
BRD4蛋白作为恶性胶质瘤治疗靶点的研究进展*
2021-07-03 02:01:36闻乃妍刘恬恬杜艳伟黄卫东
中国病理生理杂志 2021年6期
闻乃妍,刘恬恬,付 双,杜艳伟,黄卫东
(长春中医药大学,吉林长春130117)
近年来研究表明,脑胶质瘤中广泛存在表观遗传学异常现象。与基因突变不同,表观遗传学是可逆的,它可以调控胶质瘤细胞致瘤和非致瘤状态间的转换。组蛋白乙酰化是一种重要的表观遗传学修饰方式。组蛋白乙酰化阅读器——布罗莫结构域蛋白4(bromodomain-containing protein 4,BRD4)作为胶质瘤治疗靶点,为恢复胶质瘤细胞正常的表观遗传学状态,更有效地治疗恶性胶质瘤提供了新的研究角度[1-3]。目前研究表明,下调BRD4基因或采用BRD4选择性抑制剂JQ1,能够有效地抑制胶质母细胞瘤干细胞(glioblastoma stem cells,GSCs)增殖,促进细胞凋亡,但作用效果有一定的局限性,这提示不能被JQ1抑制的胶质瘤细胞存在其它途径维持异常的自我更新,因此BRD4抑制剂与其它药物联用治疗脑胶质瘤可能具有广阔的前景。本文主要总结BRD4蛋白的生物学功能及对脑胶质瘤发生发展的影响,总结和介绍BRD4抑制剂的发展现状及发展趋势,为找寻更有效的胶质瘤治疗方法提供理论基础。
1 脑胶质瘤概述
脑胶质瘤是成人最常见的原发性脑肿瘤之一,占恶性脑肿瘤70%以上。2007年世界卫生组织分类标准中根据组织学和恶性程度将胶质瘤分为4级[4]:I级和II级为低级别胶质瘤(low-grade glioma,LGG),预后较好,主要为年轻患者,对治疗敏感性强;III级和IV级属于高级别胶质瘤(high-grade glioma,HGG),尤其是IV级多形性胶质母细胞瘤(glioblasto‐ma multiforme,GBM),恶性度最高,中位生存期仅能达到9~12个月,5年生存率不超过5%[5]。GBM可分为原发性和继发性两种,原发性占绝大多数比例,继发性较少见,临床表中男性患GBM的发病率明显高于女性。GBM具有浸润性、侵袭性、迁移性等多种生物学特征,局部侵袭性能够侵袭到脑实质、蛛网膜下腔、血管周围腔和白质束;浸润性可以浸润到距离原发肿瘤1~2 cm处[6];GSCs可沿脑白质进行迁移,即便进行手术切除,仍然会伴有很高的复发率[7]。目前针对于恶性胶质瘤的标准治疗方法是最大安全范围手术切除肿瘤组织配合放化疗,虽然有一定疗效但是患者中位生存期小于2年,因此亟需找寻更有效地治疗脑胶质瘤的方法[1,8]。
2 BRD4蛋白的生物学功能
BET(bromodomain and extra-terminal domain)家族是一类包含布罗莫结构域的蛋白,包括BRD2、BRD3、BRD4和BRDT四种亚型,BRD4是其中被研究得最多最深入的一个亚型[9]。BRD4蛋白具有组蛋白乙酰化阅读器功能,能够选择性地与组蛋白尾部的乙酰化赖氨酸残基相结合,通过布罗莫结构域募集转录调节因子与乙酰化组蛋白相结合,从而调节一系列重要的生物过程,例如基因转录调节、染色质重塑、DNA损伤和修复等[10]。
