
生物炭稻田施用下的土壤固碳减排效应及其微生物群落结构分析
2021-07-02 08:59孙再庆符菁徐晓云赵远
农业与技术 2021年12期
孙再庆符菁徐晓云赵远
(1.常州市生态环境监控中心武进分中心,江苏 常州 213000;2.常州大学环境与安全工程学院,江苏 常州 213000)
我国是农业耕作大国,每年秸秆产量相当可观,有大量的水稻秸秆直接被就地焚烧。秸秆焚烧生成大量温室气体,排放大量粉尘颗粒物,流失大量土壤有机质。将水稻秸秆制备成生物炭还田后不仅可以避免上述危害的发生,而且还可以改变土壤中细菌总量和微生物种群结构特征。
生物炭是秸秆原料在低氧高温条件下裂解炭化制备的新型环境类功能性材料。其具有孔隙结构发达、官能团丰富、阳离子交换量和含C量较高、生物炭芳香化程度高且具有生物化学和热稳定性等性质。不但具有改良土壤肥沃度、降低温室气体(CH4、CO2和N2O)排放的能力,而且具有极其优良的持续性碳封存能力,从而改变土壤生物组成。
生物炭作为掺合物混入稻田土壤后,通过对微生物的直接或间接作用,影响其生长代谢。因此,研究生物炭作为土壤基肥混入,对土壤微生物群落结构的影响意义重大。研究证明,在土壤中加入生物炭后,土壤中的细菌、真菌群落结构都发生复杂化。韩光明等发现,在菠菜土壤中添加适量生物炭后,土壤根系功能性微生物数量(根际细菌及氨化细菌、放线菌、真菌、固氮菌和反硝化细菌等)发生明显的变化。Graber等在辣椒种植过程中掺入一定比例生物炭,对土壤中芽孢杆菌、丝状真菌、假单胞菌等微生物的群落丰度产生影响,研究发现,生物菌群数量随着生物炭比例增加而提升。但是,因为生物炭的添加对土壤中的生物影响非常复杂且不同的微生物需要不同特定的生长环境,所以生物炭的添加可能对原有土壤中的微生物生长没有影响。宋延静等通过研究在滨海盐碱土施加不同比例生物炭后,发现生物炭添加后Ideonella和Skermanella等属的丰度相对会增加,而对固氮螺菌属生长没有产生影响。
结合作物产量同温室效应的气体排放强度研究成为当下综合评估农田管理措施的导向和热点,施用不同种类的生物炭和不同量的生物炭对CH4、N2O、CO2等温室气体排放的影响差异性较大。刘翔等以伊犁河谷农田为研究对象,探究土壤冻融过程中N2O、CO2和CH4的排放量对生物炭的响应程度,发现生物炭的添加虽然会促进冻融土壤过程的CO2排放,但是对CH4、N2O排放的抑制作用更为明显。Zhang等研究发现,生物炭能够抑制N2O的排放,但是会较小程度地提高CH4的排放。
本试验针对丹阳大田中不同施肥基质的土壤微生物群落结构和CH4、N2O、CO2气体排放,通过高通量测序法和静态箱-气相色谱法,探究不同温度条件和不同区域水稻秸秆制备的生物炭对土壤微生物群落结构和CH4、N2O、CO2气体排放的影响,对于固体废弃物再利用以及固碳减排的研究有重大意义。
1 材料与方法
1.1 试验地与试验材料
