏 盐城 224001)
大麦黄花叶病是由大麦黄花叶病毒(BaYMV)和大麦温和花叶病毒(BaMMV)引起的土传病害。大麦黄花叶病不但造成大麦减产,而且会降低籽粒品质[1]。我国长江中下游冬大麦产区是大麦黄花叶病的高发区,病田轻者减产30%~40%,重者颗粒无收。随着全球气候变暖,大麦适宜的生态区域增加,加上我国大型农机的大面积推广和使用,大麦黄花叶病发病面积可能进一步增加。由于寄主和病原菌相互作用,协同进化,不断产生出新的病毒株系,导致我国原有抗源材料单二、苏啤6号等优质啤酒大麦相继丧失抗性[2]。因此,如何利用有效抗病基因快速选育抗大麦黄花叶病新品种,成为当前啤酒大麦生产上亟待解决的重要问题。
目前,我国大麦黄花叶病主要依靠大田或病圃进行表型鉴定,需大量人力、物力来进行多年多点多个时期的鉴定。病害发生的严重程度与当年气候条件相关,年度间的病级不稳定,导致鉴定结果具有一定差异。分子标记是在DNA水平上鉴定抗病基因,进行抗病性选择时不受环境影响。利用分子标记辅助选择(molecular marker-assisted selection,MAS)可实现抗病基因在品种选育中的快速应用,有助于在生产上合理布局抗病基因资源和延长抗病品种使用年限[3]。
我国大麦黄花叶病病毒存在6个不同的BaYMV株系(A1、A2、B1、B2、B3和B4)与1个BaMMV株系[4],与日本的病毒株系有很高的相似度但又有所差异[5]。本研究利用InDel标记聚合法,对180份啤酒大麦资源进行检测,结合表型鉴定,评价标记检测的有效性,以期为选育啤麦抗黄花叶病新品种提供广谱性抗性资源。
1 材料与方法
1.1 供试材料
以收集的国内外200份啤酒大麦种质资源为材料,其中:80份由江苏沿海地区农业科学研究所提供,60份由扬州大学大麦研究所提供,60份由中国农业科学院作物科学研究所提供。
1.2 田间种植
分别于2019年和2020年秋季将参试材料种植于江苏沿海地区农业科学研究所南洋试验场(120.1614°E、33.3891°N)大麦黄花叶病病圃,采用随机区组设计,每个材料播种1行,每行点播40粒,设3次重复。
1.3 表型抗性鉴定
