
1株产GES-1型碳青霉烯酶铜绿假单胞菌的全基因组测序分析*
2021-07-02 07:27郭庆昕蔡加昌周宏伟张嵘蒋华蔚胡燕燕泉州市正骨医院检验科福建泉州362000浙江大学医学院附属第二医院检验科血液科杭州30009
临床检验杂志 2021年5期
郭庆昕,蔡加昌,周宏伟,张嵘,蒋华蔚,胡燕燕(.泉州市正骨医院检验科,福建泉州362000;2.浙江大学医学院附属第二医院 a.检验科,b.血液科,杭州30009)
铜绿假单胞菌是重要的医院感染菌之一,可引起重症肺炎、败血症、烧伤伤口感染等。产碳青霉烯酶铜绿假单胞菌(carbapenem resistantPseudomonasaeruginosa, CRPA)可水解几乎所有可用的β-内酰胺类抗菌药物,对全球公众健康构成了严重威胁[1]。β-内酰胺酶可分A、B和D三大类,A类包括KPC、BEL、CTX-M、GES家族的成员,B类包括NDM、IMP家族和VIM家族,D类主要是OXA家族。GES类型的β-内酰胺酶在20世纪后期被发现,因其含有丝氨酸碳β-内酰胺酶的结构特征(即Cys69和Cys239之间的二硫键),故与KPC型一起被分在A类中[2]。blaGES基因容易发生突变,目前已鉴定出40个亚型[3],GES酶水解谱也不断扩大,从原先的超广谱β-内酰胺酶逐渐进化为碳青霉烯酶[4]。为充分阐明耐药菌的传播动态,本研究对1株产GES-1型碳青霉烯酶铜绿假单胞菌进行全基因测序分析,采用生物信息技术分析本菌的多位点序列分型(multisite sequencing typing, MLST)、毒力基因和耐药基因,并分析blaGES-1基因所处的周围环境。
1 材料与方法
1.1研究对象 研究菌株来自浙江大学医学院附属第二医院重症医学科1例重症肺炎患者,该患者住院期间多次发热,曾先后使用哌拉西林钠/他唑巴坦钠、比阿培南、左氧氟沙星等抗感染治疗。该菌株于患者入ICU 13个月后从合格痰标本中分离,经基质辅助激光解吸/电离飞行时间(MALDI-TOF)质谱及微量肉汤稀释法确定为CRPA,此后7个月中反复从痰标本中检出。应用哌拉西林/他唑巴坦、比阿培南等抗菌药物治疗,但患者肺炎持续加重,呼吸功能衰竭,最终死亡。
1.2主要仪器与试剂 MALDI-TOF质谱仪(德国Bruker公司),PCR扩增仪、凝胶成像系统(Bio-Rad公司),Illumina HiSeq X10测序平台(美国Illumina公司),3730测序分析仪(美国ABI公司),电泳仪(北京六一仪器厂);血琼脂平板(郑州安图生物公司),铜绿假单胞菌药敏试剂盒(珠海迪尔公司),引物(上海生工生物公司),dNTP mix、10×Buffer、Taq酶、加样缓冲液(大连宝生物公司),细菌全基因组提取试剂盒(广州Magen公司)。
1.3细菌鉴定 标本接种于血琼脂平板培养过夜,分离菌采用MALDI-TOF质谱进行菌种鉴定。
