
水淹胁迫诱导喜旱莲子草茎发生类环割效应
2021-07-09 05:07:16任心怡阿依巧丽张小萍
生态学报 2021年11期
邹 曼,任心怡,林 锋,阿依巧丽,曾 波,张小萍
三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室,西南大学生命科学学院, 重庆 400715
在全球气候变化的大背景下,强降雨天气频繁出现,洪涝灾害也时有发生,越来越多的陆生植物面临不同程度的水淹胁迫[1- 2]。在植物遭受的水淹胁迫中,以植株的部分结构或器官遭受水淹(即局部水淹)最为常见。喜旱莲子草(Alternantheraphiloxeroides(Mart.) Griseb.)是在我国有广泛分布的一种苋科植物,具有较强的水淹耐受能力。野外观察发现,当喜旱莲子草遭受局部水淹时,位于水面上的未受淹的茎段与位于水面下的受淹茎段相比,常会表现出加粗膨大的现象。
植物光合作用生产的碳水化合物是植物赖以生长和存活的物质基础。光合器官(主要是叶)生产的光合产物会被运输到所有需要碳水化合物的组织和器官中,根据植物的需求用于生长、代谢利用、或合成贮藏化合物[3- 6]。因此,碳水化合物在植物体内的运输是基础而重要的生理过程。碳水化合物的运输需要畅通的运输通道,如果运输通道被阻断而不畅通或者阻塞,碳水化合物就无法运输到需要碳水化合物的组织和器官中从而在运输通道被阻断处或者阻塞处产生积累[7- 8]。我们所熟知的环割效应就是植物主干或枝在被环割损伤韧皮部(即运输通道)后导致环割位置处产生瘤状物而致环割位置处的形态学上端的主干或枝发生膨大的现象[9]。由于环割导致的碳水化合物积累可以促进花芽分化及果实和种子的产量,环割在园林园艺生产上有广泛应用[10- 14]。
已有不少研究表明碳水化合物的运输是一主动运输过程,在运输中需要耗能才能使运输顺利进行[15- 17]。可以想象,如果在运输中能量供应不足,碳水化合物的运输将受阻从而使碳水化合物发生积累;这种由于能量供应不足导致的碳水化合物运输受阻而发生积累致使植株主干或枝膨大的现象与植物由于环割损伤导致的碳水化合物运输受阻而发生积累使植株主干或枝膨大的现象类似,在外观表现上是相同的。我们把这种不是由于物理损伤而是因能量供应不足导致碳水化合物运输受阻积累而使植株主干或枝膨大的现象称为类环割效应。
在降雨、洪汛、潮汛、水坝修建等引发的水淹发生时,除了植物被完全淹没于水下外,常见的更多的情况是植物被局部淹没,即位于土壤基质中的根以及植株的下部组织和器官被淹没于水中,而植株上部的组织和器官仍位于水面上。植物遭受局部淹没时,由于氧在水中的低溶解度和低扩散能力,水体内氧含量远低于空气中的氧含量[18- 20],假如植物没有发达的气体运输通道能够通过其位于水面上的组织器官把氧气从空气中有效地运输到位于水面下的组织和器官中,则被水淹没的组织和器官将面临氧气缺乏从而发生有氧呼吸减弱能量供应不足的问题。在此情况下,我们可以推测,当局部淹没的植物其水上部分光合组织生产的光合产物向水面下的根系及其他需要光合产物的组织器官进行输送时,由于被淹组织内能量供应不足,光合产物向被淹组织中的运输会受限从而在植物位于水面上的未受淹组织中发生积累而引发类环割效应。
为了明确在遭受局部水淹时喜旱莲子草未受淹的茎段是否确实会发生增粗膨大以及茎段中的碳水化合物是否会发生积累,以探究遭受局部水淹的喜旱莲子草茎的膨大与茎中碳水化合物积累的关系,我们对喜旱莲子草在遭受局部水淹时植株体内的碳水化合物状况和茎形态变化进行了研究。
1 材料与方法
1.1 研究材料
喜旱莲子草又名空心莲子草,是苋科莲子草属的多年生草本植物。喜旱莲子草叶对生,茎由节和节间组成,节实心无孔隙孔道等通气组织,节间具有中空的髓腔。喜旱莲子草原产于南美洲,在我国分布广泛,水陆生境均可生长[21]。该物种耐淹能力强,在遭受长时间水淹后存活率仍非常高[22],是非常好的用于研究植物水淹胁迫响应和水淹耐受机制的植物物种。