次年2月至3月对参试材料抗病性进行调查和鉴定,每个材料连续调查10株,每隔7 d调查1期,连续调查4期。调查时期:2020年2月20日、2月27日、3月5日与3月12日,2021年2月24日、3月3日、3月10日、3月17日。参照黄培忠等的标准[6]进行抗性级别判定:1级(高抗)—叶片叶色正常,无黄花斑点;2级(抗病)—叶片叶色基本正常,有黄花斑点,但斑点未连成线;3级(感病)—叶片病斑的斑点连成线,叶片黄化,但植株不矮化;4级(高感)—叶片出现大片黄花病斑,叶片黄化,植株萎缩、矮化,趋于死亡。
1.4 分子标记
利用InDel标记JSB056与JSB060进行基因型检测,其中:JSB056与rym5紧密连锁,JSB060与rym1紧密连锁;2个InDel标记均由徐婷婷等开发[7]。引物信息见表1,rym1与rym5的基本信息见表2[8-10]。
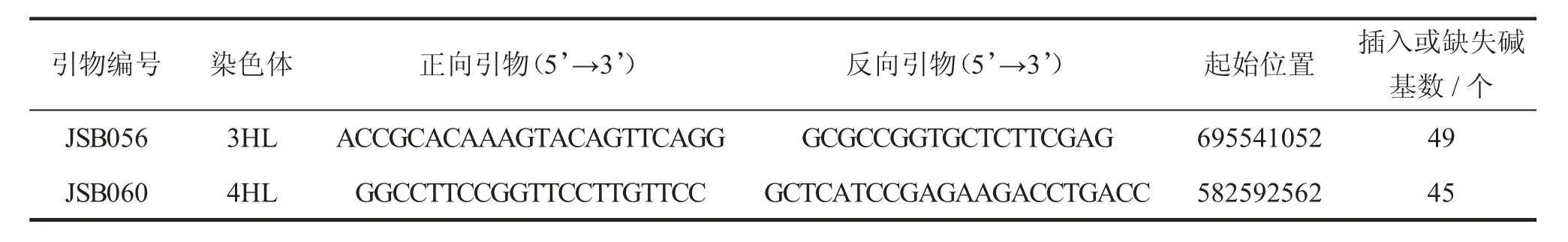
表1 InDel标记信息
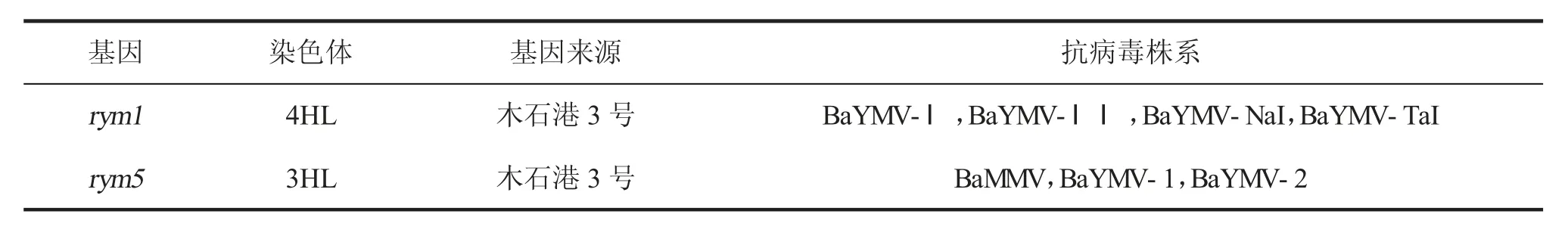
表2 抗大麦黄花叶病基因的基本信息
1.5 自然群体基因型分析
采用CTAB法[11]提取参试材料DNA。利用PCR扩增技术鉴定参试材料基因型。PCR总体系为20 μL,包含1μL上下游引物(浓度为10μmol/L)、2 μL 10×PCR Buffer、13.4μL ddH2O、2μL MgCl2(Mg2+20 mmol/L)、0.4μL DNTPs(浓度为10μmol/L)、0.2 μLTaq酶(浓度为2.5 U/μL)、模板DNA 1μL(浓度为50 ng/μL)。PCR扩增程序为:预变性94℃5 min;变性94℃45 s,退火55℃45 s,延伸72℃45 s,共进行35个循环;再延伸72℃10 min,12℃保存备用。以琼脂糖凝胶(2%)上电泳分离PCR扩增产物,经溴化乙锭(EB)染色,在凝胶成像仪照相、记录,以木石港3号为对照,其带型记为“A”,其余带型记为“B”,进行基因型分型统计。
2 结果与分析
2.1 自然群体的黄花叶病抗性
取每年度4个时期发病病级最高值作为本年度自然群体200份啤酒大麦种质资源黄花叶病抗性鉴定结果,2年数据表明,均表现为抗病的有37份材料,均表现为感病的有143份材料,20份在年份间表现不稳定的材料予以剔除。用于基因型鉴定的啤酒大麦种质资源共180份,啤酒大麦名称及表型抗性信息见表3。
2.2 分子标记鉴定及其准确性