本试验水稻田位于丹阳市珥陵镇,具体位置为E119.598452°,N31.86952°。珥陵镇具有得天独厚的地理位置,属于亚热带季风气候,春季和秋季为冬、夏季风的转换季节,冷暖气团相互交织,导致寒暑干湿差异性显著。
本试验中设计的收割后秸秆生物炭制备方式:将收割的水稻秸秆放置在干燥的环境中自然风干;将风干后的水稻秸秆切割成小段,放入多功能粉碎机(转速为3万r·min-1),分成4步粉碎(时间为30s、30s、90s、30s);用100目筛子将粉碎后的秸秆过筛,并装入30mL的坩埚中压实(将颗粒之间的空气挤出);坩埚放入马弗炉中,在500℃条件下炭化120min。炭化后得到2种秸秆生物炭,将300℃、700℃条件下制备的生物炭分别标注为DY-300、DY-700,生物炭的理化性质见表1。本试验使用的秸秆生物炭均未经过活化,直接施入土壤中。
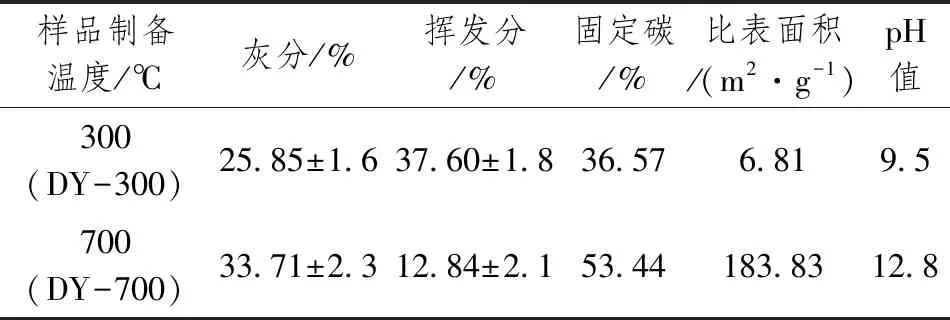
表1 不同温度下制备的秸秆生物炭的理化性质
1.2 试验设计
本课题针对丹阳大田,将生物炭作为外源性基质添加进土壤中,研究水稻秸秆经过不同温度炭化处理后,土壤微生物群落结构和物种丰度的变化,同时针对水稻不同生长时期CH4、N2O、CO2排放通量的变化规律进行分析。具体分组如表2所示。
试验为田间小区试验,前茬作物为小麦。采用随机区组设计,设置施用生物炭DY-0、DY-300和DY-700共3个处理。6月上旬播种,7月上旬施用生物炭,7月中旬移栽,分蘖期7月28日采集土壤样品和气体样品,8月13日采集气体样品,抽穗期8月28日采集土壤样品和气体样品,9月19日采集气体样品,成熟期10月12日采集土壤样品和气体样品。土壤样品可短期保存于-20℃冰箱,或者长期储存于-80℃超低温冰箱;气体样品可常温常压储存在铝箔采样袋,需尽快送样检测。
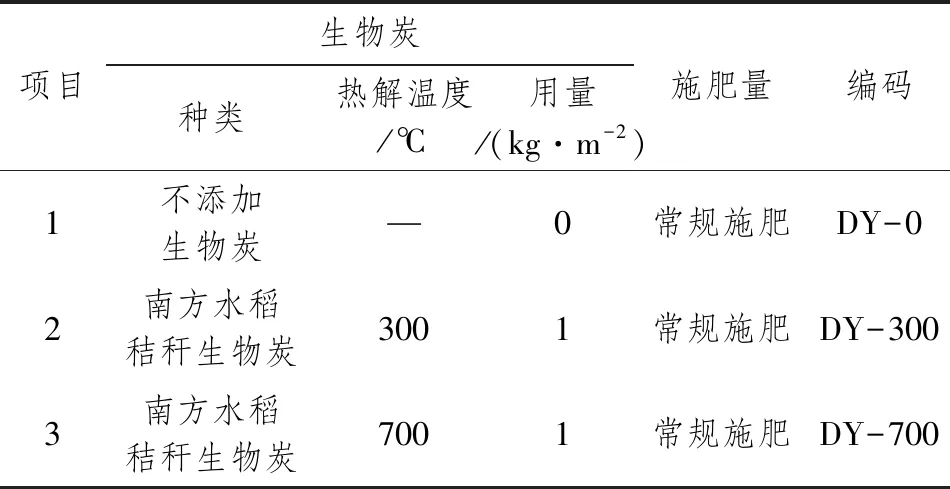
表2 丹阳实验大田分组表
1.3 土壤CO2、CH4和N2O排放通量测定
本试验中土壤温室气体的采集方式是静态箱,测定方式是气相色谱法。静态箱-气相色谱法联用是田间温室气体测定的常用方法。
依据箱内气体浓度变化率计算土壤温室气体排放通量,计算公式:
(1)
式中,F为温室气体通量,mg·(m-2·h-1)或μg·(m-2·h-1);ρ为标准状态下温室气体质量浓度,kg·m-3;V为静态箱体积,m3;A为静态箱水平面面积,m2;dc、dt为温室气体排放速率,mL·(m-3·h-1);T为采样箱内气体温度,℃;T0为0℃对应的热力学温度值。
依据排放通量计算土壤温室气体累积排放通量[14],计算公式:
(2)
式中,CE为气体累积排放量,mg·m-2;F为气体排放通量,mg·(m-2·h-1);i为第i次气体采样;ti+1-ti为2个相邻测定日期的间隔,d;n为累积排放量观测时间内总测定次数。
1.4 土壤微生物高通量测序
