
食物丰度对宽鳍鱲的能量代谢、个性和集群行为的影响
2021-07-09 09:23付世建
生态学报 2021年11期
王 蕾, 付世建
重庆师范大学 进化生理与行为学实验室 重庆市动物生物学重点实验室, 重庆 401331
代谢是机体与环境能量和物质的交换过程,机体代谢水平是重要的生理特征,对适合度有重要影响[1- 3]。鱼类的标准代谢率(Standard metabolic rate, SMR)指其在静止且空腹条件下的最低能量消耗强度,是与生活史理论相关的重要参数[4-5]。SMR的高低直接或间接影响鱼体的生长、运动和繁殖等重要生理功能,进而对鱼类适合度产生重要影响[6]。
鱼类的“个性”指个体间行为上的稳定差异,通常用勇敢性、探索性、活跃性和社会性等来度量[7-10]。勇敢性是指机体为获取食物、配偶、空间等资源而承担风险意愿的高低[8]。探索性是指机体在陌生环境空间收集信息过程的行为特征[10]。活跃性是指动物自发行为活动特征,与觅食、寻找配偶、避敌等行为相关[7,11]。社会性则指动物对同种个体的回避和亲和反应,即加入群体的意愿[9]。研究发现,个性反映了动物的生活史策略,并与其种群动态、遗传变异以及物种形成等有关联[11-12]。
集群为动物的常见行为,自然界超过50%的鱼类种类至少在某个特定的生活史阶段表现出典型的集群行为[13]。集群行为在逃避敌害、合作捕食等诸多方面体现出生态收益,但也会加剧食物资源竞争[11]。除群体大小、游泳速度等常见物理特征,鱼群群体水平的功能特征主要体现为凝聚力和协调性两个方面。前者主要通过群体内部成员的距离来评判,而后者通常以群体成员之间游泳行为在速度和方向上一致性等来衡量[14]。
由于自然和人为活动的因素经常导致水体食物资源发生剧烈变动,包括鱼类在内的水生生物在自然选择压力下塑造出系列的行为和生理生态对策以适应食物资源波动,如下降生理功能或增加觅食活动等[15-17]。这些改变与机体SMR、个性和集群行为相关[17-18],具有重要的生态和进化意义[9]。研究发现,SMR通常随食物丰度的上升而上升[2],而个性和集群行为与食物丰度的关系的研究目前十分匮乏[19]。
本研究旨在探讨食物丰度是否影响鱼类的能量代谢、个性和集群行为及其稳定性,以小型的鲤科鱼类宽鳍鱲(Zaccoplatypus)为实验对象,测定不同食物资源条件下摄食4周前后的 SMR、个性和集群行为。选取宽鳍鱲为实验对象是因为以往的研究发现该种鱼为典型的喜好集群鱼类,且个体间存在巨大的形态、生理和行为特征差异[20-21]。
1 材料与方法
1.1 实验鱼来源及其驯化
宽鳍鱲采自重庆市武隆县白马镇乌江江段,选取雌性个体作为实验对象(宽鳍鱲雌鱼通常集体活动,而雄性成鱼往往单独活动)。实验鱼从采样点运回实验室后,在实验室自净化控温循环养殖系统中(约250L)进行2周适应。适应期水温为(27.0±0.5)℃,实验用水为除氯自来水,溶氧水平保持在90%饱和水平以上。每日以红线虫投喂至饱足两次(9:00AM和17:00PM),待食物摄食完1 h后清除粪便和食物残渣。实验期间光照周期设定为12L∶12D,日换水量约为20%。
1.2 实验设计
驯养结束后,选取鱼体健康、体重相近80尾宽鳍鱲雌鱼(大小见表1)进行体内标记(Passive integrated transponder,PIT)以便在实验过程中辨别不同个体,标记后恢复14 d,随后测定每尾实验鱼的SMR、个性指标和集群行为(4尾组成1群,N =10)(指标和具体操作见参数的测定)。测定结束后,随机分为低食物丰度和高食物丰度(N=40)两组,前者每日饱足投喂1次(9:00AM,食物为红线虫),后者每日饱足投喂2次(9:00AM和17:00PM),待食物摄食完1 h后吸取粪便和食物残渣。经过28 d喂养处理后再次测定实验宽鳍鱲的SMR、个性和集群行为。实验过程中溶氧、水温、光照周期等条件同适应期间。
1.3 实验参数的测定
1.3.1SMR的测定