1.4抗菌药物敏感性试验 采用微量肉汤稀释法检测庆大霉素、阿米卡星、哌拉西林、哌拉西林/他唑巴坦、头孢他啶、头孢吡肟、氨曲南、亚胺培南、美罗培南、环丙沙星、左氧氟沙星、黏菌素、多黏菌素、头孢哌酮/舒巴坦的敏感性,以铜绿假单胞菌ATCC 27853作为质控菌株,结果判读参照据美国临床和实验室标准协会(Clinical and Laboratory Standards Institute,CLSI)M100-S28标准[5]。亚胺培南和/或美罗培南的最低抑菌浓度≥8 μg/mL时判定为CRPA,头孢哌酮/舒巴坦敏感性判读参照CLSI-M100-S28中头孢哌酮方案进行。
1.5全基因组测序及分析 使用细菌基因组提取试剂盒提取DNA,用TE缓冲液洗脱,用紫外分光光度计对所提取的DNA进行浓度测定,测定后的合格样品送北京安诺优达基因科技公司,经DNA片段化、末端补平、3′端加A、连接测序接头、PCR扩增与纯化等完成文库制备,然后采用Illumina HiSeq X10测序平台通过双末端150 bp测序读长策略进行高通量测序。原始序列通过SPAdes v3.11.1[6]进行组装,“cov-cutoff”参数选为“auto”,“phred-offset”参数选为“33”,并用RAST[7]和Prokka[8]进行注释,Prokka软件的“kingdom”参数选择“Bacteria”并且选用“metagenome”参数提高基因预测能力。然后,通过Kleborate和ResFinder[9]分析菌株的MLST、毒力基因和耐药基因,参数均采用软件默认的设置(threshold for ID%为90%,minimum length为60%)。该铜绿假单胞菌的全基因测序原始数据已提交NCBI网站(http:∥www.ncbi.nlm.nih.gov/sra),BioProject登记号:PRJNA648026,Biosample登记号:SAMN15616709。
1.6blaGES-1基因环境分析 全基因组测序分析显示blaGES-1基因位于NODE_219片段中,长度3 150 bp,上游为整合子1(intl1)部分序列,下游为插入序列(ISPa21e)的部分序列。为呈现intl1和ISPa21e的完整序列,各设计了3对引物对这2个基因进行PCR扩增,引物序列见表1,产物送上海生工生物工程公司采用3730测序分析仪测序,结果用DNAMAN软件拼接。

表1 intl1和ISPa21e扩增延伸引物
2 结果
2.1抗菌药物敏感性试验 根据CLSI-M100-S28判读标准,该菌对美罗培南、黏菌素和多黏菌素B敏感,对其他抗菌药物均为耐药。见表2。
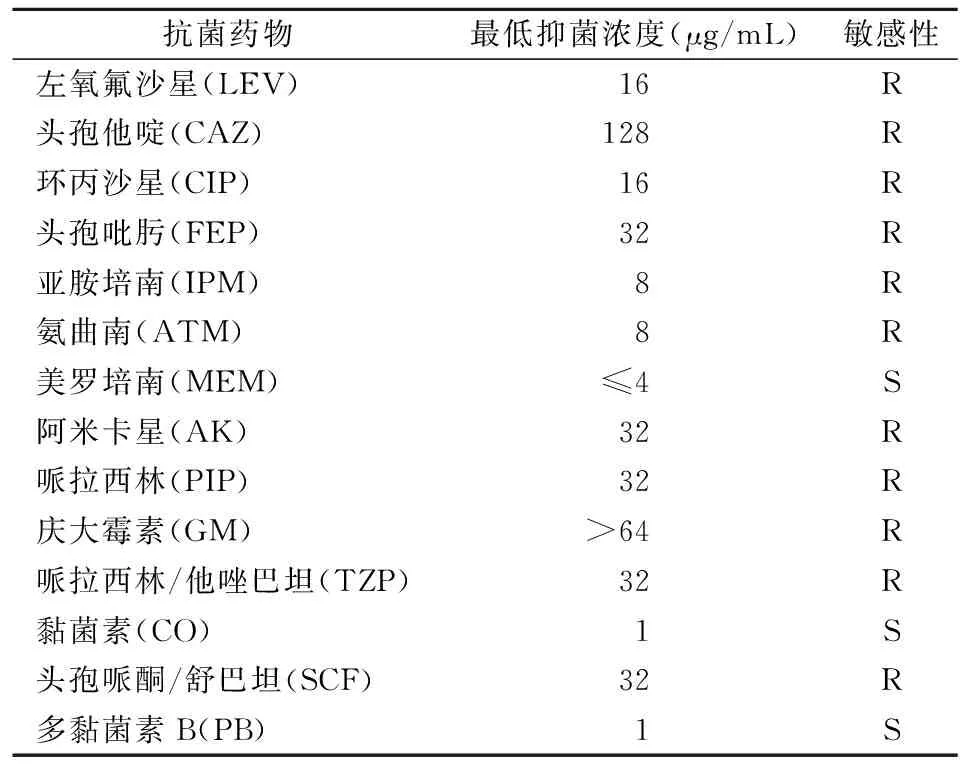
表2 抗菌药物敏感性试验结果
2.2全基因组测序结果 该菌全基因组测序分析结果显示,测序深度75×,基因组覆盖度100%。
2.2.1MLST及血清分型结果 经全基因组测序对7对管家基因(acsA、aroE、guaA、mutL、nuoD、ppsA、trpE)[10]进行分析,经https:∥cge.cbs.dtu.dk/services/MLST/查询,结果为(38-11-3-13-1-2-4),经比对,MLST分型为ST235型,血清分型为O11型。
2.2.2耐药基因测序结果 经ResFinder数据库比对,本株分离菌共携带有12个抗菌药物耐药基因,见表3。

表3 耐药基因测序结果