1.4.1 土壤样品采集
在DY试验大田中,考虑到每组试验田的面积较小且形状规整,因此本次采样选用对角线法,单组试验田共设置5个点,每个点采集地下0~10cm表层土壤200g左右,将混匀后的水稻田土样装入无菌塑封袋中,挤出袋内多余的空气,贴上标签。-20℃冰箱保存待用(短期保存)。
1.4.2 微生物DNA抽提、扩增及测序
通过强力土壤DNA提取试剂盒抽提水稻田环境中高品质的总DNA(条带长度:15000bp),再按照PCR扩增仪(2720PCR+)提供的试验流程进行,引物由宏众基因公司提供,其中PCR正向引物序列为6’—AYTGGGYDTAAAGNG—3;PCR反向引物序列为6’—TACNVGGGTATCTAATCC—3’。经过16S rRNA扩增(条带长度:400bp),最后进行建库测序,测定土壤中细菌界微生物。采用琼脂糖凝胶电泳试验对提取的长链DNA和PCR反应的短链DNA进行定性分析,作为测序工作继续进行的依据。
1.5 秸秆生物炭理化性质的测定
1.5.1 秸秆生物炭制备
水稻秸秆收集后,风干5d,洗净,经多功能高速粉碎机切割后,过100目筛,置于30mL陶瓷坩埚中,将秸秆压实后盖上盖,再将其置于马弗炉内,以15℃·min-1的升温速度分别升温至300℃、700℃,保温3h,自然无氧冷却至室温。取出后研磨,过100目筛,密封干燥保存。
1.5.2 秸秆生物炭表征
生物炭的产率采用重量法计算;pH值利用pH计(FE28,中国,梅特勒-托利多)测定,方法参照GB/T 12496.7—1999标准;污泥生物炭比表面积(BET)采用物理吸附仪(Autosorb-iQ2-MP)进行测试,方法参考Joyner L G等研究方法;采用扫描电镜(SEM)仪(JEOIJSM-6360LV)进行表面形貌分,方法参照苗壮等研究方法。
2 结果与分析
通过上述步骤得出的结果进行分析,生物炭基肥理化性质分析、温室气体排放通量分析、琼脂糖凝胶电泳图分析、微生物群落多样性分析、土壤微生物群落结构与温室气体排放的相关性分析,具体分析如下。
2.1 生物炭基肥理化性质分析
生物炭作为一种外源性基肥,其多孔性和颗粒结构对土壤物理结构和化学性质有直接和间接影响,本试验通过马弗炉炭化水稻秸秆制得生物炭基肥,并对其理化性质进行分析。具体分析结果如表3、图1所示。
如表3所示,700℃条件下生物炭中的含C量高于300℃条件下生物炭,高出31.6%。2种条件下制备的生物炭的比表面积具有显著差异,两者的比表面积差异高达27倍,700℃高温条件下使得羟基、烃基、芳香烃环先后发生裂解,产生大量的挥发性气体,气体的释放冲击生物炭表面使得孔隙的张开,形成丰富的孔径结构,从而致使比表面积的增加。通过分析图1中DY-300和DY-700结果表明,相比300℃条件下制备的生物炭,700℃的微孔数量密集、孔隙率高、表面形状规整、分层现象明显、出现少量晶面,说明在700℃下生物炭基肥的理化性质优于300℃下。