在结构上,BRD4蛋白有两个保守的串联布罗莫结构域BD1和BD2,以及一个ET结构域。BD1和BD2可与组蛋白和(或)转录因子上的乙酰化赖氨酸残基结合。BRD4的ET结构域主要通过与染色质调控因子(精氨酸脱甲基酶JMJD6和赖氨酸甲基转移酶NSD3)相互作用而发挥转录调节作用。此外,ET结构域还与ATP依赖性染色质重构复合物SWI/SNF和克罗莫结构域解旋酶DNA结合蛋白4(chromodo‐main helicase DNA binding protein 4,CHD4)相互作用,表明BRD4具有改变染色质结构的作用[9]。
最初,发现BRD4具有基因转录调节作用是由于BRD4可以与正性转录延伸因子b(positive transcrip‐tion elongation factor b,P-TEFb)复合体和调节因子构成的蛋白复合物相结合[11]。P-TEFb复合体是一个由细胞周期蛋白依赖性激酶9(cyclin-dependent ki‐nase 9,CDK9)及其调节亚基cyclin T1、T2或K构成的异二聚体,通过磷酸化RNA聚合酶II(RNA poly‐merase II,Pol II)延长复合物来激活Pol II[12]。BRD4有2个结构域可以与P-TEFb相互作用,分别是C末端结构域(C-terminal domain,CTD)和BD2结构域。CTD与cyclin T1和CDK9相互作用;BD2结构域能够识别cyclin T1的乙酰化区域。7SK核内小RNA与HEXIM1蛋白形成核糖核蛋白复合物,该复合物可以隔离P-TEFb并使其功能失活,BRD4阻止P-TEFb与7SK/HEXIM复合体结合,从而保持P-TEFb的活性。因此,BRD4是乙酰化组蛋白和基因转录之间相互作用的重要条件[13]。此外,BRD4能够募集JMJD6到增强子使H4R3me1和H4R3me2去甲基化。增强子与BRD4/JMJD6/P-TEFb复合物相结合促进启动子区域Pol II的释放,促进转录延长[9]。
BRD4是一种在哺乳动物组织中广泛表达的核蛋白,最初被认为是一种控制细胞周期的蛋白[14]。BRD4通过与乙酰化组蛋白(H3Lys14和H4Lys5/8/12)及非组蛋白结合,调节DNA复制和基因转录,从而影响细胞周期[15]。在G0/G1期转化过程中,细胞通过有丝分裂刺激诱导BRD4表达。当细胞进入M期阶段,BRD4与染色体相互作用,标记那些在G1期转录的基因,以确保细胞周期正常进展。微量注射BRD4抗体到HeLa细胞核中可完全抑制细胞的有丝分裂,表明BRD4在细胞周期中也发挥重要作用。在胶质瘤细胞及GSCs中沉默BRD4表达或利用BRD4抑制剂能够有效抑制细胞活力和增殖能力,通过抑制p21、cyclin D1等细胞周期相关蛋白的表达诱导细胞大量停滞在G1期,S期比例减小。长链非编码RNA(long noncoding RNA,lncRNA)HOTAIR对胶质瘤细胞增殖具有重要作用,BRD4能够与HOTAIR启动子相结合,调节lncRNA的表达,表明BRD4可以通过调控lncRNA的表达促进GSCs增殖[16-18]。
除了调控基因转录和细胞周期,BRD4在DNA损伤检查点的激活调控方面也发挥着重要作用。BRD4通过募集凝集素II染色质重构复合物到乙酰化组蛋白上,从而发挥内源性DNA损伤应答信号抑制剂的作用。BRD4缺失导致染色质结构松弛,γH2AX噪点增多,而当BRD4表达增强会降低γH2AX表达水平,减弱DNA损伤信号对电离辐射的应答[19]。Wen等[18]发现,siBRD4或JQ1能引起GSCs中γH2AX表达水平升高,DNA损伤加重。
