
黄河源区不同退化程度高寒草原群落生产力、物种多样性和土壤特性及其关系研究
2021-07-09 09:23:32李成阳张文娟赖炽敏陈小杰尤全刚杜鹤强
生态学报 2021年11期
李成阳, 张文娟, 赖炽敏, 彭 飞, 陈小杰, 薛 娴, 王 涛, 尤全刚, 杜鹤强
1 中国科学院西北生态环境资源研究院, 中国科学院沙漠与沙漠化重点实验室, 兰州 730000 2 中国科学院大学, 北京 100049
黄河源区是我国重要的水源涵养区和生态屏障,高寒草原是该区主要的植被类型之一,该区草地资源丰富,牲畜喜食的牧草种类多,牧草营养价值较高,因此成为青藏高原最主要的畜牧业基地之一[1- 3]。随着近几十年来气候的逐渐干旱化,草地载畜量的迅速增加[4],使该区植被破坏严重、黄沙肆虐、鼠害频发[2]。据报道,该区1/3的草地发生了退化,退化面积约357.13×104hm2,其中严重退化草地面积占退化草地面积的26.79%[5]。在高寒草地退化后,恢复与原始条件相似的生态结构和生物多样性将需要45—60 a的时间[6]。因此,研究黄河源区草地退化机理以及如何恢复高寒草地生态系统具有十分现实的意义。
高寒草地生态系统脆弱,稳定性和抗干扰能力差,植被一旦遭受破坏,恢复难度极大[7]。退化不仅直接影响植被地上生物量和群落结构,而且对土壤的物理和化学性质也会产生影响。在退化的影响下,植被不同功能群在面对土壤环境因子的变化具有不同的适应策略[8],使植被功能群与土壤属性间达到一种反馈平衡。植被群落和土壤因子相互影响、相互作用决定了植物群落结构的发展,并进一步影响植被生产力[9- 10]。土壤有机碳、总氮、速效氮等养分通常被认为是草地生态系统中植物生长的主要限制因子[11]。草地土壤状况直接影响退化高寒草地的恢复效果和能力。在黄河源区草地研究中,大多数研究集中于高寒草甸群落结构和功能对退化的响应[5,12- 13],而对退化高寒草原的研究较为薄弱。在黄河源区退化高寒草原的研究中,周华坤等[3]发现退化后植被盖度、草地质量指数和优良牧草地上生物量比例逐渐下降,草地间的相似性指数减小;王婷等[14]的研究结果表明退化后禾本科功能群丰富度和重要值呈先增后降趋势,杂类草功能群的相对重要值增加,而土壤有机质、全氮和孔隙度呈降低趋势;马世震等[15]发现退化后紫花针茅、早熟禾等优势物种生长势减弱,物候期推迟,群落优势植物逐步演替为以菊科、豆科等杂类草植物为主,物种多样性指数下降。以上研究均从不同的角度分析了群落结构和功能对退化的响应,且均为单点研究,但植被因子和土壤要素之间究竟如何相互作用、相互影响,使生态系统保持稳定或恶化,尚不明确。本研究通过野外调查在黄河源区采用空间退化梯度代替时间退化序列的方法[16],根据植被和土壤特征选取了未退化、轻度退化、中度退化、重度退化和严重退化5个退化阶段,探讨以下3个主要问题:(1) 不同退化程度对植被生物量、不同功能群植物和群落物种多样性的影响;(2) 不同退化程度对土壤特性(土壤水分,容重,土壤有机碳,总氮,铵态氮和硝态氮)的影响;(3) 退化过程中植被要素与土壤因子的相互关系以及影响植被变化的主要土壤因子。