表3 生物炭基肥理化性质

图1 生物炭的电镜扫描
2.2 温室气体排放通量分析
在DY实验大田中,用静态箱-气相色谱法测定水稻生长过程中土壤CH4、N2O、CO2浓度,分析水稻秸秆在不同温度条件下制备的生物炭对水稻生长过程中土壤CH4、N2O、CO2气体排放通量和累计排放量的影响,并比较水稻不同生长时期CH4、N2O、CO2气体排放的差异性。
将采样气体浓度同标准样品校核后,计算出一定间隔情况下气体浓度的差值,并带入公式(1)和公式(2)求出不同气体在不同时期的排放通量和累积排放量,具体排放通量和累积排放量分析如图2至图5所示。
观察CH4、N2O、CO23种气体在不同时期的排放通量以及不同处理后气体累积排放量的差异。
由图2可知,在生物炭添加进土壤的初期,由于生物炭多孔和富含有机质的特性,使土壤的孔隙度和有机质增加,致使微生物的有氧呼吸作用加强,在DY-300和DY-700的处理中明显CO2的排放通量显著高于DY-0的处理。随着微生物反应和生物炭矿化作用,经过生物炭处理的样品CO2排放的优势趋于显著。处理中期,由于生物炭的优良性状,促使DY-300和DY-700样品的CO2排放量明显低于DY-0的样品。水稻生长的后期,由于生物炭的综合性作用逐渐减弱,对于稻田微生物的优势性作用缓慢减弱,土壤中CO2的排放在DY-0、DY-300和DY-700处理之间趋于稳定。
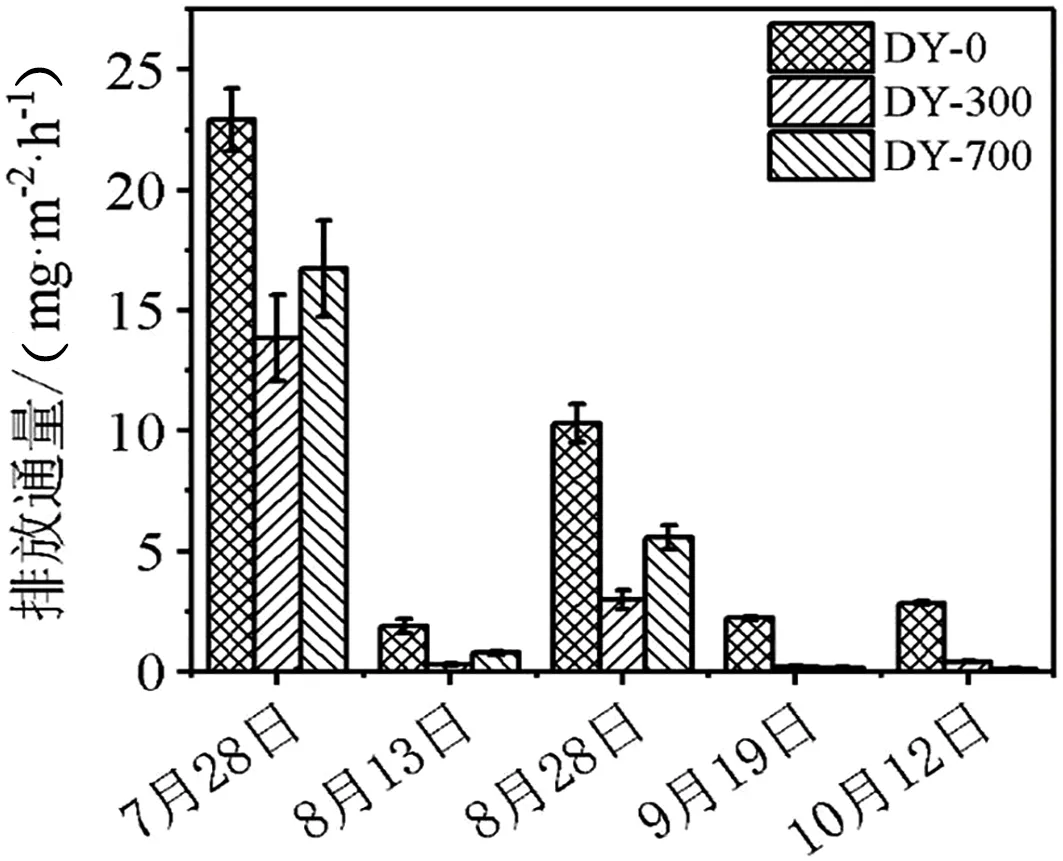
图2 不同时期CO2气体排放通量
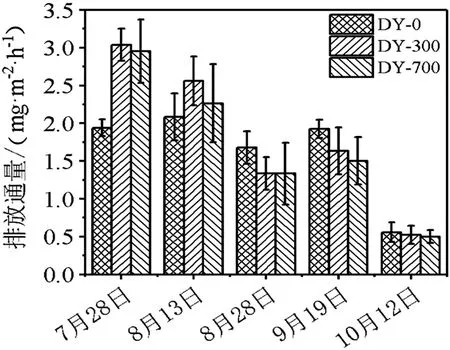
图3 不同时期N2O气体排放通量
由图3可知,在水稻生长的前期,DY-0处理中土壤含氧量低,微生物进行反硝化过程,将硝酸盐或者硝态氮还原成氮氧化物(N2O、NO等)和氮气。因此,水稻生长前期N2O排放通量>水稻生长中后期N2O排放通量;未经生物炭处理的水稻田土壤N2O排放通量>经过生物炭处理后的N2O排放通量。基于生物炭基肥水稻田土壤的N2O排放抑制性能为DY-300>DY-700>DY-0。生物炭对N2O排放的影响既是直接作用又是间接性作用。生物炭通过改变土壤的pH值、土壤的有机质含量、土壤的孔隙度、土壤的含水率等性质间接对N2O排放通量产生影响。
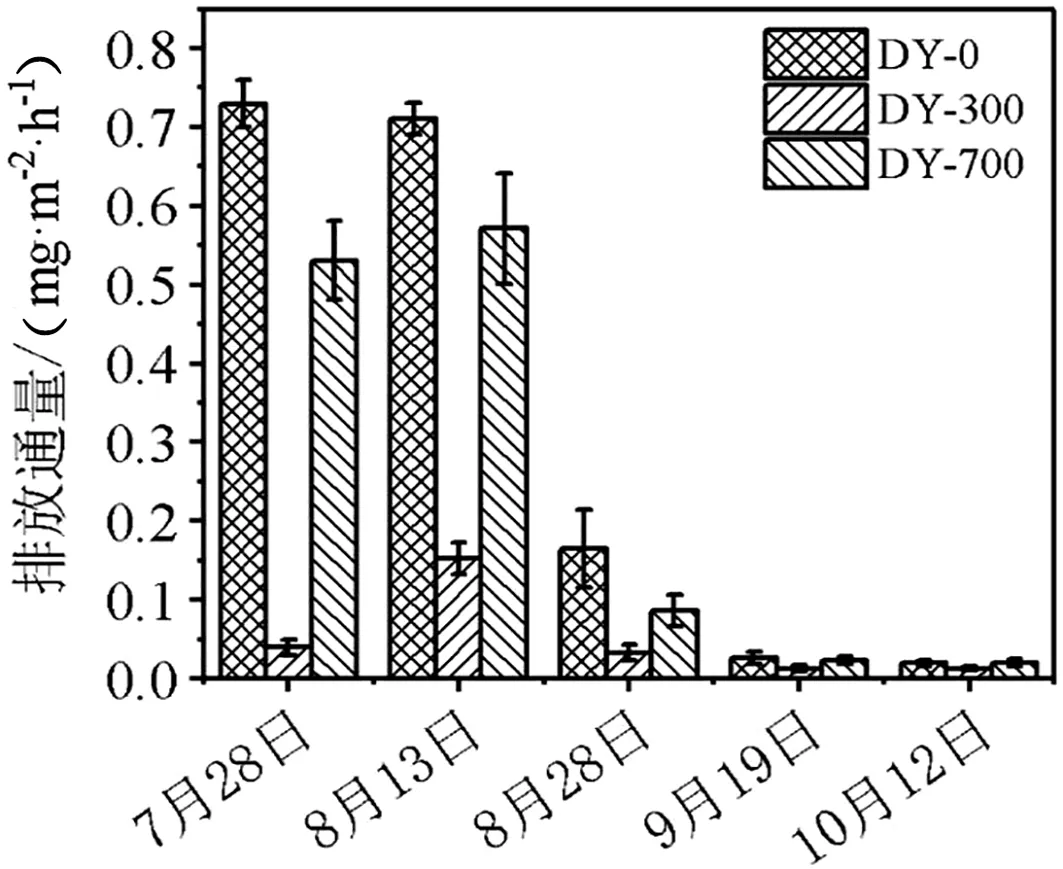
图4 不同时期CH4气体排放通量
由图4可知,在水稻生长前期,CH4排放通量为DY-0>DY-700>DY-300,土壤淹水其还原电位降低,甲烷促生菌大量生成,甲烷的排放量显著提升。水稻田晒田期间,土壤中含水量降低,与空气的气体交换加强,甲烷促生菌产甲烷受到抑制,也可能是由于季节性变化导致CH4排放通量整体下降。由图4得知,水稻生长前期土壤中CH4气体排放量显著高于水稻生长后期。
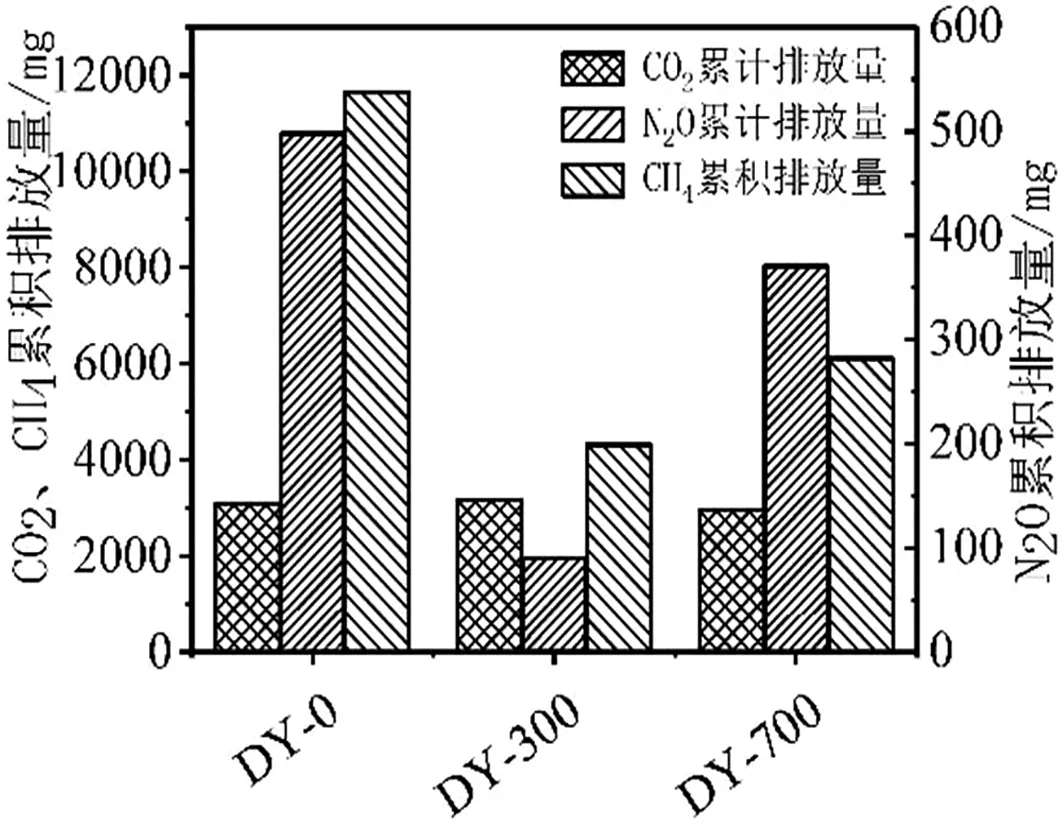