GSCs被认为是导致脑胶质瘤术后高复发率、对放疗抵抗和化疗耐药的主要原因[20-21]。通过转录组测序发现,对JQ1敏感的GBM细胞中OLIG2的表达水平上调明显;OLIG2是proneural亚型GBM的标志物,异常表达的OLIG2与其他神经发育转录因子相互作用可诱导GBM细胞多能性分化,提示JQ1对GSCs表型稳定产生重要影响[22-23]。其它研究表明,敲除BRD4能够减弱GSCs的干性,使乙醛脱氢酶1活性下降,干性调节因子表达下调,神经球成球能力减弱。此外,抑制miR-142-5-p或重新激活Wnt/βcatenin信号通路可逆转沉默BRD4对GSCs干性的抑制作用,表明BRD4/miR-142-5-p/Wnt/β-catenin可能是胶质瘤细胞中负责维持干细胞特性的信号通路,这可能成为一个具有前途的表观遗传学治疗脑胶质瘤的方法[15]。Wen等[18]的研究表明,siBRD4或JQ1可降低GSCs神经球的成球能力,抑制GSCs自我更新;JQ1还可有效抑制GSCs标志物nestin的表达水平,提示BRD4对于GSCs维持干性具有重要作用。
3 BRD4抑制剂进入前期临床试验的进展
2010年Filippakopoulos等[24]发现了第一个具有甲基三唑并二氮杂䓬类骨架结构的选择性BRD4抑制剂——JQ1。之后的几年里越来越多的BET抑制剂被发现。目前虽然还没有BET抑制剂获得FDA的批准,但是有几种药物已处于临床试验阶段(表1)。

表1 BET抑制剂的临床试验Table 1.Clinical trials with BETinhibitors(https://www.clinicaltrials.gov/)
目前,针对BET抑制剂耐受性和毒副作用的研究还不够全面。I期药物存在一定的剂量限制性毒性(dose-limiting toxicity,DLT),表现为头痛、呕吐、腹泻、疲劳、厌食、味觉障碍、血小板减少等。DLT较小的药物有MK-8628/OTX-015、CPI-0610、RO6870810/TEN-010和GSK525762。文献表明,在OTX015的Ib期试验中,47例晚期实体肿瘤患者中仅有7例出现部分不良反应,相较于其它BET抑制剂,OTX015的毒副作用反应较少[25]。在给药剂量方面,最初许多关于用药剂量的研究表明药物连续剂量应为每天1次。然而,由于药物毒性等多种原因,对于一些药物根据肿瘤类型制定了替代方案,推荐进行间歇性给药。以OTX015为例,实体瘤给药剂量为每日80 mg,造血系统恶性肿瘤推荐间歇性给药,每日80 mg,给药2周,停药1周。绝大多数的BET抑制剂是口服的,只有RO6870810是一种用于皮下注射的化合物[13]。目前,进入临床前期试验用于治疗脑胶质瘤的BET抑制剂只有MK-8628(NCT02296476),BET抑制剂对脑胶质瘤的治疗作用有待进一步的研究。
4 BRD4抑制剂在脑胶质瘤治疗中的作用