1 材料与方法
1.1 研究区概况
研究区地处黄河源头的青海省果洛藏族自治州玛多县(33°50′—35°40′ N,96°50′—99°20′ E; 图1),青海省南部,平均海拔在4200 m以上。该区属于高原大陆性半湿润气候,气候寒冷,年平均气温-4.1 ℃,降水少且集中,年均降雨量为303.9 mm,其中86%的降水发生在5—6月,年蒸发量为1264 mm[2]。土壤以高山草甸草原土和高山草甸土为主,土壤的平均土层厚度为38 cm,植被遭受破坏,恢复难度较大[2]。该区天然草地面积约2.3万km2,其中可利用草地面积约占总草地面积的78.26%。植被类型主要是高寒草甸和高寒草原,莎草科的小蒿草(Kobresiapygmaea)、矮蒿草(K.humilis)、青藏苔草(Carexmoorcroftii),禾本科的冷地早熟禾(Poacrymophila)和紫花针茅(Stipapurpurea),菊科的沙生风毛菊(Saussureaarenaria)和矮火绒草(Leontopodiumnanum)为常见种。在研究区,生活着大约有475,000个羊单位的大型野生食草动物和家畜,放牧压力指数为1.38,大部分草原处于过度放牧状态[17]。
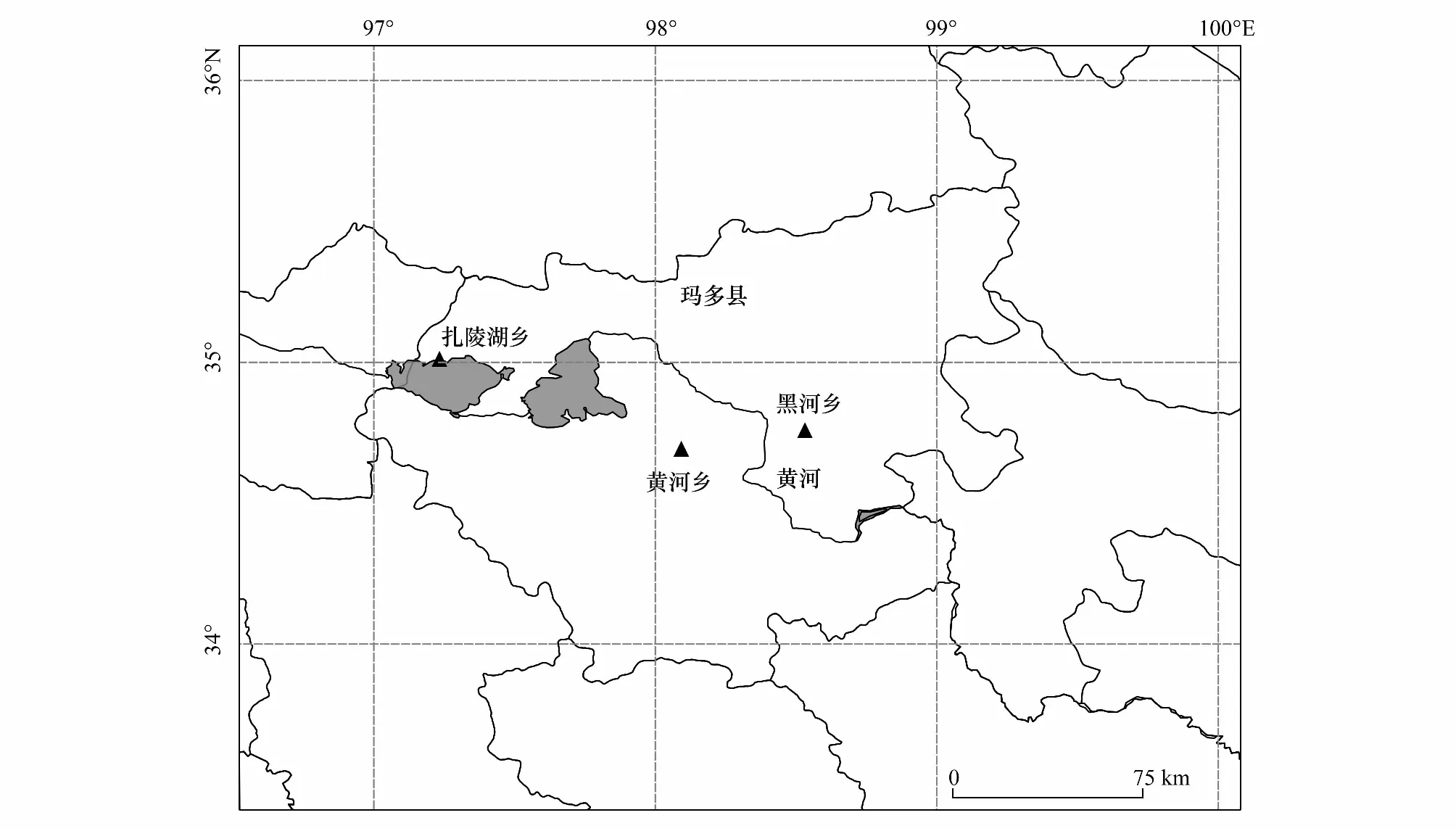
图1 研究区地理位置图以及采样点Fig.1 Location map of the study area and sampling points
1.2 野外调查与取样
于2017年7月在玛多县的扎陵湖乡和黄河乡,2019年7月在玛多县的黑河乡对植被和土壤特征进行野外调查,植被类型为高寒草原。参照Xue等[16]在黄河源和长江源以植被盖度、地表裸露度及莎草科和禾本科所占比重作为草地退化5级梯度标准(表1),采用空间分布代替时间演替的方法来研究植物群落生产力和物种多样性的变化,5个退化演替阶段分别为未退化(CK,intact)、轻度退化(LD,slightly degraded)、中度退化(MD,moderately degraded)、重度退化(SD,severely degraded)和严重退化(VSD,very severely degraded)。于扎陵湖乡、黄河乡和黑河乡分别设置了5个退化演替阶段,每个退化梯度样方面积为10 m×10 m。在每个大样方中随机选择3个30 cm×30 cm的小样方进行植被群落结构调查,调查的内容包括每个物种的高度、盖度和频度。调查完毕,将小样方的地上植被齐地剪下,带回实验室。将地上植被分成禾本科、莎草科和杂类草三大经济类群,放进65 ℃的烘箱中烘48 h,然后取出称重,为不同经济类群的地上生物量。采用土钻法用内径为7 cm的土钻对小样方的地下生物量进行获取,分0—10、10—20、20—30 cm和 30—50 cm 4个土层钻取根系样品[18],将取得的土样带回实验室,根系部分冲洗风干后,根据根的颜色、柔韧性及是否附着毛根辨别出活根,放进65 ℃的烘箱中烘48 h,然后取出称重,因此,本研究中涉及的地下生物量均为活根地下生物量。