1.2 植株培养与实验处理
2015年7月,在重庆北碚嘉陵江河岸带自然生长的喜旱莲子草克隆种群中,选择健壮、生长良好、茎粗细相近的植株,剪取长约20 cm具有完整顶梢的茎段做为扦插苗,带回西南大学三峡库区生态环境教育部重点实验室实验研究基地后,用混合均匀的黄壤和腐殖土(比例为1∶1)作为栽培基质,将扦插苗栽植于装满栽培基质的花盆中(盆高10 cm,内径12 cm),每盆栽植一株,每株扦插苗茎基部的两个节埋入土中以使其生根。所有植株均放置于相同的环境中进行培养,培养期间接受自然光照并进行浇水、除草等日常管理,保证培养的植株具有良好的生长条件。一个月后,待植株培养生长到合适大小后,选择生长健壮、茎长一致、具有的叶片数量相同且叶片大小尽量一致的植株用于实验处理(表1),所有的植株均由茎、叶、地下部分构成,茎上不具有侧枝。

表1 实验处理前喜旱莲子草植株的茎长、茎基径、茎节间数量
本研究设置2个实验处理组,把上述选择好的植株随机分配到这两个实验处理组中,每组20株。第一个处理组为水淹处理组,将该组的植株(连同栽种植株的花盆)悬吊在装满自来水的水池中(水池长3 m、宽2.5 m、水深2 m),每株植株使其茎的一半淹没于水中(茎被淹没的节间数约为7个),一半露出于水面上,并用红色丝线标记茎被水淹没的位置,保证在整个实验期间水池中的水位不变同时植株茎被淹的位置不变。第二个处理组为不进行水淹的对照组,同样在植株茎一半的位置(相当于水淹处理组植株茎被水淹没的位置)用红色丝线进行标记。两个处理组中的所有植株具有的叶的数量相同、叶片大小近乎一致,且所有的叶均位于茎的远端段上(即红色丝线标记处以远的茎段上)。对照组植株放置于水池旁的旷地上,使其与水淹处理株具有相同的光照和温度条件,在整个实验期间对对照组植株进行浇水管理保证其土壤具有合适的水分含量。在实验期间,使水淹处理组所在水池中的水处于缓慢流动和缓慢连续换水的状态,水池中的水pH值保持在7.6—7.9之间,溶氧含量保持为饱和溶氧含量的95%—100%;对水淹处理组和对照组植株均进行防虫、除草等常规管理。
1.3 植株测量与碳水化合物测定
2015年12月,实验处理结束后,所有实验植株均由茎、叶、地下部分构成,茎上无侧枝产生;对所有实验植株进行测定。测量水淹处理组和对照组各植株的茎各节间的直径,并切取水淹处理株水面上未淹茎段中膨大程度最大的节间的横切切片和对照株对应节间的横切切片,用体视显微镜(SMZ25, 日本尼康)观察并测定节间的茎壁厚度(NIS-elements Imaging Software, version 4.30)。测量结束后,取植株茎红色丝线标记位置以上的茎段(对水淹处理株即水面上的未受淹茎段)和成熟全展叶,在80 ℃下烘干至恒重后分别用球磨仪(WS-MM400,德国)研磨成粉末,测定茎和叶中的非结构性碳水化合物含量。
植物体内的非结构性碳水化合物主要由单糖、低聚糖、多糖构成。在本研究中非结构性碳水化合物的测定采用在传统的蒽酮硫酸法[23]的基础上改进后的方法进行。先在80 ℃水浴中用80%乙醇浸提茎或叶粉末样品中的单糖及低聚糖,80%乙醇浸提后的样品残渣在80 ℃水浴中用纯水浸提水溶性多糖[24],经纯水浸提后的样品残渣加入2 mol/L盐酸在60 ℃水解后用于检测不可溶性多糖。上述操作所得提取液(待测液)用蒽酮试剂测定各类糖的具体含量,这三类糖含量的总和即为样品中非结构性碳水化合物的含量。
1.4 数据分析
为了检测水淹植株位于水面上的未受淹茎段的节间相比水面下的被淹茎节间是否发生了膨大,以植株被淹没茎段(未水淹对照处理株相应地为红色丝线标记处以下的茎段)所有节间直径的平均值为参照(以后简称参照直径),对植株未受淹茎段(对照处理株相应地为红色丝线标记处以上的茎段)的以下指标进行测定:
膨大节间数(个):未淹茎段上直径比参照直径大的节间数量;
膨大节间比例(%):未淹茎段所有节间中膨大节间占有的百分比;
最大膨大量(cm):未淹茎段所有节间中膨大程度最大的节间的直径与参照直径的差值;