4.1 JQ1和I-BET Fiskus等[26]发现,JQ1和I-BET具有相似的化学结构和靶向抑制模式。这2种抑制剂都可以与BET蛋白的布罗莫结构域竞争性地结合乙酰化肽,从而导致细胞暴露于这些化合物的时候,BET蛋白从染色质上被置换下来,影响BET蛋白发挥正常功能[24,27]。JQ1是一种基于噻吩二氮平的小分子,在低纳摩尔范围内对BET亚家族表现出良好的抑制作用。JQ1的晶体结构尤其与BRD4的BD1结构域具有良好的结构互补性,因此JQ1对BRD4蛋白的效果特别明显。此外,JQ1具有良好的血脑屏障通透性(AUCbrain/AUCplasma=98%),这为应用于治疗脑胶质瘤提供了可能[28]。
JQ1在不同遗传背景的胶质瘤细胞中具有广泛的活性,对不同遗传背景的胶质瘤细胞均可有效地抑制细胞增殖,促进细胞凋亡,诱导细胞产生细胞周期阻滞。胶质瘤的一些关键信号通路如c-Myc、Akt、p53或Rb通路等对JQ1都表现出很高的敏感性[29-30]。除了传统的药物治疗以外,近年来还出现了非电离光动力疗法(photodynamic therapy,PDT)治疗脑胶质瘤的案例,PDT与传统的胶质瘤细胞治疗相比具有更明显抗肿瘤优势。JQ1可以抑制iNOS表达,提高PDT细胞毒性,抑制iNOS/NO对PDT的抵抗作用,从而有效提高抗肿瘤效果;经PDT后,JQ1可以抑制U87细胞中BRD4与NF-κB p65之间的相互作用,这表明JQ1能够极大地提高PDT治疗脑胶质瘤或其他恶性肿瘤的临床效果;这是在体外实验中展现出非常好的疗效,但还需进一步开展体内实验来深入验证[31]。JQ1和I-BET除了抑制胶质瘤细胞增殖外,在促进细胞分化方面也具有一定的潜能。恶性胶质瘤细胞能够依赖特有的表观遗传途径抑制自身的分化,从而维持异常的自我更新[32]。在弥漫性内生性脑桥胶质瘤(diffuse intrinsic pontine glioma,DIPG)中,JQ1在有效抑制DIPG细胞增殖的同时能够有效促进肿瘤细胞终末神经元分化[33]。I-BET151有效抑制胶质瘤细胞增殖的同时可以有效促进鼠源胶质祖细胞分化为少突胶质细胞[34]。关于BRD4抑制剂在胶质瘤分化方面的作用还很少,有待进一步研究。
4.2 OTX015(MK-8628) OTX015(MK-8628)是一种新型的BRD2/3/4抑制剂,具有良好的通过血脑屏障的能力(肿瘤∶血浆=1∶10),其在肿瘤中的水平是正常组织的7至15倍,并与肿瘤组织优先结合。由于OTX015的亲脂性,瘤周皮肤和肌肉组织OTX015蓄积较多[35]。OTX015对胶质瘤细胞的增殖和细胞周期进展起到了明显的抑制作用,并且比JQ1效力更强[半数细胞生长抑制浓度(the concentration causing 50%cell growth inhibition,GI50)=183.0~223.1 nmol/Lvs618.1 nmol/L]。针对OTX015的临床前药理学研究为OTX015进入临床治疗胶质母细胞瘤提供了理论基础。目前,对OTX015治疗成年胶质母细胞瘤患者的剂量递增研究(NCT02296476)正在进行[36]。
4.3 BRD4抑制剂与其他药物联合用药的发展前景 胶质瘤具有很强的异质性,单一小分子药物治疗胶质瘤的效果有限,因此BRD4抑制剂与其他药物联合用药为脑胶质瘤的治疗开辟一个新途径[34,37]。目前,已有几种BET抑制剂与其它抑制剂联合用药处于临床试验阶段(表2)。
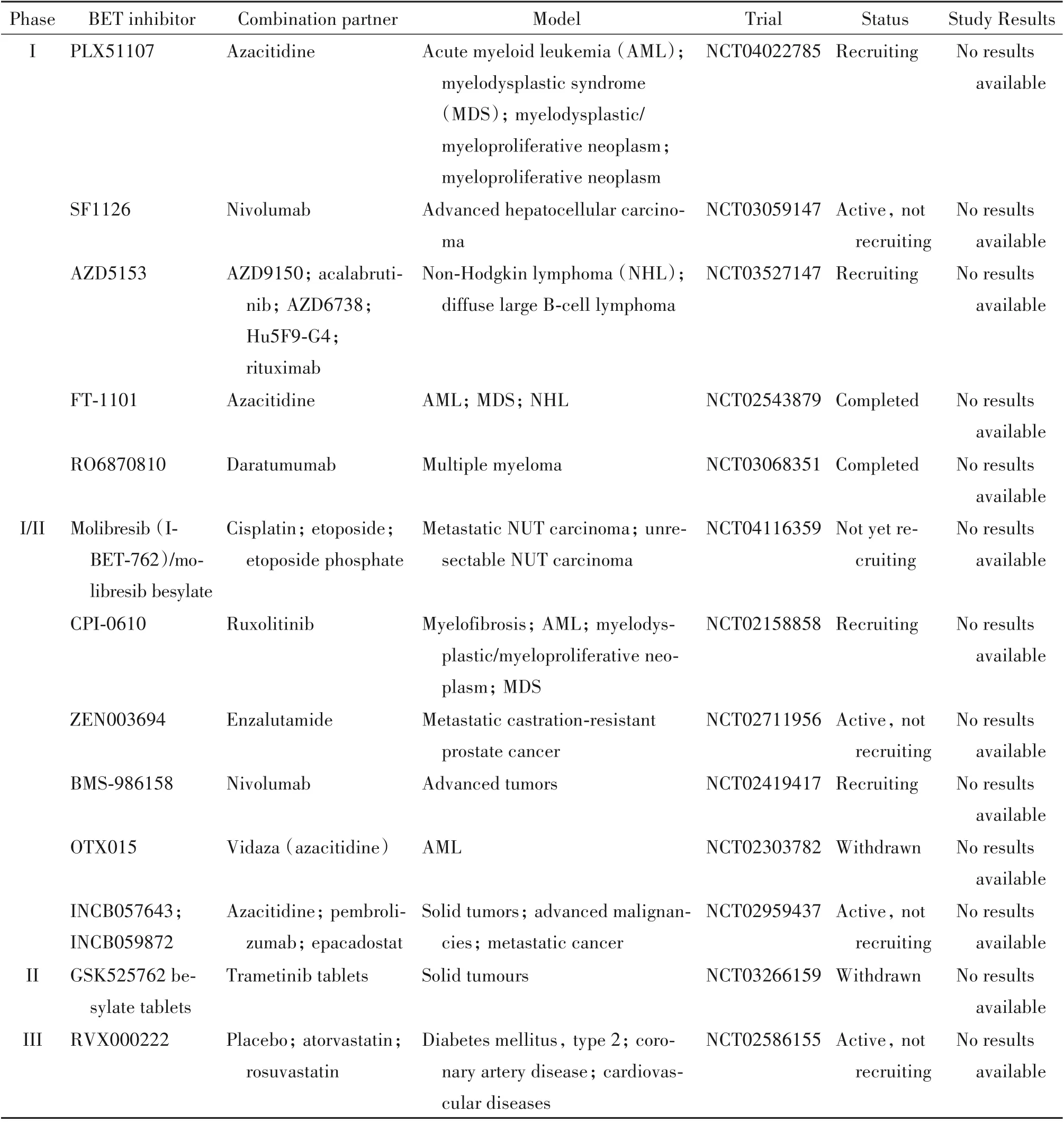