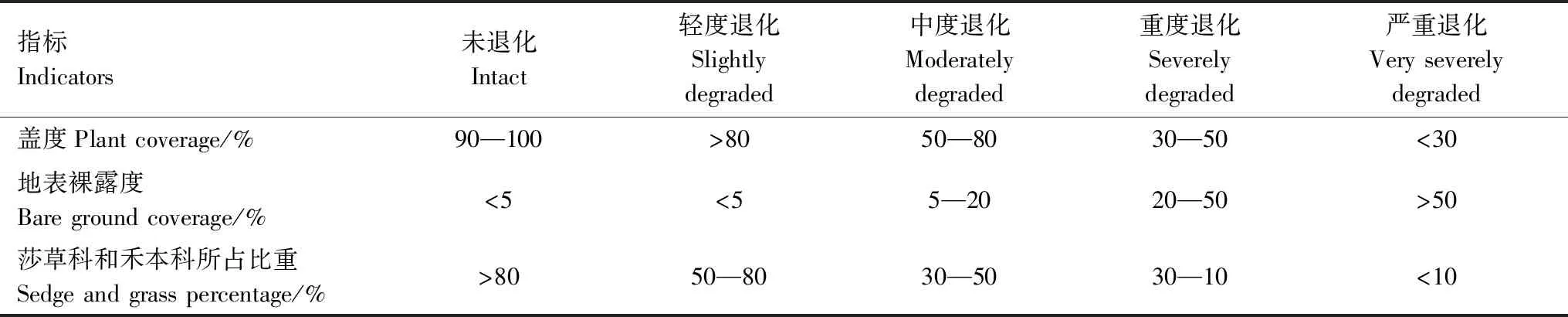
表1 不同退化程度草地划分指标及标准
1.3 土壤理化特征测定
采用环刀法取环刀样用于测定0—10 cm土壤容重(Φ=50 mm)[19];采用TDR- 200水分仪测定0—10 cm土壤水分;另分0—10、10—20、20—30 cm和 30—50 cm四层取土壤样品带回,去除石块、根系等杂物,自然风干,磨碎后过60目筛后,采用重铬酸钾氧化—稀释热法测定土壤有机碳[3];采用重铬酸钾硫酸消化法测定土壤的全氮[3];采用靛酚蓝比色法测定铵态氮[3];采用紫外分光光度法测定硝态氮[3]。
1.4 物种多样性的计算
物种重要值的变化可以反映植物群落组成的变化。
重要值的计算方法:重要值=(相对盖度+相对高度+相对频度)/3[18]
群落物种多样性采用丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度来表示。计算公式如下:
丰富度指数:
S=n
Shannon-Wiener多样性指数:
H=-∑PilnPi
Pielou均匀度指数:
J=H/lnS
式中,Pi为第i种的相对重要值;S为所在样方的物种总数。
1.5 数据处理
本研究中数据分析采用Microsoft Excel 2010和SPSS 23.0软件进行处理,图采用origin 9.0进行绘制。采用双因素方差分析法(Two-way analysis of variance)分析样点(三个不同乡镇)和退化程度及二者的交互作用对地上生物量、丰富度指数、Shannon-Wiener多样性指数、Pielou均匀度指数、土壤容重、土壤水分的影响。采用三因素方差分析法(Three-way analysis of variance)分析样点、退化和土壤深度及三者的交互作用对地下生物量、土壤有机碳、总氮、硝态氮和铵态氮的影响。采用Canoco 5.0软件进行冗余分析法(Redundancy analysis, RDA),揭示植被因子(生物量和物种多样性)和土壤因子(土壤水分、容重、土壤有机碳、总氮、硝态氮和铵态氮)的相互关系。图中和表中每个指标的表现形式均为平均值加减标准误。
2 结果
2.1 生物量
2.1.1地上生物量
草地退化和不同样点对地上生物量均产生了显著影响(P<0.05),但二者的交互作用对地上生物量无显著影响(表2)。随着退化程度的加剧,整体上和各点地上生物量均呈现先稳定后降低的趋势(图2)。整体上,从CK到MD,地上生物量无显著差异,和CK相比,地上生物量在SD和VSD分别降低了36%和60%(图2,P<0.01)。随着退化程度的加剧,扎陵湖乡和黄河乡的变化趋势与整体上一致,黑河乡地上生物量在MD和LD相比显著降低了21%(P<0.05),但和CK相比无显著差异(图2)。随着退化程度的加剧,禾本科地上生物量占总地上生物量比重逐渐降低,莎草科地上生物量比重变化幅度较小,而杂类草比重则逐渐增加(图3)。禾本科地上生物量占总地上生物量比重从CK到LD、MD、SD和VSD分别降低了0.10、0.31、0.59和0.61,杂类草比重则分别增加了0.11、0.36、0.63和0.62(图3)。