采用流水式呼吸代谢仪测定实验鱼的SMR(结构见已发表论文[22])。具体操作:将实验鱼禁食1 d测定体重,随后转移至代谢仪呼吸室适应1 d。实验鱼的耗氧率采用溶氧仪(HQ30d,美国哈希公司)进行测定,每日测定8个点(08:00—22:00,间隔2h),以连续2 d测定得到的16个耗氧值的平均值作为该实验鱼的SMR。各食物丰度组均为40个重复(低、高食物丰度组分别丢失2尾和1尾实验数据)。SMR(mg O2/ h)的计算公式如下:
SMR=ΔO2×v
(1)
式中,ΔO2是与空白对照呼吸式室和实验鱼所在呼吸室出水口溶氧的差值(mg/L),v是实验鱼呼吸室的流速(L/h)。
1.3.2个性的测定
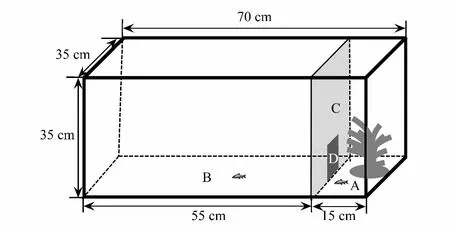
图1 鱼类勇敢性、探索性和活跃性观测装置结构示意图Fig.1 The structure of the arena for determination of boldness, exploration and activity in the present study A:隐蔽区;B:开阔区;C:隔板;D:小门
实验鱼的勇敢性、探索性和活跃性指标采用鱼类行为测定装置(图1)测定[19],该装置主体是有机玻璃制作的长方体水槽(长70 cm × 宽35 cm × 高35 cm)。以隔板将该水槽分为开阔区(长55 cm × 宽35 cm ×高35 cm)和隐蔽区(长15 cm ×宽35 cm ×高35 cm)。隔板底部正中有一边长10 cm正方形开口连通两个区域,开口处有可拆卸的小门。测定装置在测定前加入10 cm水深的曝气自来水(温度为27.0±0.5)℃。在测定装置开阔区上方放置摄像头(Logitech Webcam C930e)进行视频拍摄。
勇敢性指标以实验鱼在开阔区和隐蔽区移动情况进行分析:测定前实验鱼在隐蔽区适应0.5 h,随后移开小门,持续进行0.5 h的拍摄(15帧/s)[19]。待该指标观测结束后,通过实验鱼对陌生新奇物体的侦查行为进行探索性测定:即闭合小门,放置一个陌生物体(直径2 cm的黑色圆形或边长为2 cm蓝色正方形塑料球)于开阔区远离隐蔽区一端(距离边缘15 cm中心处),持续进行0.5 h的拍摄。探索性指标测定结束并取出陌生物体后,通过拍摄10 min的自发运动行为进行活跃性分析。
待上述指标测定结束后,实验鱼转移至社会性测定实验装置,该设备同为长方体水槽(图2,长70 cm × 宽35 cm × 高35 cm),该水槽以两块透明的隔板分成三部分,中间部分为目标实验鱼选择区域(长40 cm × 宽35 cm × 高35 cm),两边为鱼群刺激区(长15 cm × 宽35 cm × 高35 cm)。测定时先选取6尾宽鳍鱲雌鱼作为刺激鱼群移入任一一侧刺激区适应0.5 h(另一侧无刺激鱼群),随后在中间选择区放入实验鱼并进行实验视频拍摄,持续拍摄0.5 h。以目标鱼靠近刺激鱼群的程度衡量其社会性。

图2 鱼类社会性测定装置结构示意图 Fig.2 Experimental arena for observation of the sociality for fish species A、C:刺激区;B:选择区;D、E:透明隔板
实验鱼运动轨迹采用idTracker软件获取[23]。勇敢性参数采用首次进入开阔区时间(s)和开阔区停留时间比(%);探索性参数采用距离陌生物体的距离(cm)和首次靠近陌生物体时间(s);活跃性参数采用实验鱼游泳速度(cm/s)和运动的时间比(%)。社会性指标采用实验鱼与刺激鱼群的距离(cm)以及靠近刺激鱼群时间比(%)。计算方法详见已发表论文[9, 24]。实验过程均有个别视频出现技术故障无法进行视频解析(表2)。个性指标连续两日进行重复测定,以其平均值进行相关统计分析。
1.3.3集群行为的测定
实验鱼的集群行为的同样由上述长方体水槽(长70 cm × 宽35 cm × 高35 cm,无隔板)进行观测,水深10 cm。鱼群数量为4尾(鲤科鱼类在鱼群成员为4尾及以上时会出现典型的集群特征[14]),每个食物丰度处理40尾鱼随机分为10个群体。将每群实验鱼转移至观测水槽中适应5 min后开始20 min视频拍摄,视频拍摄完成后采用idTracker软件分析实验鱼在视频中的运动轨迹。根据轨迹的坐标点计算得到每尾实验鱼的自发游泳速度、实验鱼之间速度的同步性、实验鱼之间的距离(cm)和运动方向的极性等参数。实验鱼的游泳速度由实验鱼运动轨迹计算得到[14],其他参数计算如下。