表2 BET抑制剂在联合治疗中的临床前研究Table 2.Preclinical studies of BETinhibitor in combination therapy(https://www.clinicaltrials.gov/)
4.3.1 BRD4抑制剂与组蛋白脱乙酰酶(histone deacetylase,HDAC)抑制剂联合用药 Fiskus等[26]的研究表明,在急性髓系白血病(acute myeloid leuke‐mia,AML)细胞中采用JQ1与HDAC广谱抑制剂panobinostat联合用药具有协同作用。JQ1和panobi‐nostat协同作用显著降低了神经母细胞瘤细胞中NMyc和Bcl-2的表达,有效抑制肿瘤发展[38]。同样,在胶质瘤细胞中panobinostat和JQ1或OTX015联合用药对U87或U251细胞具有良好的协同作用,能够有效抑制抑制细胞增殖,提高了caspase活性,有效诱导细胞凋亡,提高对细胞的毒性。此外,panobinostat与JQ1或OTX015联合用药对GBM相关致癌基因或通路的抑制作用增强,并且高度诱导GBM相关抑癌基因的表达增强[30]。抑制BRD和HDAC蛋白会抑制胶质母细胞瘤细胞糖酵解和氧化能量代谢,导致细胞内ATP产生和ATP总量的减少[39]。JQ1/OTX015和panobinostat都有良好的血脑屏障通透性,这为2种表观遗传学药物的联合应用提供了可能[40-41]。
近年来,有文献提出了多元联合治疗的概念:HDAC抑制剂(如panobinostat和vorinostat)、BRD抑制剂(OTX015)和多激酶抑制剂(sorafenib)联合用药治疗脑胶质瘤。HDAC抑制剂与BRD抑制剂联合的基础上加入sorafenib,有效抑制了U87-EGFRvIII模型和患者来源的异种移植小鼠模型肿瘤生长,延长了小鼠生存期[39]。三联疗法治疗脑胶质瘤疗效显著,在一定程度上可以考虑作为恶性肿瘤的有效治疗方法。
4.3.2 BRD4抑制剂与替莫唑胺(temozolomide,TMZ)联合用药 TMZ是一种口服的二代烷化剂咪唑四嗪类衍生物,易通过血脑屏障,是临床治疗恶性胶质瘤的标准一线化疗药物。TMZ主要通过攻击肿瘤细胞DNA,使之产生烷基化损伤,进而形成DNA交联,最终导致肿瘤细胞死亡。TMZ同步配合放化疗虽然具有一定疗效,但有效率不足50%,造成这一结果的主要原因与胶质瘤细胞对TMZ的耐药密切相关[42-43]。基因富集分析发现,对JQ1敏感的GBM细胞主要富集在DNA修复机制相关通路上,因此对JQ1敏感的胶质瘤细胞对烷化剂有更好的反应。将TMZ与JQ1相结合可以通过调节DNA损伤应答来克服细胞对TMZ耐药产生的DNA修复机制的激活。此外,JQ1能够上调促凋亡和抑制细胞增殖相关基因的表达,使GBM细胞更容易受到TMZ细胞毒性的作用,JQ1与TMZ联合用药显示出非常好的协同作用,为JQ1进一步的临床应用和解决TMZ耐药提供了具有可行性的方法[15,23]。