平均膨大量(cm):未淹茎段所有节间直径与参照直径差值的平均值。
采用独立样本T检验检测水淹植株与未水淹对照植株的茎膨大状况、最大膨大程度节间茎壁厚、非结构性碳水化合物含量是否有差异。数据分析使用SPSS 22.0完成,作图使用Origin 9.0完成。
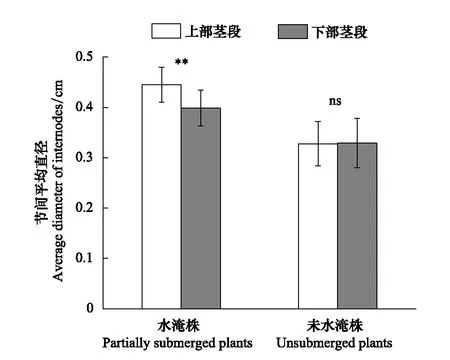
图1 水淹喜旱莲子草植株位于水面上的茎段与水面下茎段和未水淹植株对应的上部茎段与下部茎段的节间平均直径(平均值±标准差)Fig.1 The average diameter (means ± s.d.) of internodes on the abovewater and belowwater stem parts of partially submerged Alternanthera philoxeroides plants and the counterpart stem parts of unsubmerged plants **表示不同处理之间有极显著差异(P< 0.01);ns表示不同处理之间没有显著差异(P >0.05)
2 结果
2.1 水淹和未水淹喜旱莲子草植株的茎形态
研究结果表明,喜旱莲子草植株遭受水淹后,其位于水面上的未受淹茎段与水面下的受淹茎段相比发生了明显的膨大现象,未受淹茎段的节间平均直径明显大于受淹茎段节间平均直径(P<0.01,图1);但是,对未水淹喜旱莲子草植株而言,其上部茎段(相当于水淹植株的未受淹茎段)和下部茎段(相当于水淹植株的受淹茎段)节间平均直径并无差异(P>0.05,图1),上部茎段也无明显膨大现象发生。与未水淹植株相比,水淹植株水面上的未受淹茎段发生膨大的节间数显著增多(P<0.05,图2)。另外,实验结果表明,水淹植株位于水面上的未淹茎段上发生膨大的节间数量占未淹茎段上所有节间数量的百分比显著高于未水淹植株,其值高达75%(P<0.01,图2)。

图2 水淹喜旱莲子草植株位于水面上的茎段和未水淹植株对应茎段上发生膨大的节间数量及百分比(平均值±标准差)Fig.2 Number and proportion (means ± s.d.) of diameter-widened internodes on the abovewater stem parts of partially submerged Alternanthera philoxeroides plants and the counterpart stem parts of unsubmerged plants*表示不同处理之间有显著差异(P< 0.05);**表示不同处理之间有极显著差异(P< 0.01)
研究结果表明,水淹不仅增加了喜旱莲子草位于水面上的未淹茎段上的膨大节间数量,也显著增大了未淹茎段上节间的膨大程度。与未水淹植株相比,水淹植株未淹茎段的节间最大膨大量显著增强(P<0.01),节间直径最大膨大量约为未水淹植株对应茎段的节间直径最大膨大量的2倍(图3)。比较水淹植株水面上未淹茎段所有节间与未水淹植株对应茎段所有节间的膨大量平均值,发现两者也有极显著差异(P<0.01);水淹植株未淹茎段所有节间平均膨大量(即节间直径增粗量平均值)达到0.05 cm,而未淹植株对应茎段的平均膨大量小于0,这说明水淹植株位于水面上的茎段相对于水面下的茎段在膨大加粗,而未淹植株主茎上部茎段直径并不比下部茎段的直径大,相反显得要更小(图3)。此外,研究结果表明,水淹植株水面上未淹茎段中具有最大膨大程度的节间的茎壁厚显著大于未淹植株对应节间的茎壁厚,二者之间具有极显著差异(P<0.001)(图4)。
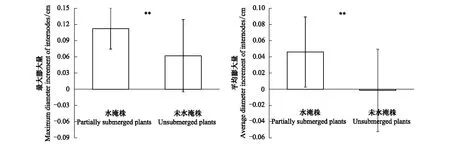