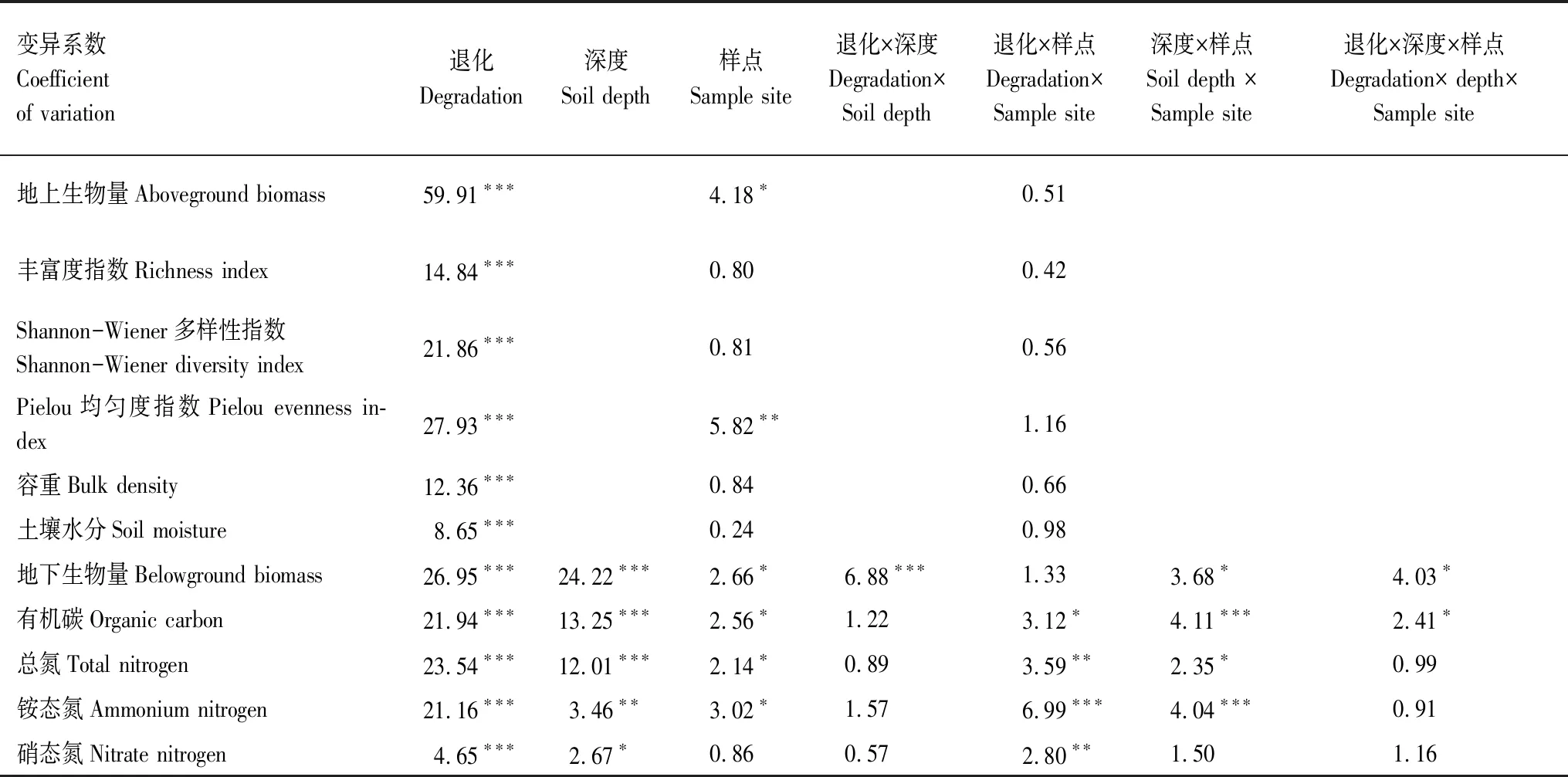
表2 退化高寒草原植被和土壤要素多因素方差分析(F)

图2 不同退化程度下整体上和各样点地上生物量Fig.2 Aboveground biomass at overall and each sample sites under different degraded stages 不同小写字母代表不同退化程度各指标差异显著(P<0.05);CK:未退化 intact; LD:轻度退化 Slightly degraded; MD:中度退化 moderately degraded; SD:重度退化 Severely degraded; VSD:严重退化 Very severely degraded
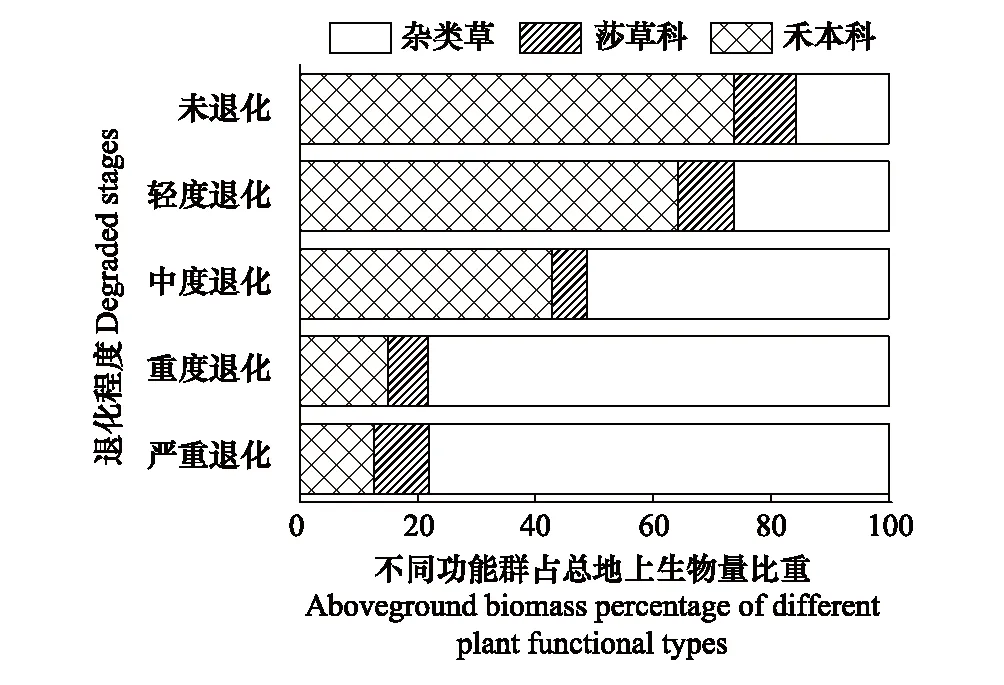
图3 不同退化程度下各功能群占总地上生物量比重 Fig.3 Aboveground biomass percentages of different plant functional types under different degraded stages
2.1.2地下生物量
草地退化程度,土壤深度和不同样点以及两两之间(除了草地退化程度和样点交互作用)和三者之间交互作用均对地下生物量产生了显著影响(表2,P<0.05)。与地上生物量的变化趋势相似,随着退化程度加剧,0—50 cm地下生物量呈现先稳定后降低的趋势(图4)。在不同样点上,0—50 cm地下生物量在CK阶段,黄河乡(1495g/m2)>黑河乡(1320 g/m2)>扎陵湖乡(1130 g/m2)。整体上,从CK到MD,地上生物量无显著差异,和CK相比,0—50 cm地下生物量在SD和VSD分别降低了44%和66%(图4,P<0.001);随着退化程度的加剧,0—50 cm地下生物量在扎陵湖乡、黑河乡和黄河乡与整体上变化趋势类似,但扎陵湖乡和黄河乡在MD和CK相比显著降低(图4,P<0.05)。随着退化程度的加剧,整体和各样点上,0—10 cm地下生物量与0—50 cm地下生物量变化趋势一致;10—20 cm地下生物量在LD和MD显著增加,随后降低;20—30 cm地下生物量在LD和MD保持稳定随后降低;整体上和扎陵湖乡,30—50 cm地下生物量无显著变化,黑河乡和黄河乡在VSD显著降低(图4,P<0.05)。随着退化程度的加剧,浅根系植物逐渐减少,而深根系植物则逐渐增加。
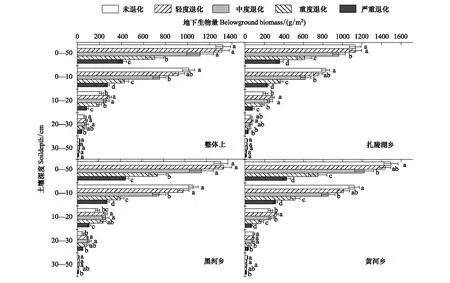
图4 不同退化程度下整体上和各样点地下生物量及各层分布Fig.4 Belowground biomass and its distribute in different layers at overall and each sample site under different degraded stages
