
甜玉米种质资源种子性状全基因组关联分析
2021-06-30 02:23徐运林房浩周柏宇易月明王长进程昕昕余海兵
江苏农业学报 2021年2期
关键词:甜玉米
徐运林 房浩 周柏宇 易月明 王长进 程昕昕 余海兵


摘要: 粒長、粒宽、粒厚、百粒质量是影响玉米籽粒产量的重要性状,本研究主要统计分析了100份甜玉米种质资源的粒长、粒宽、粒厚和百粒质量等4个种子性状,并利用37 297个单核苷酸多态性(SNP)标记位点关联分析了控制甜玉米种子性状的重要位点。结果表明,100份甜玉米种质资源种子性状具有丰富的遗传多样性,性状变异幅度比较大,其中粒长为4.65~10.43 mm,粒宽为5.13~9.42 mm,粒厚为2.04~7.50 mm,百粒质量为4.51~20.09 g;表型变异分析发现,百粒质量的变异系数最大,为32.60%,粒宽的变异系数最小,为10.61%,且4个籽粒性状的频率分布曲线基本符合正态分布。相关性分析结果表明,粒长、粒宽、粒厚与百粒质量呈极显著正相关,粒长与粒厚呈极显著负相关。全基因组关联分析共检测到控制4个种子性状的75个SNP标记位点,其中有17个SNP标记位点与粒长性状有关,24个SNP标记位点与粒宽性状有关,11个SNP标记位点与粒厚性状有关,23个SNP标记位点与百粒质量性状有关。
关键词: 甜玉米;籽粒性状;单核苷酸多态性(SNP);全基因组关联分析(GWAS)
中图分类号: S330;S513 文献标识码: A 文章编号: 1000-4440(2021)02-0289-07
Abstract: The grain length, grain width, grain thickness and hundred-grain weight are important traits affecting the yield of maize. Four characters of seeds such as grain length, grain width, grain thickness and hundred-grain weight of 100 germplasm resources of sweet maize were summarized and analyzed in this study, and 37 297 single nucleotide polymorphism (SNP) loci were used to make correlation analysis on the important loci controlling the grain traits of sweet maize. The results showed that the grain traits of 100 germplasm resources of sweet maize had abundant genetic diversity with a relatively large variation range. The grain length was 4.65-10.43 mm, the grain width was 5.13-9.42 mm, the grain thickness was 2.04-7.50 mm and the hundred-grain weight was 4.51-20.09 g. Analysis on the phenotypic variation revealed that, the variable coefficient of the hundred-grain weight was the highest, with a value of 32.60%, and the variable coefficient of the grain width was the lowest, with a value of 10.61%. The distribution curves of frequencies of four grain traits were basically conformed to normal distribution. Results of correlation analysis showed that, the grain length, grain width and grain thickness were positively correlated with the hundred-grain weight very significantly, the grain length was negatively correlated with the grain thickness very significantly. A total of 75 SNP loci controlling the traits of four seeds were detected through genome-wide association study. Among the 75 SNP loci, 17 SNP loci were related with the grain length, 24 SNP loci were related with the grain width, 11 SNP loci were related with the grain thickness and 23 SNP loci were related with the hundred-grain weight.