2.2.3blaGES-1基因比对结果及基因环境 全基因组测序分析发现,blaGES-1基因位于长度为3 150 bp的 NODE_219片段中。该基因片段与GenBank中的 KY860573菌株的72068—75217一致率为99.94%。blaGES-1基因上游1—192为整合子1(intl1)部分序列;372—1235为β-内酰胺类耐药基因blaGES-1,长度为864 bp,经ResFinder数据库比对,与登记号为HQ170511的菌株100%一致;下游依次为:1374—1928为氨基糖苷类/氟喹诺酮类耐药基因aac(6′)-Ⅰb3,长度为558 bp;2009—2253为无功能基因gcuE15,长度为245 bp;2261—3055为氨基糖苷类耐药基因aph(3′)-XV,长度为795 bp;3077—3150为插入序列ISPa21e部分序列。采用PCR扩增加测序对整合子intl1和插入序列ISPa21e进一步延伸,intl1全长为1 014 bp,ISPa21e全长为1 297 bp。blaGES-1基因环境见图1。

注:整合子1(intl1)长度1 014 bp,β-内酰胺类耐药基因(blaGES-1 )长度为864 bp,氨基糖苷类/氟喹诺酮类耐药基因[aac(6′)-Ⅰb3]长度为558 bp,无功能基因gcuE15长度为245 bp,氨基糖苷类耐药基因[aph(3′)-XV]长度795 bp,插入序列ISPa21e长度为1 297 bp。
2.2.4毒力基因测序结果 全基因组测序分析共查询到该菌包含有357个毒力基因,其中胞外酶3种,T3SS转录因子5种,胞外多糖20种,外毒素1种,溶血磷脂酶C 3种,溶血磷脂酶D 1种,鞭毛蛋白14种,鞭毛丝蛋白15种,菌毛蛋白25种,主要毒力基因见表4,该菌为exoU+/exoS-基因型。
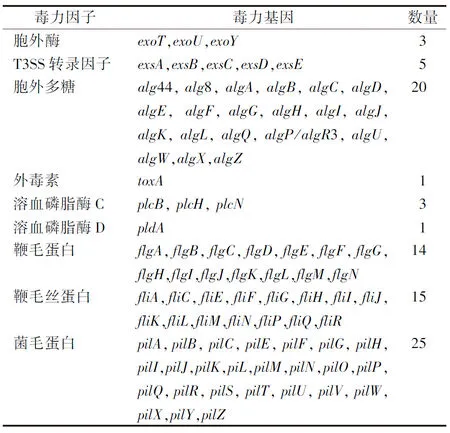
表4 本菌携带的主要毒力基因
3 讨论
铜绿假单胞菌的碳青霉烯酶编码基因(carbapenemase-encoding genes,CEGs)并非来源于其本身,而是水平基因转移获得的外源基因[11]。尽管其获得性CEGs的起源尚不清楚,但最有可能是来源于具有相同生态位的环境细菌[12]。以往认为GES酶水解碳青霉烯的能力很低,可被碳青霉烯类抗菌药物亚胺培南抑制,但GES酶的独特之处在于能通过单点突变将其水解谱扩展至碳青霉烯类[13]。
综合全球的多项研究,发现ST235、ST175和ST111是CRPA 3个主要的高风险克隆,而ST235分布最广,遍布全球五大洲,且ST235与O11型血清型相关[14]。Mulet等[15]研究了可能导致铜绿假单胞菌高风险克隆成功的生物标记,发现其在运动和产生色素上存在缺陷,但生物膜形成水平和自发突变频率增加,这些特征与高风险克隆引起的慢性感染互相印证。高风险克隆的耐药性与转移耐药和自发突变机制有关,其中ST235型中报道了近100个水平获得耐药元素,绝大多数产生金属β-内酰胺酶或超广谱β-内酰胺酶,其中GES酶最为多样化[16]。