OTX015与TMZ联合用药能够有效提高异种移植脑胶质瘤小鼠的生存率,并且OTX015剂量组均未发现任何毒性,而TMZ组5 d后小鼠体重明显下降。在不存在药物毒性的情况下,OTX015和TMZ联合用药能够有效提高GBM模型小鼠生存期。OTX015与TMZ、依维莫司或SN38联合用药时对胶质瘤细胞也表现出协同作用。虽然单独使用OTX015发挥了良好的抗恶性胶质瘤作用,但随着对表观遗传药物领域研究的不断深入,OTX015联合常规化疗或靶向药物对恶性胶质瘤的治疗更能发挥良好的潜力[36]。此外,I-BET151可通过PUMA诱导GBM细胞增强对TMZ的敏感性[44]。综上所述,BRD4抑制剂与TMZ联合用药,为解决GBM对TMZ耐药提供了新思路。
4.3.3 鸡尾酒治疗法 如何在抑制GBM细胞增殖的同时促进细胞分化是提高GBM治疗有效性的关键问题。有研究提出小分子介导的重编程可以将GBM细胞转化为神经元。研究表明,forskolin、ISX9、CHIR99021、I-BET151和DAPT混合形成的小分子鸡尾酒可以高效地将GBM细胞转化为神经元。遗传分析显示这种小分子鸡尾酒能够上调Ngn2、Ascl1、Brn2和MAP2基因表达,导致神经元重编程。这些重编程细胞高表达神经元标志物和前神经转录因子,形态学和免疫荧光检测证明重组细胞具有与神经元表型相似的特征。此外,鸡尾酒法可以抑制GSCs神经球的形成,证实该疗法能够抑制GSCs的自我更新能力[45]。这项较新颖的研究证明了将GBM细胞重编程为神经元是一种有潜力的治疗恶性胶质瘤的方法。
5 结语与展望
近年来,BRD4在脑胶质瘤中发挥的作用逐步受到关注,但是目前针对于BRD4抑制剂在脑胶质瘤中的研究和应用仍然具有局限性。治疗GBM的有效性受到血脑屏障和对单一药物易产生耐药性的限制,因此剂型优化和联合用药有望成为提高BRD4抑制对脑胶质瘤治疗效果的有效手段。有文献报道,与无载体组相比,利用转铁蛋白纳米粒子(transfer‐rin-coated nanoparticle,Tf-NP)为载体携带TMZ和JQ1可有效促进胶质瘤细胞DNA损伤和凋亡。对GBM原位移植瘤小鼠模型进行小鼠活体成像检测表明,携带TMZ和JQ1的Tf-NP具有良好的血脑屏障渗透性,能够直接携带药物作用于肿瘤组织。此外,与无载体组相比,Tf-NP联合用药载体组能够更好地减轻小鼠肿瘤负担,延长小鼠生存期[46]。药物载体的创新性为BRD4抑制剂更好应用于治疗脑胶质瘤提供了新的研究方向。目前的研究在一定程度上证明了BRD4是极具潜力的脑胶质瘤治疗靶点,未来进一步深入研究BRD4将为探寻胶质瘤表观遗传学新靶点、新机制提供理论依据。
猜你喜欢
中国生物化学与分子生物学报(2022年8期)2022-09-08 00:39:50
国际呼吸杂志(2019年4期)2019-03-12 01:07:30
中华老年多器官疾病杂志(2016年7期)2016-04-28 08:43:05
癌症进展(2016年10期)2016-03-20 13:15:43
中国组织化学与细胞化学杂志(2016年4期)2016-02-27 11:15:53
磁共振成像(2015年8期)2015-12-23 08:53:14
吉林大学学报(医学版)(2015年5期)2015-12-16 15:43:57
医学研究杂志(2015年5期)2015-06-10 06:43:26
中国当代医药(2015年16期)2015-03-01 02:03:13
中国当代医药(2015年9期)2015-03-01 02:02:00