2.2 物种多样性
草地退化程度对物种丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数均产生了显著影响(P<0.001),不同样点对Pielou均匀度指数具有显著影响(表2,P=0.003)。在整体上,丰富度指数从CK到LD和MD无显著变化,SD和VSD与CK相比显著降低了27%和31%(P=0.045和P=0.034);和CK相比,Shannon-Wiener多样性指数在LD和MD显著增加了20%和15%(P=0.025和P=0.039),在SD和VSD显著降低了20%和39%(P=0.021和P=0.007);Pielou均匀度指数从CK到SD均无显著变化,和CK相比,VSD显著降低了22%(图5,P=0.018)。随着退化程度的加剧,扎陵湖乡和黄河乡Pielou均匀度指数变化趋势和整体上一致,但黑河乡在SD出现显著降低(图5,P=0.041)。

图5 不同退化程度下整体上和各样点物种多样性指数Fig.5 Species diversity index at overall and each sample site under different degraded stages
2.3 土壤物理和养分特征
草地退化程度对土壤水分和容重具有显著影响(表2,P<0.001)。随着退化程度的加剧,土壤水分呈现降低趋势,和CK相比,LD和MD无显著变化,SD和VSD分别显著降低了31%和59%(图6,P=0.029和P<0.001)。土壤容重随着退化程度的加剧呈增加趋势,和CK相比,MD、SD和VSD分别显著增加了16%、14%和30%(图6,P<0.05)。
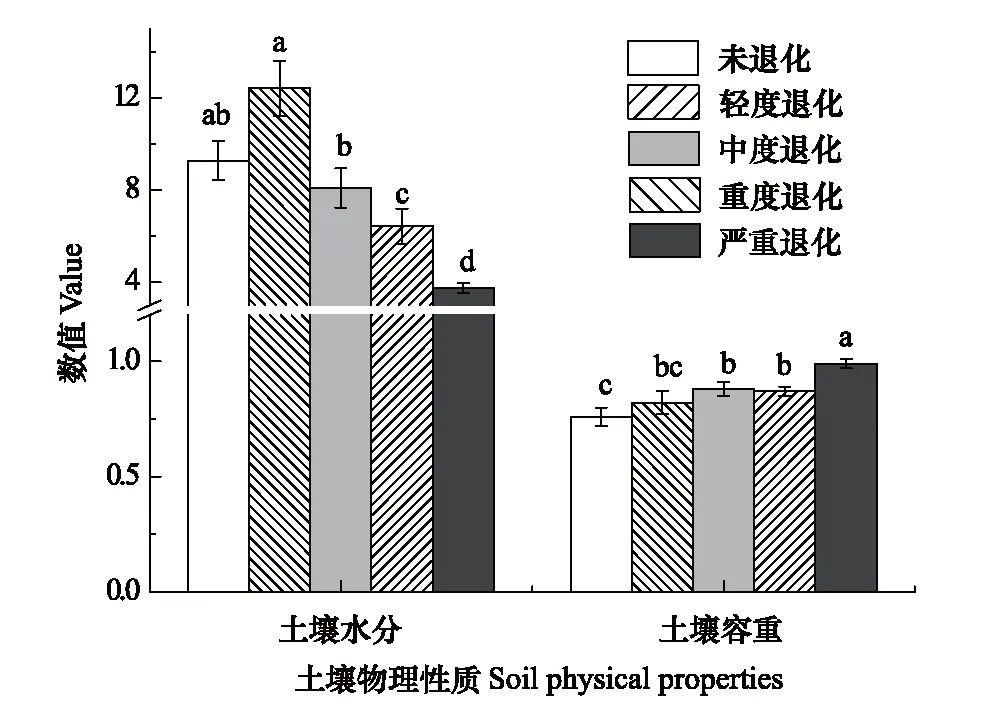
图6 不同退化程度下0—10 cm土壤深度土壤水分(%)和容重(g/m3)Fig.6 Soil moisture and bulk density under different degraded stages at 0—10 cm depth
草地退化程度,土壤深度和不同样点两两之间(除了退化程度和深度的交互作用)和三者之间的交互作用均对土壤有机碳产生了显著影响(表2,P<0.05);草地退化程度,土壤深度和不同样点两两之间(除了退化和深度的交互作用)均对土壤总氮和铵态氮产生了显著影响(表2,P<0.05);草地退化程度,土壤深度以及草地退化和样点的交互作用均对硝态氮产生了显著影响(表2,P<0.05)。随着退化程度的加剧,无论是整体上还是各样点,土壤有机碳、总氮、铵态氮和硝态氮在0—10、10—20、20—30 cm和 30—50 cm均呈现降低趋势(图7), 其过程大致可以划分为两个层次。第一个层次为CK到MD阶段,土壤有机碳、总氮和铵态氮相对SD和VSD阶段含量较高,无论是整体上还是各样点在LD阶段0—10 cm土壤有机碳和总氮均有一定降低,而10—20 cm土壤有机碳和总氮则有一定增加;第二个层次为SD到VSD阶段,土壤有机碳、总氮和铵态氮显著降低,并在VSD达到最低值(图7)。硝态氮随着退化程度的加剧表现土壤有机碳、总氮和铵态氮略有不同,但总体趋势一致。如在整体和各样点上,0—10 cm和10—20 cm硝态氮在MD显著降低(图7,P<0.05)。