图3 水淹喜旱莲子草植株位于水面上的茎段和未水淹植株对应茎段上节间的最大膨大量和平均膨大量 (平均值±标准差)Fig.3 The maximum and average diameter increment (means±s.d.) of internodes on the abovewater stem parts of partially submerged Alternanthera philoxeroides plants and the counterpart stem parts of unsubmerged plants**表示不同处理之间有极显著差异(P< 0.01)

图4 水淹喜旱莲子草植株位于水面上茎段中具最大膨大程度的节间和未水淹植株对应节间的茎壁厚(平均值±标准差)Fig.4 The stem wall thickness (means±s.d.) of the widest internodes on the abovewater stem parts of partially submerged Alternanthera philoxeroides plants and the counterpart internodes of unsubmerged plants***表示不同处理之间有极显著差异(P< 0.001)
2.2 水淹和未水淹喜旱莲子草植株茎、叶非结构性碳水化合物含量
对水淹喜旱莲子草植株位于水面上的未淹茎段和未水淹喜旱莲子草植株对应的茎段(即红色丝线标记位置以上的茎段)中的非结构性碳水化合物含量检测发现:醇溶性单糖及低聚糖含量在水淹植株与未水淹(对照)植株之间无显著差异(P>0.05);不可溶性糖含量在二者之间有显著差异(P<0.05),分别为13 mg/g和17 mg/g;水溶性多糖在喜旱莲子草茎中含量较高,水淹植株和未水淹植株中的含量分别为557 mg/g和432 mg/g,水淹植株中的含量显著高于未水淹植株(P<0.01,图5)。就非结构性碳水化合物来看,水淹植株位于水面上的未淹茎段的非结构性碳水化合物含量显著高于未水淹对照植株对应茎段(P<0.01)。
对水淹喜旱莲子草植株位于水面上的未淹茎段上的叶(位于水面下的水淹茎段上没有叶存在)和未受淹喜旱莲子草植株对应茎段上的叶的糖含量检测发现:叶中的醇溶性单糖及低聚糖含量在水淹处理株和未水淹对照株之间无显著差异(P>0.05),叶中的不可溶性糖在水淹处理株和未水淹对照株之间也无显著差异(P>0.05),但水淹处理株叶中的水溶性多糖含量显著高于未水淹对照株,达到未淹对照株的6.5倍之多(P<0.001,图5)。

图5 水淹喜旱莲子草植株位于水面上的茎段和叶和未水淹植株对应茎段及其上的叶的非结构性碳水化合物含量(平均值±标准差)Fig.5 Non-structural carbohydrates content (means±s.d.) in the abovewater unsubmerged stem parts and leaves of partially submerged Alternanthera philoxeroides plants and the counterpart stem parts and leaves of unsubmerged plants对每一类型的糖,不同字母表示其含量在不同处理之间有显著差异(P < 0.05)
3 讨论
植物茎的生长包括从形态学下端向形态学上端的伸长生长和茎直径增加的增粗生长。植物的茎或枝在向形态学上端伸长生长的过程中,已长出的茎或枝同时也在进行着增粗生长。虽然双子叶植物和单子叶植物茎增粗生长的方式和过程不同,但毫无疑问,两者的茎形态学下端开始增粗生长的时间都要早于茎形态学上端,形态学下端生长时间相对较长,从而都表现出茎形态学下端比茎形态学上端粗的形态特征[25]。观察自然界中具有茎和枝的植物,无论是乔木、灌木还是草本植物,都会发现其茎、枝从形态学下端向形态学上端逐渐变细,这是植物茎、枝通常具有的特点。本研究发现,未遭受水淹的喜旱莲子草的茎从形态学下端向形态学上端总体呈现出逐渐变细的趋势,符合植物茎通常具有的特点;但遭受水淹的喜旱莲子草其茎未受淹的属于形态学上端的茎段比受淹的属于形态学下端的茎段更粗,直径更大(图1)。另外,遭受水淹的喜旱莲子草位于水面上的茎段中膨大节间的茎壁厚极显著地大于未水淹喜旱莲子草植株茎的对应节间(图4)。上述研究结果说明水淹胁迫导致了喜旱莲子草的茎形态发生了改变,使得水淹的喜旱莲子草位于水面上的茎段膨大加粗。