图5 不同生物炭处理气体累积排放量
由图5可知,水稻田土壤气体排放规律:CO2累积排放量,2种生物炭处理(DY-300、DY-700)后,土壤CO2累积排放量与对照组(DY-0)基本保持一致,并无显著差异;N2O累积排放量,DY-0>DY-700>DY-300,DY-300处理组的氮抑制性能最优;CH4累积排放量,DY-0>DY-700>DY-300,DY-300处理组的甲烷抑制性能最优。
2.3 微生物群落多样性分析
丹阳实验大田中采集到水稻土壤,选择通用引物,以16S rRNA基因为标靶,通过高通量测序分析土壤微生物群落,研究在不同温度条件制备的水稻秸秆生物炭对土壤微生物的群落结构的影响,水稻生长的不同时期对土壤微生物的群落结构的影响。
2.3.1 微生物丰度与多样性指数分析
土壤样品DNA高通量分析,选用Alpha多样性分析,统计结果如表4所示。
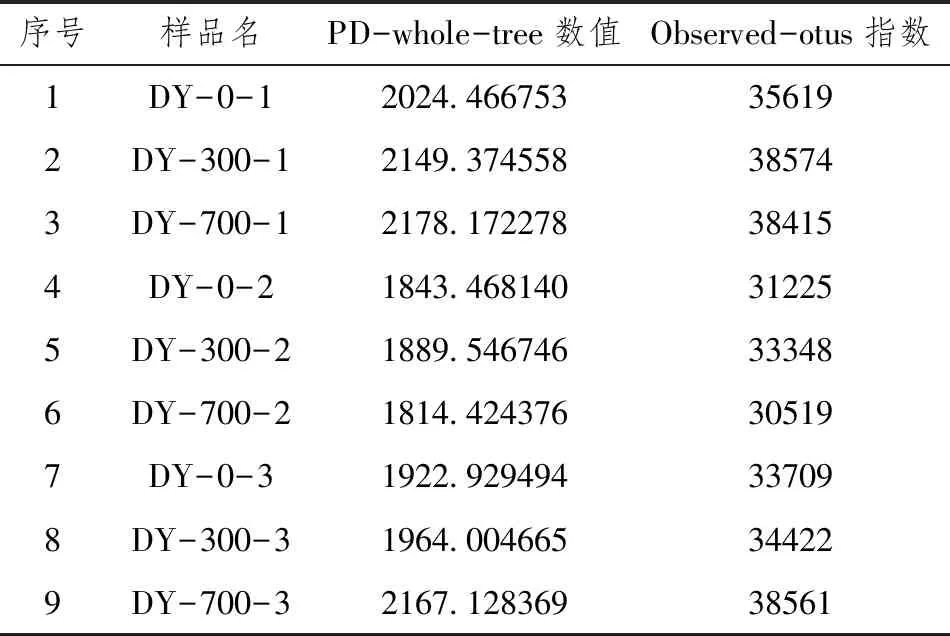
表4 Alpha(PD-whole-tree、observed-otus)多样性统计
如表4、图6、图7所示,不同条件生物炭处理对土壤微生物多样性指数的作用差异性是不同的。从PD-whole-tree指数分析,水稻生长前期多样性比较为PD whole tree(DY-700-1)>PD whole tree(DY-300-1)>PD whole tree(DY-0-1);水稻生长中期多样性比较为PD whole tree(DY-300-1)>PD whole tree(DY-0-1)>PD whole tree(DY-700-1);水稻生长期多样性比较为PD whole tree(DY-700-1)>PD whole tree(DY-300-1)>PD whole tree(DY-0-1)。从Observed OTUs指数分析,分析结果同PDwholetree指数分析比较,在水稻生长前期Observed OTUs(DY-300-1)>Observed OTUs(DY-700-1)>Observed OTUs(DY-0-1)。Shannon指数分析结果同PD-whole-tree指数分析分析结果相似。由于不同多样性指数算法存在差异,所以不同自变量组之间的多样性指数是具有一定差异的。从chao1指数看,探究同一时期物种丰度,水稻生长前期和后期均为Chao1(DY-700)>Chao1(DY-300)>Chao1(DY-0),水稻生长中期为Chao1(DY-300)>Chao1(DY-700)>Chao1(DY-0)。上述图表均表明,生物炭的添加有效提高了种群多样性和丰度,在水稻不同生长时期作用具有差异性。
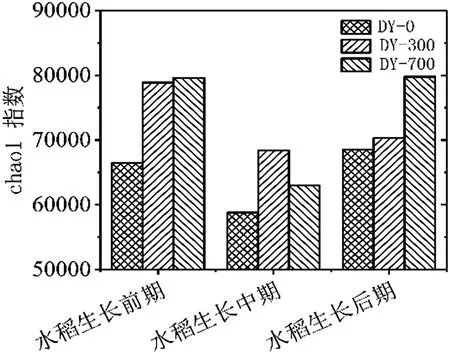
图6水稻不同生长时期chao1指数分析
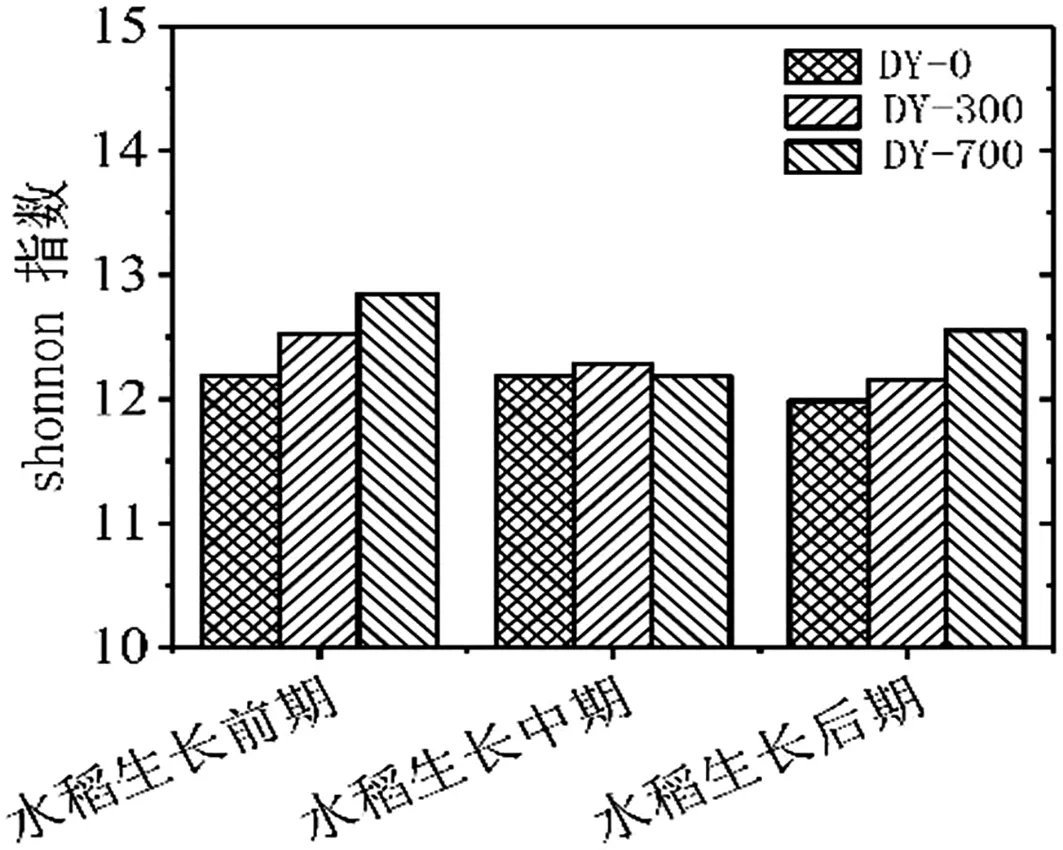