图7 不同退化程度下整体上和各样点0—50 cm土壤深度土壤养分特征Fig.7 Soil nutrients in 0—50 cm at overall and each sample site under different degraded stages
2.4 退化高寒草原植被要素与土壤因子的关系
基于Canoco 5.0软件分析,趋势对应分析(DCA)结果显示,第一轴的梯度范围是1.6,这表明植被要素和土壤因子之间适应于多元统计RDA分析。RDA分析表明:第一、二排序轴累计解释率分别为61.8%和3.0%(图8),说明第一、二排序轴能够较好的反映植被要素和土壤环境因子之间的关系,并且第一排序轴起了决定性作用。地上生物量、地下生物量、丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数与土壤水分、有机碳、总氮、铵态氮和硝态氮呈正相关关系,与土壤容重呈负相关关系(图8)。土壤因子对驱动植被要素变化的贡献由大到小分别为:土壤容重,硝态氮,有机碳,铵态氮,总氮和土壤水分(表3)。其中,土壤容重(P=0.001),硝态氮(P=0.024),有机碳(P=0.093)对植被要素变化具有显著影响。
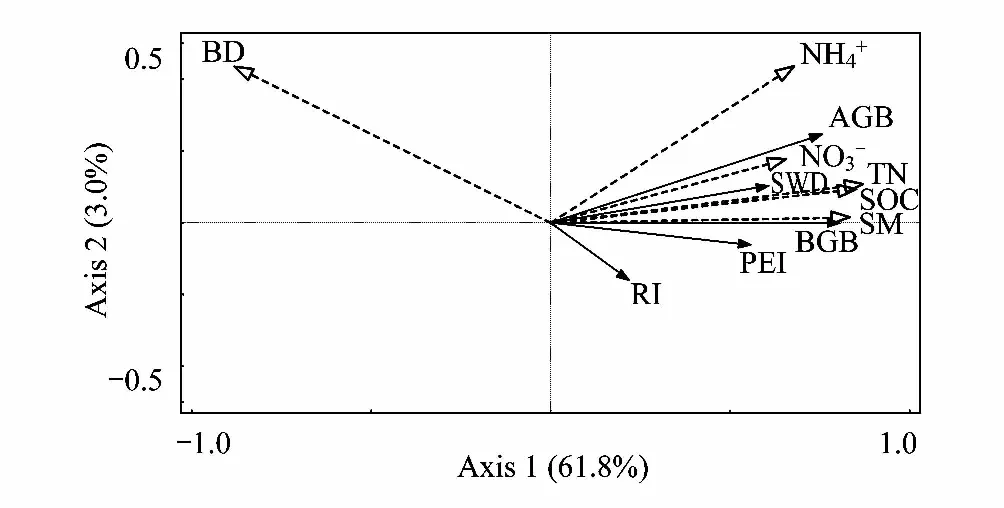
图8 植被要素和土壤环境因子的RDA分析 Fig.8 Redundancy analysis of vegetation elements and soil environmental factorsAGB:地上生物量 Aboveground biomass; BGB:地下生物量 Belowground biomass; RI:丰富度指数 Richness index; SWD: Shannon-Wiener多样性指数 Shannon-Wiener diversity index; PEI:Pielou均匀度指数 Pielou evenness index; BD:容重 Bulk density;SM:土壤水分 Soil moisture; SOC: 土壤有机碳 Soil organic carbon; TN:总氮 Total 铵态氮 Ammonium nitrogen; 硝态氮 Nitrate nitrogen
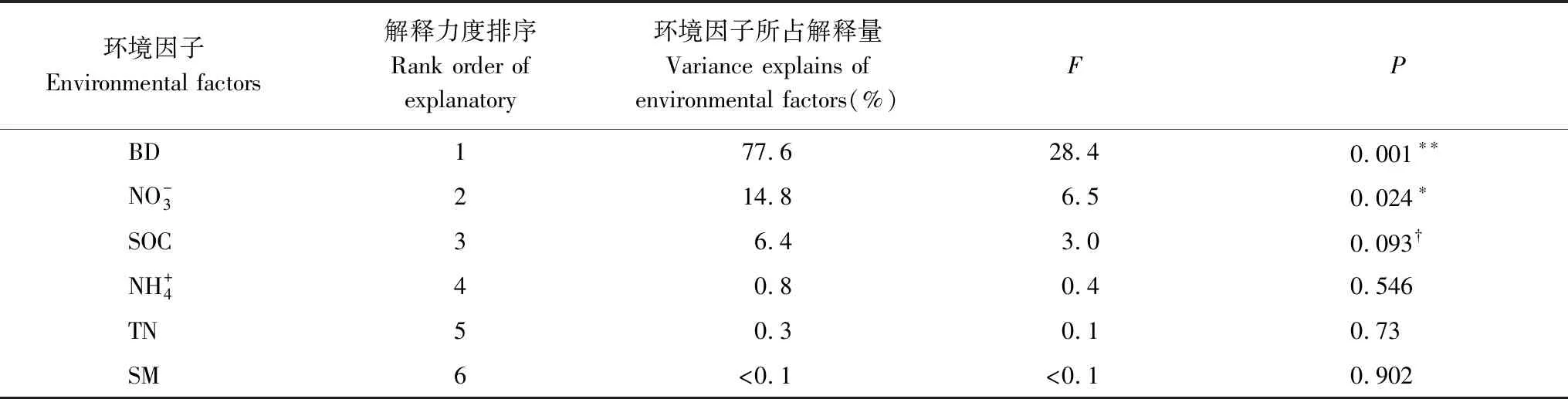
表3 退化高寒草地环境解释力度和显著性检验结果
3 讨论