以木石港3号为对照,利用InDel标记JSB056与JSB060对180份啤酒大麦进行基因型鉴定,即与木石港3号带型一致为抗性材料,其余均为感病材料。结果分析表明,JSB056与JSB060的基因型与表现型符合率分别为80.6%和77.8%。由表3可知,聚合2个标记,抗病种质的检测率(检测率=JSB056与JSB060均为抗病基因型材料数/抗病表现型材料数×100%)为91.9%。
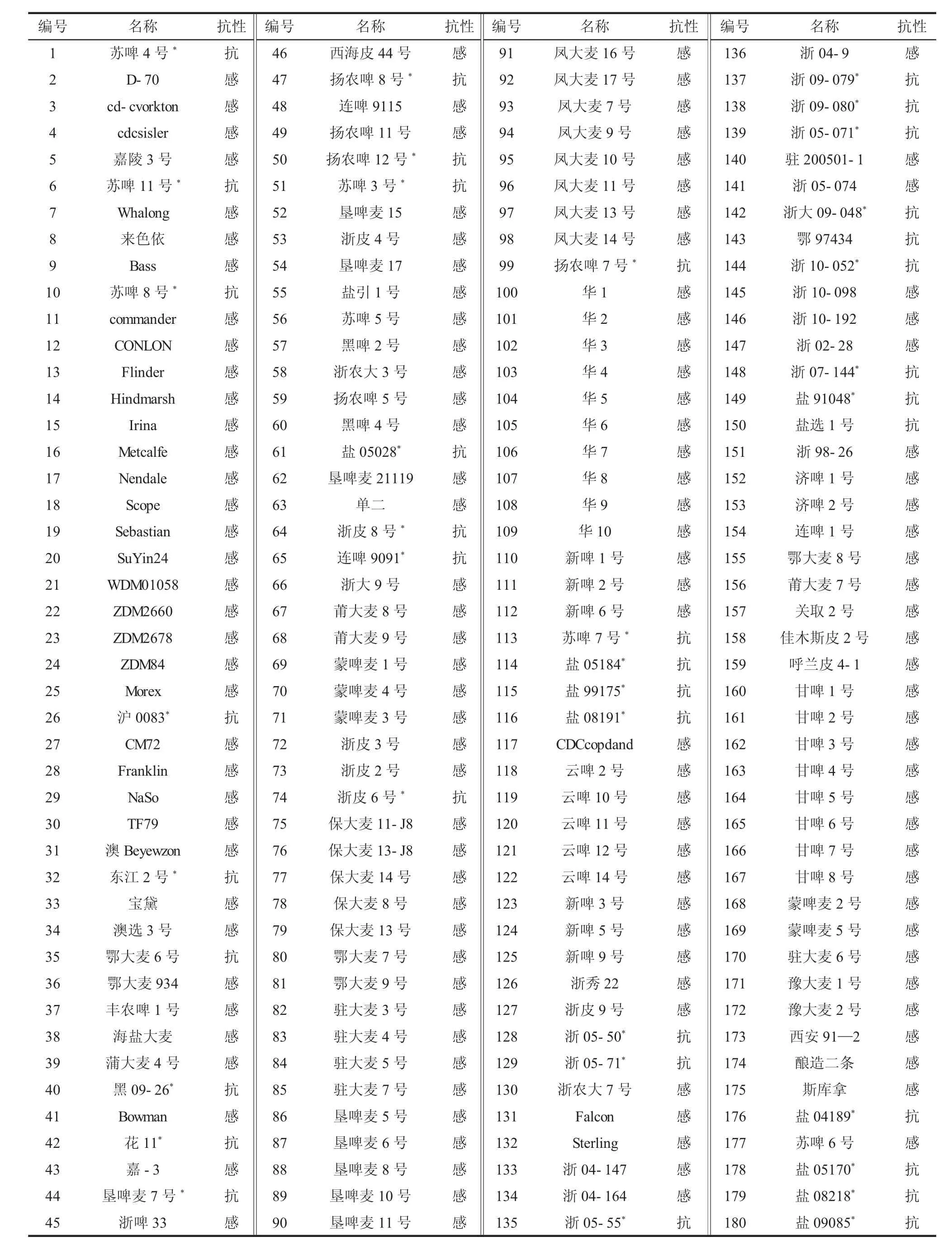
表3 180份啤酒大麦名称及大麦黄花叶病表型抗性信息
JSB056是rym5的连锁分子标记,JSB060是rym1的连锁分子标记。单纯利用JSB056与JSB060均可在自然群体中鉴定大麦黄花叶病抗病材料,但检测效率不高。聚合2个标记,可使检测率高达91.9%。由于rym5与rym1是国内外应用较多的大麦黄花叶病抗性基因,对我国大麦黄花叶病病毒株系具有一定的抗性,因此聚合使用JSB056与JSB060对抗大麦黄花叶病材料检测效果较好,可用于大麦抗黄花叶病的分子辅助选择育种。
标记JSB056的PCR扩增带型见图1,扩增带型清晰度较高,无杂带,进一步说明了InDel标记稳定性强。

图1 InDel标记JSB056在部分啤酒大麦资源中的检测结果
3 讨论与结论
迄今,已报道出21个正式命名的大麦黄花叶病抗性基因,随着大麦黄花叶病病毒生理小种的不断变异,如单二等生产上大面积种植的品种大麦黄花叶病抗性也逐渐丧失。聚合不同抗病基因至同一品种中,提高品种抗性的同时,也能延长其使用年限,是解决生产上抗病问题的有效措施[12]。目前,国内外已获得大量与抗大麦黄花叶病相关的分子标记,除少数克隆基因已经获得共分离标记外,其他分子标记是在特定的遗传背景下基于特定的遗传群体进行定位而获得的,在应用于抗病性育种时需要使用不同遗传背景的分离群体来验证其紧密连锁分子标记的有效性。
DNA分子标记鉴定方法不受环境因素、植物不同发育阶段不平衡等影响。最普遍用于大麦品种鉴定的DNA分子标记为简单重复序列(SSR)和单核苷酸多态性(SNP)。王艳平等利用28个SSR引物对29份大麦资源进行基因型检测,将不同穗棱型品种聚为一类[13]。朱彩梅等利用50个SSR标记对76份糯大麦进行聚类,结果表明种群划分与皮裸性和地理分布有关[14]。