图7 水稻不同生长时期Shannon指数分析
2.3.2 微生物相对丰度分析
如图8所示,土壤中微生物浮霉菌门(Planctomycetes)丰度差异较小,为2.3%±0.2%。变形菌门(Proteobacteria)是测序微生物中的优势菌种,相对丰度值为40.7%~55.2%,其性状表现为水稻种植前期不同处理的优势度DY0-1>DY700-1>DY300-1、种植中期样本间优势度DY700-2>DY300-2>DY0-2、种植后期样本间优势度DY0-3>DY300-3>DY700-3;硝化螺旋菌门(Nitrospirae)作为硝化细菌的大类,可将亚硝酸盐氧化成硝酸盐,是土壤中氮循环的重要成员,相对丰度值为1.7%~8.9%,水稻生长前期硝化螺旋菌门丰度DY300-1>DY700-1>DY0-1、水稻生长中期相对丰度DY-0-2>DY300-2>DY700-2、水稻生长后期相对丰度DY300-3>DY700-3>DY0-3,且水稻水稻生长后期硝化螺旋菌门的种群相对丰度明显高于前中期。不同生物炭处理,不同时期的水稻微生物Phylum水平相对丰度是明显发生改变的,同一时期增添生物炭的水稻田微生物群落结构具有相对优势。
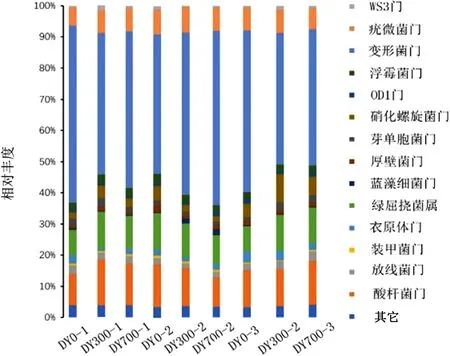
图8 土壤微生物相对丰度

图9 Phylum水平物种丰度热图
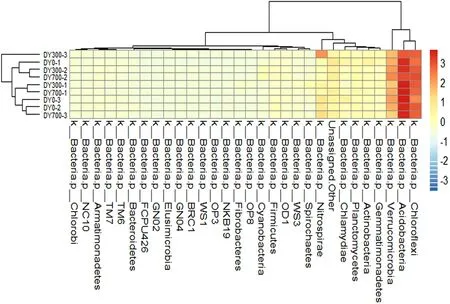
图10 Phylum水平物种的聚类分析
如图10所示,土壤样品中细菌微生物丰度Phylum水平(除变形菌门外)前30个进行分层聚类分析。聚类结果表明,DY300-2、DY700-3和DY0-1土壤样品集聚在一起,DY300-1、DY700-1土壤样品集聚在一起,DY0-2、DY0-3和DY700-3集聚在一起。生物炭的增添为水稻田原著微生物和菌落生存提供了有利条件,这种优良的环境条件致使土壤微生物的种类和丰度发生改变,从而改变了不同时期、不同处理的土壤微生物聚类。
2.4 土壤微生物群落结构与温室气体排放的相关性分析
在DY实验大田中,经过不同温度条件炭化的生物炭处理后,土壤微生物群落结构产生不同的响应。分析在不同土壤微生物群落结构与CH4、N2O、CO2排放通量量变化关系。通过IBM SPSS Statistics 20软件分析温室气体排放量和微生物两者之间的相关性。
2.4.1 温室气体排放与细菌Phylum水平相对丰度的关系
下述分析中细菌Phylum物种水平相对丰度均高于0.3%。
将CO2排放通量与细菌门类相对丰度进行相关性分析,如表5所示。CO2排放通量与衣原体门相对丰度呈显著负相关(p<0.05);CO2排放通量与OD1门相对丰度呈显著负相关(p<0.05);CO2排放通量与浮霉菌门、螺旋体门相对丰度呈显著正相关(p<0.05);OD1门为基因在NCBI库中未检索到。此外,经研究发现部分的CO2排放可能来源于水稻作物的呼吸作用。