已有很多研究表明,当木本植物茎被环割后,环割部位以上属于形态学上端的茎会膨大加粗,直径增大[8,26]。有研究表明木本植物环割后上部茎的直径增大,可能是碳水化合物在上部茎中积累而引起的[27- 28]。王文杰等研究发现茎被环割后的红松(PinuskoraiensisSieb. et Zucc.)其环割位置上部的树皮中碳水化合物含量显著升高[29]。本研究发现,水淹后喜旱莲子草未受淹的形态学上端茎段的碳水化合物含量要显著高于未水淹的喜旱莲子草形态学上端茎段的碳水化合物含量(图5),说明水淹胁迫导致了碳水化合物在喜旱莲子草位于水面上的未受淹茎段中的积累。水淹的喜旱莲子草体内碳水化合物在未受淹的形态学上端茎段中积聚,增加了形态学上端茎段中的碳水化合物含量,促进了形态学上端茎段的增粗生长,使得形态学上端的茎段膨大加粗。
碳水化合物作为植物生存与生长的物质基础,由源(光合器官)生产后输送至各需求库。碳水化合物的运输需要运输通道,运输通道受损会导致运输受阻,比如茎、枝环割使作为运输通道的韧皮部受损导致环割部位以上的形态学上端积累营养物质而发生膨大。此外,由于碳水化合物的运输是一耗能的过程,因此碳水化合物的运输不仅需要通道,同时也需要能量的提供[17]。在本研究中,喜旱莲子草茎被淹后,由于水体环境中的氧含量远低于空气中的氧含量,同时喜旱莲子草茎中致密的不具通气组织的节影响了氧气在茎中的运输,因此喜旱莲子草被淹没的茎段和地下根系难以获得和未水淹前一样充足的氧供应。尽管当氧气供应不足时缺氧的组织和器官也可以通过无氧呼吸获得一些能量,但获得能量的数量和效率是远远不能和有氧呼吸相比的,在其他条件一致的情况下同样数量的碳水化合物在有氧条件下呼吸所产生的能量比无氧条件下产生的能量要高许多倍[30](如1 mol葡萄糖在有氧呼吸的情况下最多可产生38 mol ATP,而在无氧呼吸的情况下最多只能产生8 mol ATP[31])。因此,喜旱莲子草受淹的茎段和地下根系必定会遭遇因氧气缺乏而带来的能量产生不足的问题。本研究发现,与未水淹喜旱莲子草相比,当喜旱莲子草茎遭受局部水淹时,茎未被淹的形态学上端茎段中碳水化合物含量增高(图5),同时茎直径增粗发生膨大(图1、2、3、4);尽管水淹喜旱莲子草植株的茎并没有发生类似于环割一样的物理损伤使碳水化合物运输通道受损,但水淹导致的受淹茎段缺氧而带来的被淹组织器官能量不足同样也会使碳水化合物运输受限和积累并在喜旱莲子草茎上引发与物理环割一样的组织和器官膨大的现象。本研究结果表明,在水淹的情况下,喜旱莲子草未受淹茎段会因营养物质向受淹茎段运输受限而发生与物理环割类似的茎直径增粗膨大的类环割效应。
猜你喜欢
中国果业信息(2023年12期)2024-01-09 06:59:14
汽车实用技术(2022年16期)2022-09-03 01:28:09
湖北农业科学(2021年13期)2021-08-04 03:56:54
World Journal of Clinical Cases(2020年9期)2020-05-13 07:34:08
种子(2019年4期)2019-05-28 02:04:10
小哥白尼(军事科学)(2018年3期)2018-06-15 09:56:56
现代农业科技(2017年24期)2018-01-22 21:23:15
水能经济(2017年8期)2017-10-19 18:10:32
农家顾问(2016年7期)2016-05-14 15:23:15
少儿科学周刊·儿童版(2015年7期)2015-11-24 03:37:01