速度同步性(Sv):
(2)
式中:v1、v2表示用于计算Sv任意两尾鱼的游速,4尾鱼共计算6个Sv,取其平均值为群体的Sv。
运动时极性(用于评价鱼群运动时个体的排列情况):
(3)
式中:vi(t) 为第i尾实验鱼的单位运动矢量。N是群体个体数(N=4)。极性的数值在0到1之间。当群体成员游泳方向完全一致时,极性为1;当群体成员运动方向相互抵消时,极性则为0。
个体间距离
(4)
式中:xt和yt分别代表不同实验鱼在t时刻的横、纵坐标点。
1.4 数据统计和分析
实验数据采用SPSS17.0进行统计分析。实验处理前后不同测定时间点SMR和个性指标的重复性使用可靠性分析。食物丰度处理和测定时间点对宽鳍鱲SMR和个性指标的影响采用线性混合模型(Linear Mixed Model, LMM)进行分析,其中以鱼的PIT标记编号为随机因子,实验组别(食物丰度高低)和测定时间(实验前后)为固定因子;食物丰度和测定时间对集群行为的影响采用双因素方差分析(Analysis of Variance, ANOVA)进行检验分析。若存在显著差异,则分别采用独立和配对t检验分析不同食物丰度组和测定时间的统计差异。实验数据以平均值±标准误(Mean±SE)表示,统计的显著性水平为P<0.05。
2 结果
2.1 食物丰度对实验鱼大小和SMR的影响
低食物丰度组体重和SMR在养殖过程中均没有显著变化,而高食物丰度组的体重和SMR养殖过程均显著增加(P<0.05)。养殖结束时,高食物丰度组的体重和SMR均显著高于低食物丰度组(P<0.05)(表1)。另外,可靠性分析发现高食物丰度组实验鱼的SMR不同测定时间具有重复性,即保持稳定,但低食物丰度组SMR不同测定时间不具重复性(表2)。

表2 SMR和个性参数不同测定时间的重复性
2.2 食物丰度对个性的影响
食物丰度处理对所有个性参数均无显著影响(表3)。测定时间仅对与陌生物体距离和与刺激鱼群的距离有显著影响:与实验初始值相比,与陌生物体距离在实验结束时显著增加,而与刺激鱼群的距离实验结束时显著下降(P<0.05)。可靠性分析发现,除首次进入开阔区时间、开阔区停留时间和与陌生物体距离外,其它所有指标均具有重复性(P<0.05)。

表3 食物丰度对宽鳍鱲个性参数的影响
2.3 食物丰度对集群的影响
处理前,高、低食物丰度组各个集群特征参数均没有差异,经过4周实验处理过程,低食物丰度组的游泳速度和个体间距离均上升,而高食物丰度组的两参数均没有显著变化(图3)。因此,养殖结束时,高食物丰度组的游泳速度和个体间距离均显著低于低食物丰度组(P<0.05)。测定时间对排列极性有显著影响(P<0.05),经过4周实验处理后的集群的极性显著高于实验处理前,但同一测定时间的两食物丰度组间均没有显著差异。游泳速度同步性不受测定时间和食物丰度的影响。

图3 食物丰度对宽鳍鱲集群行为特征的影响(N=10)Fig.3 Effect of food availability on schooling behavior of pale chub (N=10)*不同字母者(a,b)表示不同测定时间存在差异,星号(*)表示高低食物丰度组的数据存在差异
3 讨论
3.1 食物丰度对宽鳍鱲SMR和体重的影响
SMR是动物最重要的生理参数之一,在食物资源不足时鱼类通常会降低SMR以节约能量。研究发现大多数鱼类,如大马哈鱼(Onchorhynchuskisutch)[1]、湘云鲫 [(Carassiusauratusred var. (♀) ×CyprinuscarpioL.) (♂)][24]和南方鲇(Silurusmeridionalis)[25]等均会随着可支配食物资源的多少相应调整生理功能,维持能量消耗即SMR随之改变。本研究中,低食物丰度组宽鳍鱲的SMR显著小于高食物丰度组,说明与其他鱼类一样,宽鳍鱲也会根据食物资源的多少而调节维持能量消耗。