Key words: sweet maize;grain traits;single nucleotide polymorphism(SNP);genome-wide association study(GWAS)
甜玉米(Sweet corn)是一种重要的果蔬作物,又称甜质型玉米 (Zea mays),其籽粒中含有较高含量的水溶性多糖、粗脂肪、淀粉、多种游离氨基酸和维生素[1],主要是由胚乳中su1、su2、sh1、sh2、sh4等控制糖分的基因发生突变获得的材料[2]。
种质资源是研究生物遗传的重要材料,利用种质资源多样性,可使群体内有更多的重组交换,创造新的种质类型,进而在分子水平上精确定位,也可检测多个目标性状和等位基因,为后续的育种工作提供研究基础[3-5]。关联分析以连锁不平衡为基础,将变异基因整合到一起,可分为全基因组关联研究(GWAS)和候选基因关联分析[6]。全基因组关联研究指用一定数目的单核苷酸多态性(SNP)分子标记对目标群体中的各个试验材料进行全基因组范围内的检测分析,再用相关软件对分子标记多态性和表型性状的数据进行关联分析,从而得到与表型性状变异相关联的分子标记[7]。近年来,GWAS分析已在水稻[8]、亚麻[9]、小麦[10]、大麦[11]等作物上得到广泛应用。
SNP(Single nucleotide polymorphism)即单核苷酸多态性,主要是指基因组上单个核苷酸发生的突变 [5]。SNP以较高的遗传稳定性、数量大、代表性强、易于实现自动化分析以及性价比高的特点迅速替代SSR成为新一代分子标记。史亚兴等[12]利用1 031个SNP标记将39份甜玉米自交系划分为5个类群;Bel等[13]利用8 590个SNP标记定位到控制玉米籽粒脂肪酸含量的候选基因fad2;2013年,Yang等[14]利用527份玉米自交系关联定位到控制玉米籽粒油分的74个显著位点;Rashid等[15]利用368个玉米自交系关联定位到高粱霜霉病的26个SNP位点。而有关甜玉米种子粒型性状关联分析的研究鲜见报道,因此,本研究利用安徽科技学院玉米工程技术研究中心100份甜玉米种质资源统计分析粒长、粒宽、粒厚和百粒质量4个性状,并利用37 297个SNP分子标记进行GWAS分析,为解析控制甜玉米种子性状的位点提供理论支持。
1 材料与方法
1.1 试验材料
本研究主要采用2018年、2019年安徽科技学院凤阳校区雷达山实验基地所种植的100份甜玉米自交系育种资源进行试验。100份甜玉米材料为M610(红)、F91GT8-4-1、14T4、F910GT8-4-1/Q267、1029黑-1-2-3、泰鲜泰-1、MS10-1、Q8(父)、3102(母)、YNT48、YNT6-2、9357、 Q7(母)、15NKY238、圣甜169-2、宫玉甜7号-3、15美玉甜007、FPT161、3174、YNT7-1(白)、斯达甜221-1、云甜王9号-10、YNT11、上海T、YNT50-2、YNT19-2、E134(白)、金甜2号(母)、秦甜98-4、MS9-1、F910GT8-4-1、E52(白)、E66-2、16粤甜16号、15HT0213X夏王、A38-11、16H沪甜2号、晶甜-1-1、E46-1、12HT022、TBF3109、E36、云南玉9号-4、16筑甜213、筑甜213-亲、BF2101(白)、E27-2、夏王(母)、广(甜糯)07、Q11(母)、Q10(父)、15白甜-1-1、T18(母)、E23-1、HT060、FT11、SH-X、T20嘉(母)、16黃T、M610(黄)、14花甜、YW1602、尖叶团粒、FT4、XSJD10、广甜()-2-2、黑CSB(母)、16HT01(父)、金甜2号(父)、黄甜-21-8、Q267、HT2、T16(母)、奥弗兰(SO)、引外甜、3176、BF3109、14WTX10、仲鲜甜-2、628(父)、夏王(父)、GS1-1、1048、19FT135、夏王(母)、TB010-3、广甜2号、T358(母)、14ATQT、T4、12HT00-1-黑、华珍(母)、2497黄、F910GT8-4-1/Q267、12HT073-4-1(黄)、12HT073-2、Q267/夏王、14T2-1、广甜(父)、粉红甜-1-2。
1.2 试验方法