表5 CO2排放通量与细菌Phylum水平物种相关性

表6 N2O排放通量与细菌Phylum水平物种相关性
将N2O排放通量与细菌门类相对丰度进行相关性分析,如表6所示。N2O排放通量与FCPU426门相对丰度呈显著正相关(p<0.05);N2O排放通量与SC4门相对丰度呈显著负相关(p<0.05)。FCPU426门、SC4门为基因序列在NCBI基因库中未检索到对应的细菌门。

表7 CH4排放通量与细菌Phylum水平物种的相关性
将CH4排放通量与细菌门类相对丰度进行相关性分析,如表7所示。CH4排放通量与硝化螺旋菌门、OD1门相对丰度呈显著负相关(p<0.05)。
2.4.2 温室气体排放与细菌Genus水平相对丰度的关系
下述分析中细菌的Genus物种水平相对丰度均高于0.1%。
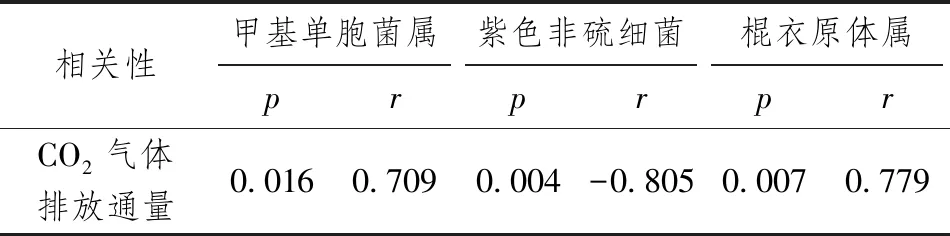
表8 CO2排放通量与Genus水平物种相关性
将CO2排放通量与细菌属类相对丰度进行相关性分析,如表8所示。CO2排放通量与甲基单胞菌属、棍衣原体属呈显著正相关性(p<0.05);CO2排放通量与紫色非硫细菌呈高度负相关性(p<0.05)。

表9 N2O排放通量与Genus水平物种相关性
将N2O排放通量与细菌属类相对丰度进行相关性分析,如表9所示。N2O排放通量与泉发菌属、甲基球菌属呈高度正相关性(p<0.05);N2O排放通量与甲基单胞菌属、甲基弯曲菌属呈显著正相关性(p<0.05);N2O排放通量与紫色非硫细菌呈显著负相关性(p<0.05)。

表10 CH4排放通量与Genus水平物种相关性
将CH4排放通量与细菌属类相对丰度进行相关性分析,如表10所示。CH4排放通量与泉发菌属、甲基球菌、甲基杆菌属、地发菌属呈显著正相关性(p<0.05);CH4排放通量与甲基单胞菌属呈高度正相关性(p<0.05);CH4排放通量与紫色非硫细菌呈高度负相关性(p<0.05);CH4排放通量与脱硫杆菌属呈显著负相关性(p<0.05)。
甲基球菌属、甲基单胞菌属、甲基杆菌属3种菌属都与CH4排放通量呈正相关性。由于上述3种菌属菌是以短链有机碳(甲烷、甲醇、甲醛等)作为生命质C源,因此土壤中甲基球菌属、甲基单胞菌属、甲基杆菌属的相对丰度可以表征土壤中CH4排放浓度,但是这种表征方式时效性差,具有一定的延迟性。甲基球菌属、甲基单胞菌属、甲基杆菌属之间属于竞争关系。