鱼类的SMR具有较稳定的个体差异,这种差异在一段时间可以重复[3]。然而,本研究发现SMR的稳定性差异仅在高食物丰度组存在,而低食物丰度组SMR在实验前后没有关联。这可能是因为在低食物丰度下,为了维持必要的生理功能,宽鳍鱲能量分配的比重发生了较大改变,对其SMR产生了较大的影响。以往对大西洋鲑(Salmosalar)的研究也发现,在限制摄食的条件下,SMR的稳定性下降的情况[3,26]。但对湘云鲫的研究发现,每日1次和2次饱足摄食条件下其SMR均在一定程度上保持稳定[25]。说明SMR稳定性对食物丰度的响应具有种间差异,可能与不同鱼类生境食物资源状况、觅食习性以及SMR相对大小等有关。
经过4周的不同食物丰度喂养,高食物丰度组体重显著大于低食物丰度组,这与本研究预期相符。但低食物丰度组实验过程体重没有变化,说明该食物资源仅能满足宽鳍鱲维持基本的生理功能和日常活动。以往研究发现,类似的投喂水平(即每日1次饱足摄食)下,多数鲤科鱼类表现出一定程度的生长[24]。相同投喂水平下宽鳍鱲生长较其它鲤科鱼类慢的原因可能因为宽鳍鱲喜好生活在食物资源相对分散的激流生境,维持能量需求高、日常自发活动活跃[20-21]。最近的研究比较了分布于长江流域的31种鱼类的维持能力消耗,发现宽鳍鱲仅小于同样喜好流水生境的宽口光唇鱼(Acrossocheilusmonticolus)和短须裂腹鱼(Schizothoraxwangchiachii)[27]。
3.2 食物丰度对宽鳍鱲个性和集群行为的影响
和多数鱼一样,宽鳍鱲个性存在明显的个体差异(如首次进入开阔区时间从几秒钟到超过1200 s,而停留在开阔区的时间从不到5% 到超过80%),可能有利于鱼群的集体协作以及适应多变的生活环境[28-29]。这种个体间差异的稳定性一直受到动物行为学家的关注。如个性的稳定性在食蚊鱼(Gambusiaaffinis)、孔雀鱼(Poeciliareticulata)等鱼类的研究中均有报道[30-31]。本研究通过比较实验处理前后个性指标的重复性发现宽鳍鱲的活跃性、社会性在均表现出较高的稳定性;但勇敢性和探索性指标的稳定性较低,具体原因还有待进一步研究。
相关研究发现个性受食物丰度等环境因素的影响[32-33]。然而,本研究发现,宽鳍鱲所有的个性指标均不受食物丰度的影响,类似的结果见于对湘云鲫等鱼类的研究,说明宽鳍鱲等鲤科鱼类的个性相对保守[24]。另外,本研究未发现食物丰度对个性特征产生影响也可能因为实验的食物丰度的差异不大,鱼类的个性常需在饥饿条件下才发生显著的变化[19,34-35]。
有趣的是,本研究发现,尽管食物丰度不影响所有个性参数,但会显著影响鱼类的集群行为:低食物丰度组的游泳活动增加、个体间的距离加大,即活跃性上升而凝聚力下降。这可能是低食物丰度下群体成员增加觅食活动,竞争食物资源所致。以往的研究也发现,随着食物资源的减少和个体间食物竞争加剧,鱼类往往会增加觅食活动、扩大觅食空间,进而会增加个体间距离[19,36]。研究者认为这种鱼群针对食物丰度变化的空间动态调整有利于提高其整体的生存能力[37]。值得一提的是,宽鳍鱲在集群条件下活跃性上升,而在单尾鱼的活跃性指标却不变的现象值得鱼类行为学家关注。
综上所述,宽鳍鱲个性不受食物丰度的影响。低食物资源丰度条件下,宽鳍鱲下调SMR以节约能量。集群时宽鳍鱲的活跃性上升而凝聚力下降,这可能是食物资源竞争的结果。
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
儿童时代·幸福宝宝(2020年9期)2020-09-08
军事运筹与系统工程(2019年4期)2019-09-11
电子制作(2019年23期)2019-02-23
电子制作(2018年11期)2018-08-04
中外文摘(2017年19期)2017-10-10
中国篆刻·书画教育(2017年5期)2017-06-08
中国交通信息化(2017年3期)2017-06-08
知识就是力量(2017年2期)2017-01-21
湖南大学学报·自然科学版(2014年3期)2014-12-30