1.2.1 测定方法 在甜玉米自交系果穗吐丝前进行套袋,抽雄散粉后人工辅助授粉,待成熟后,收获,晒干,脱粒,取3个果穗中部种子混合备用,分别称量其种子的百粒质量,设置3次重复,再从中各取20粒大小、形状相似,健康饱满的种子,测量粒长、粒宽、粒厚(使用游标卡尺),最后以每次重复的平均值作为所调查籽粒性状的值。
1.2.2 表型变异分析和相关性分析 利用SPSS Statistics 22统计分析软件和Excel 2016统计分析100份甜玉米自交系的粒长、粒宽、粒厚和百粒质量4个种子性状,计算最大值、最小值、平均数、标准差、变异系数,绘制正态分布图,并对4个性状进行相关性分析。
1.2.3 全基因组关联分析 采用TASSEL 2.1软件中的一般线性模型(Q,GLM),分别对4个种子性状进行全基因组关联分析,推断出SNP标记与各个性状之间存在的关联性。
2 结果与分析
2.1 100份甜玉米材料的群体结构分析
选取过滤后的SNP标记进行群体结构分析(图1)发现:当K为2时,对应的△K最大,然后△K迅速下降并趋于平缓,因此可以将这100份甜玉米材料划分为2个亚群,其中亚群1包括92份种质,亚群2包括8份种质。
2.2 表型变异分析
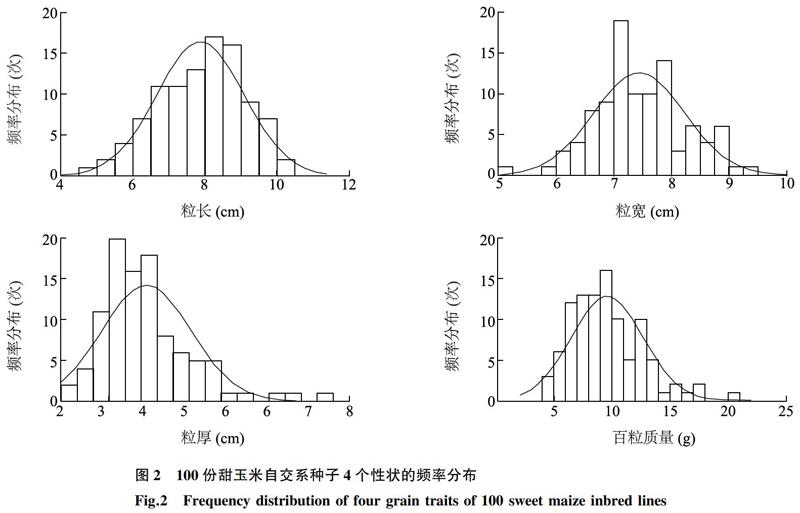
从表1可知,100份甜玉米种子的粒长、粒宽、粒厚和百粒质量等4个性状的差异比较大,其中粒长的变幅为4.65~10.43 mm,均值为7.87 mm,标准差为1.22 mm;粒宽的变幅为5.13~9.42 mm,标准差为0.79 mm;粒厚的变幅为2.04~7.50 mm,标准差为0.93 mm;百粒质量的变幅为4.51~20.09 g,标准差为3.12 g;百粒质量的变异系数最大(32.60%),粒宽的变异系数最小(10.61%)。从图2来看,4个性状的频率分布曲线基本符合正态分布,表现出数量性状特征,说明粒长、粒宽、粒厚和百粒质量的遗传受多基因控制,进一步表明100份甜玉米自交系具有较丰富的遗传多样性,可为甜玉米育种提供丰富的亲本材料。
2.3 相关性分析
100份甜玉米自交系种子的粒长、粒宽、粒厚和百粒质量等性状的皮尔逊相关性分析结果表明,百粒质量与粒长、粒宽、粒厚均表现为极显著正相关,粒长与粒厚表现为极显著负相关,粒长、粒厚与粒宽的相关性均不显著(表2)。
2.4 关联分析
通过对甜玉米56 000个SNP标记中连锁不平衡强度较高(R2>0.3)的标记进行过滤,得到了37 297个SNP位点,共检测到75个与粒长、粒宽、粒厚和百粒质量有关的显著位点,其中控制粒长的有17个显著关联位点,控制粒宽的有24个显著关联位点,控制粒厚的有11个显著关联位点,控制百粒质量的有23个显著关联位点。
2.4.1 粒长 如表3所示,在-lgP>3水平下,利用GLM-Q方法分析得出位于甜玉米1、3、4、8、10号染色体上的17个SNP位点与粒长性狀显著关联,其中在-lgP>5水平下,位于1号染色体上第19 611 619个碱基上的Affx-11533是与粒长性状极显著关联的SNP位点,贡献率最大,为25.49%。
2.4.2 粒宽 如表4所示,利用GLM-Q方法在-lgP>3水平的GWAS分析结果表明,24个SNP位点与粒宽性状相关联,分别位于第1、2、3、4、5、8、9、10号染色体上。其中在-lgP>4水平下,位于1号染色体上第244 631 581、289 538 157个碱基上的Affx-91280、Affx-11533的2个SNP位点与粒宽性状极显著关联,贡献率分别为18.46%、17.39%。
2.4.3 粒厚 如表5所示, 在-lgP>5水平的GWAS分析结果表明,11个SNP位点与粒厚性状相关联,分别位于第2、3、4、10号染色体上,其中在-lgP>6水平下,检测到5个SNP位点与粒厚性状极显著相关,分别位于3号染色体上第140 868 078、88 384 601、140 285 367、57 762 013个碱基上的Affx-90839、Affx-90505、Affx-90178、Affx-90075处和第4号染色体第200 963 685碱基上的Affx-90110,贡献率分别为27.97%、27.53%、25.79%、25.36%和 25.37%。