高寒草地的退化主要表现在三个方面:一是植被的退化,如群落生产力降低,群落结构恶化[20];二是土壤的退化,如土壤持水性能和碳氮等土壤养分的下降,砂粒含量增加[21];三是植被和土壤系统耦合关系的丧失和系统相悖所致[22]。在本研究中,随着退化程度的加剧,整体上和各样点的地上和地下生物量在总体上均呈下降趋势,这与藏北退化高寒草地[23]的变化趋势一致。但在轻度和中度退化阶段,生物量无显著变化,甚至在轻度退化阶段地上和地下生物量均有不显著增加,这与青藏苔草草原[24]有所不同,可能和本研究以盖度等指标,而青藏苔草草原以优势种频度等指标作为划分依据以及草地类型的优势种生活型特征不一致有关。退化后杂类草所占比重越来越大,杂类草往往具有较高的地上和地下生物量[25],弥补了轻度和中度退化后禾本科植被盖度和生物量的降低。研究区放牧压力较大,牲畜一般喜食禾本科和莎草科植物,间接增加了杂类草的竞争能力[26],同时杂类草根系较深,在环境变恶劣时,可以更好的利用深层的土壤水分和养分[25],因此,在退化过程中,杂类草能更好的适应环境的变化,群落优势种由禾本科和莎草科植物向杂类草植物转变。物种丰富度指数、多样性指数和均匀度指数均随退化程度的增加而降低,和高寒草甸在退化过程中的演替规律相一致[10]。但在植被退化演替过程中具体的某些阶段,物种多样性可能会增加。如轻度和中度退化阶段,由于禾本科和莎草科植物减少,降低了群落的高度,使处于群落下层的一些低矮的杂类草植物光照条件变好[27],为处于竞争劣势的杂类草植物的入侵提供了机会[24],导致本研究中Shannon-Wiener多样性指数在轻度和中度退化阶段增加。各指数最小值均出现在严重退化阶段,过度放牧导致群落盖度急剧下降,群落物种数量和物种个体分配不均匀可能是导致不同退化阶段生物多样性差异的主要原因[28]。
黄河源区草地土壤粗骨性强,成土过程相当缓慢,生态系统一旦遭受破坏,恢复难度极大[2]。一般来说土壤性质的变化与植物群落结构特征息息相关。在本研究中退化后土壤水分和土壤养分在整体上与植被盖度、生物量等变化趋势一致,表现出整体上变化的同步性,这与很多学者的研究结果一致[3, 23]。但就具体的退化阶段而言,土壤性质可能表现出与植物群落结构特征变化的同步性,也有可能是异步性。如在轻度退化阶段,土壤养分表现出和地下生物量变化的同步性。和未退化草地相比,0—10 cm土壤有机碳和全氮有一定降低,10—20 cm土壤有机碳和全氮则有一定增加,土壤中有机碳、全氮主要来源于凋落物和植物根系的分解[11],本研究中0—10 cm地下生物量在轻度退化阶段有一定降低,而10—20 cm地下生物量在轻度退化阶段则有一定增加可能是土壤中有机碳和全氮变化的主要原因。重度和严重退化阶段,地上和地下生物量急剧下降,导致土壤中有机碳和全氮降低。各层铵态氮和硝态氮在重度和严重退化阶段均显著降低,这可能和重度和严重退化阶段土壤侵蚀加强,土壤有机质流失严重,土壤氮存量的下降有关[29]。但在中度退化阶段,土壤性质和植物生物量的变化却表现出异步性。土壤水分显著降低,而土壤容重则显著增加,土壤有机碳和全氮有一定的降低,但生物量却无显著变化。这可能是由于中度退化阶段植被盖度下降幅度较大(表1),草毡层剥落、变薄,减少了植被对降水的截留与缓冲,增加了降水对裸地的溅蚀[19],导致了土壤持水能力下降,同时降低了土壤容重的垂向异质性,使表层土壤容重显著增加[30],土壤养分流失。此外,杂类草在中度退化阶段已经逐渐成为群落的优势物种,杂类草往往生物量较高,且多为轴根系[24],持水能力低下。因此,在中度退化阶段,尽管生物量无显著变化,但盖度和群落结构的变化也能导致土壤物理化学性质发生明显变化。