本菌基因环境分析显示,在blaGES-1上游为Ⅰ类整合子(intl1),下游为耐药基因aac(6′)-Ⅰb3和aph(3′)-XV,右侧翼的插入序列(ISPa21e)。有研究发现CRPA中获得性碳青霉烯酶大多数存在于染色体位置tn402类转座子内的Ⅰ类整合子上[17]。整合子是编码产生整合酶,介导位点特异性重组,负责获得或切除载有抗菌药物耐药基因的基因盒,多个基因片段可能被同一整合子捕获到可变区域[18],这解释了本菌blaGES-1基因环境中3 150 bp的片段中集聚着3个重要的耐药基因,对β-内酰胺类、氟喹诺酮类、氨基糖苷类等抗假单胞菌抗菌药物耐药的原因。下游为IS110家族中的插入序列ISPa21e,该插入序列属于高度可移动性转座因子,是对基因组一个或多个靶位点具有插入能力的小分子片段,并可以移动邻近的基因[19]。该序列在染色体上的特定位置在不同的ST235分离株之间有所不同,表明在该克隆谱系的进化过程中发生了染色体重排[20]。在医院环境中,非发酵革兰阴性菌是这些元素的主要贮藏库,当转移到铜绿假单胞菌高风险克隆中时,会发生垂直扩增,导致CRPA的发生[21]。
经比对,本菌含有357个毒力基因,这些因子在细菌黏附、运动、生物膜形成、抗菌药物耐药和细胞毒性等不同环节中起作用。Ⅲ型分泌系统(T3SS)在铜绿假单胞菌生物膜形成、细胞毒作用中起重要作用,在感染过程中铜绿假单胞菌将毒素ExoS、ExoT、ExoU、ExoY注入到靶真核细胞的胞浆中,其中ExoU是一种磷脂酶效应细胞毒素,可致呼吸道、尿道、角膜和软组织等急性上皮损伤[22]。大多数铜绿假单胞菌均可检出ExoY和ExoT,而ExoS和ExoU则相对立,exoU-/exoS+基因型通常与侵袭表型相关,会导致宿主细胞的缓慢死亡;exoU+/exoS-基因型与细胞毒性高的菌株相关,导致宿主细胞快速而稳定的裂解,经常与慢性感染相关。高风险克隆ST235均为exoU+/exoS-基因型,与其他克隆相比,其临床效果差,预后不良[23]。ExsA、ExsB、ExsC、ExsD、ExsE特别是ExsA,是T3SS基因表达的主要调控因子,鞭毛丝蛋白(FliC)、胞外多糖(alg)等是T3SS的其他效应子。外毒素、溶血磷脂酶C、D是重要的细胞毒素,也可导致宿主细胞裂解。
临床早期发现高风险克隆对医院感染控制具有重要意义,有助于避免其在医院环境中扩散。全基因组测序在多重耐药菌的医院感染调查中提供基因分型、耐药基因、毒力基因等多种重要信息,在充分阐明耐药菌的传播动态,了解高风险克隆成功的因素等方面发挥重要作用。
猜你喜欢
现代医学与健康研究电子杂志(2022年6期)2022-11-26
浙江临床医学(2022年8期)2022-11-22
传染病信息(2022年2期)2022-07-15
系统医学(2022年10期)2022-07-05
中国兽医学报(2022年4期)2022-06-17
现代临床医学(2022年3期)2022-06-06
天津农业科学(2022年5期)2022-05-31
河北医科大学学报(2022年2期)2022-03-15
江苏农业科学(2019年14期)2019-09-23
江苏农业学报(2019年1期)2019-09-10