甲基单胞菌属、紫色非硫细菌、泉发细菌属和甲基球菌属4种菌与CH4、N2O、CO2排放通量之间的联系紧密。其中,甲基单胞菌属、甲基球菌属为异养性微生物,呼吸作用排放CO2,两者均是以CH4作为有机碳源,因此当CH4长期高浓度排放时,甲基单胞菌属、甲基球菌属的相对丰度会上升,此外,2种菌属与N2O排放量呈正相关性,说明这2种菌属在生命活动过程中释放N2O。而紫色非硫细菌为自养型微生物,能进行光合作用,固定CO2,从CH4角度分析,可能是CH4的存在对该菌种的生长产生了抑制作用。泉发菌属为兼性自养型微生物,其相对丰度同CH4、CO2呈正相关性,由于其本身的性质,本试验无法确定是高浓度的CH4、CO2排放导致泉发菌属相对丰度增加,还是泉发菌属相对丰度上升导致了CH4、CO2排放的增加,因此还需进一步试验进行验证。
3 讨论
3.1 生物炭的增添可抑制水稻田N2O、CH4排放,但对CO2排放作用不明显
DY水稻田间添加生物炭基肥和未添加生物炭基肥对水稻田中N2O、CH4气体的累积排放具有显著的差异性。从N2O排放通量上来看,水稻生长前期,DY0-1处理排放量是DY300-1处理排放量的18.3倍;水稻生长中期,DY0-1处理排放量是DY300-1处理排放量的5.0倍;水稻生长后期,DY0-1处理排放量是DY300-1处理排放量的1.5倍。DY0-1同DY700-1处理组之间的N2O排放通量差异性相对较小。CH4累积排放量在未经处理和经生物炭处理后并无显著性差异。
3.2 生物炭的增添可提高土壤微生物的多样性,在不同时期显现的功能性不同
DY水稻田中经过生物炭处理后的生物群落多样性显著高于未经生物炭处理的样本,在水稻生长前期和后期群落多样性呈现DY700>DY300>DY0,而在水稻生长中期群落多样性呈现DY300>DY700>DY0。从时间上分析,水稻生长前期和后期的群落多样性明显高于生长中期的多样性。从相对丰度上分析,变形菌门、绿屈挠菌属、酸杆菌门在水稻田土壤中表现为优势菌门,其优势度分布为变形菌门>酸杆菌门>绿屈挠菌属。
3.3 甲基单胞菌属、紫色非硫细菌、泉发细菌属和甲基球菌属相对丰度对温室气体排放的具有显著相关性
DY水稻实验田中CO2、CH4、N2O排放通量与甲基单胞菌属、泉发菌属、甲基球菌、甲基弯曲菌属、紫色非硫细菌、甲基杆菌属、脱硫杆菌属、地发菌属、棍衣原体的相对丰度呈相关性。甲基单胞菌属丰度与3种气体的排放均呈正相关性;紫色非硫细菌丰度与3种气体的排放均呈负相关性;泉发细菌属丰度与CH4、N2O排放通量呈正相关性;甲基球菌丰度与CH4、N2O排放通量呈正相关性。甲基单胞菌属、紫色非硫细菌、泉发细菌属和甲基球菌丰度对土壤温室气体排放具有相对直接的影响作用。
猜你喜欢
分子催化(2022年1期)2022-11-02
农业灾害研究(2022年1期)2022-05-07
化学工业与工程(2022年1期)2022-03-29
中国化肥信息(2020年2期)2020-11-14
今日农业(2020年19期)2020-11-06
中国资源综合利用(2017年4期)2018-01-22
中国塑料(2016年2期)2016-06-15
安全(2015年7期)2016-01-19
化工进展(2015年6期)2015-11-13
应用化工(2014年11期)2014-08-16