张利莎等将SSR标记与EST技术相结合,定性检测了4种大麦麦芽混合后混杂度高于10%的麦芽样品;利用SNP标记与KASP(Kompetitive Allele-Specific PCR)技术相结合,鉴定了混杂度低至5%的麦芽样品[15]。Pattemore等利用45个SNP位点对大麦品种进行基因分型,生成了每个参试材料SNP标记独特的条形码[16]。徐东东等利用SNP标记进行麦芽品质分析,有效地鉴定了品种真实性及麦芽纯度[17]。
SSR与SNP作为最常使用的分子标记,SSR分子标记具有共显性与稳定性的优点,且标记成本低,应用技术简单。SSR标记的缺点是在基因型鉴定过程中会出现带型杂乱、模糊或带型丢失等现象,导致基因型错误判别的概率增加。SNP分子标记分布广泛、数量众多,且是二态性分子标记,其具有检测速度快、自动化水平较高等优点,但由于大麦基因组序列较大,使得开发出的SNP分子标记在利用过程中需要花费大量人力物力,导致成本较高。二态性InDel标记具有共显性、稳定性高与多态性高等优点,通过PCR扩增技术及琼脂糖凝胶电泳即可进行基因分型,且扩增带型清晰度与准确性很高。
rym5与rym1是全球应用较多的大麦黄花叶病抗性基因,本研究表明利用InDel标记鉴定大麦黄花叶病抗性可获得理想的筛选效果。为获得更高的分子标记选择效率,还需进一步定位新的抗病基因,开发出与其紧密连锁的分子标记,从而筛选出聚合多个抗病基因的抗源,培育出抗黄花叶病啤酒大麦新品种。
猜你喜欢
作文小学高年级(2022年5期)2022-06-16 06:22:50
中国农业科技导报(2022年3期)2022-04-26 06:49:52
环境卫生工程(2021年4期)2021-10-13 06:52:26
疯狂英语·新读写(2021年2期)2021-02-25 08:58:46
中国糖料(2020年2期)2020-04-09 11:17:18
河南农业(2019年10期)2019-10-16 08:42:26
河南农业·综合版(2019年10期)2019-09-10 07:22:44
今日农业(2019年12期)2019-08-15 00:56:32
华人时刊(2016年19期)2016-04-05 07:56:10
云南农业(2012年7期)2012-07-30 07:59:12