在本研究中,从整个退化过程来看,植被生物量和物种多样性与土壤养分正相关,与土壤容重负相关。在轻度和中度退化阶段,牲畜的选择性采食导致占据群落上层的禾本科植物高度降低[31],遮阴效果减弱,杂类草接受光照的机会增加,大量杂类草入侵,尤其是菊科植物,提高了群落生物量,同时了增加了物种多样性[24],而物种多样性的增加导致土壤碳输入和氮矿化速率增加[32],有机化合物更加多样化,微生物分解加强,产生更多的细胞外酶[21],从而提高了土壤养分含量,进而增加群落的生产力。在重度和严重退化阶段,植被生物量和物种多样性急剧下降,土壤养分的来源降低[11],此时土壤养分含量随之下降。牲畜对草地土壤长期践踏以及水土侵蚀造成小粒径土壤颗粒流失会使得土壤容重增加[33-34],土壤碳和氮储存能力下降[9],进而抑制植物生长,降低生物量和物种多样性,因此,植被生物量和物种多样性与土壤养分正相关,与土壤容重负相关。在具体的退化阶段,土壤性质与植物群落结构特征的变化可能表现出异步性,植物群落结构特征与土壤性质的关系也可能会改变,因此,在对退化高寒草原进行恢复时,应多关注土壤性质与植物群落结构特征变化的异步性。
4 结论
随着黄河源区高寒草原的退化,群落地上、地下生物量和物种多样性,土壤水分、有机碳和全氮均呈先稳定后降低的变化规律,说明适度的放牧有利于植物的生长和土壤养分的增加,促进草地生态系统的良性发展。土壤容重随着退化程度的加剧而显著增加。到了严重退化阶段,土壤水分、有机碳和全氮降到了最低,已经不能满足植物生长的需求。植被生物量和物种多样性与土壤养分正相关,与土壤容重负相关。在具体的退化阶段,如中度退化,土壤性质与植物群落结构特征的变化可能表现出异步性,因此,针对不同退化阶段应采取不同的恢复治理措施,同时对中度和重度退化两个关键阶段应该给予更多的关注。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
今日农业(2020年19期)2020-12-14 14:16:52
浙江农业学报(2016年7期)2016-06-15 20:30:08
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:16
现代农业(2016年4期)2016-02-28 18:42:07
中国科技信息(2015年2期)2015-11-16 08:18:32
环境科技(2015年2期)2015-11-08 12:11:30
水生生物学报(2015年1期)2015-02-28 16:01:05
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