2.4.4 百粒质量 如表6所示,在-lgP>4水平下,利用GLM-Q方法分析得出23个SNP位点与百粒质量性状相关联,分别位于甜玉米的第2、3、4、6、7、8、9号染色体上,其中位于7号染色体上第129 236 674、123 110 014、149 940 420个碱基上的Affx-90265、Affx-91170、Affx-90113 3个SNP位点与百粒质量性状极显著相关,贡献率分别为25.27%、24.77%、22.13%。
3 讨论
3.1 100份甜玉米种质资源4个籽粒性状的表型变异分析和相关性分析
李用财[16]的研究结果表明,玉米产量与百粒质量呈极显著正相关,即通过提高百粒质量可以有效提高玉米产量;李春雨[1]研究得出133份甜玉米自交系百粒质量的均值为26.31 g,变幅为13.28~45.12 g。本研究结果表明,100份甜玉米种质资源种子4个性状分布曲线基本符合正态分布,表现出数量性状特征,说明粒长、粒宽、粒厚和百粒质量4个籽粒性状的遗传受多基因控制。相关性分析结果表明,甜玉米粒长、粒宽、粒厚与百粒质量呈极显著正相关;粒长与粒厚呈极显著负相关,与粒宽无显著相关性;粒宽与粒厚无显著相关性。因此,100份甜玉米自交系4个籽粒性状间差异较大,具有较丰富的遗传多样性,可为甜玉米育种提供丰富的亲本材料。
3.2 100份甜玉米种质资源4个种子性状的全基因组关联分析
前期一些学者们利用SNP位点得出控制小麦、水稻、玉米等主要作物粒型的主要位点[17-20],其中,赵檀[17]检测到9个与小麦百粒质量相关联的位点,Fang等[19]检测到8个与水稻粒型相关联的位点,Hu等[20]对282份玉米自交系用2 271 584个SNP标记检测到17个耐冷性遗传位点,李春雨[1]用一般线性模型(GLM)和混合线性模型(MLM)检测到位于第1、5和7号染色体上的9个与百粒质量相关联的SNP标记位点。杨宁[5]检测到9个与玉米粒宽性状相关联的SNP标记位点。本研究经GWAS分析共获得75个控制甜玉米种子性状的SNP标记位点,其中控制粒长的SNP标记位点有17个,控制粒宽的SNP标记位点有24个,控制粒厚的SNP标记位点有11个,控制百粒质量的SNP标记位点有23个, 在候选区间内共有231个基因,找到96个候选基因参与玉米种子生长发育,这些基因主要与玉米赤霉素(GAs)、生长素(IAA)、油菜素内酯(BRs)的生物合成和信号转导途径有关。
参考文献:
[1] 李春雨.利用染色体片段代换系及关联群体鉴定甜玉米产量性状遗传位点[D].广州:华南农业大学,2018.
[2] CHENG X X,XIONG F, WANG C J, et al.Seed reserve utilization and hydrolytic enzyme activities in germinating seeds of sweet corn[J]. Pakistan Journal of Botany,2018, 50(1):111-116.
[3] 张秀荣.花生关联分析群体构建及抗旱相关分子标记研究[D].泰安:山东农业大学,2017.
[4] 唐富福. 水稻农艺和品质若干性状的全基因组初步关联分析[D]. 杭州:浙江大学, 2013.
[5] 杨 宁.玉米17个农艺性状的全基因组关联分析及Mo17和大刍草基因组组装[D]. 武汉:华中农业大学,2016.
[6] 向星洁.基于候选基因标记的香菇重要农艺性状关联分析[D].武汉:华中农业大学,2015.
[7] 李桂冬.小麦3个重要农艺性状的SSR标记全基因组关联分析[D].杨凌:西北农林科技大学,2015.
[8] 侯青青,司丽珍,黄学辉,等. 水稻复杂性状研究的新途径:水稻重要农艺性状全基因组关联分析[J].生命科学,2016,28(10):1250-1257.
[9] BRAULIO J, SOTO C, FEMANDO W, et al. Assessing the agronomic potential of linseed genotypes by multivariate analyses and association mapping of agronomic traits[J].Euphytica,2014(5):35-49.
[10]WANG G, JEFFREY M, LEONARDJ Z, et al. Marker-trait association analysis of kernel hardness and related agronomic traits in a core collection of wheat lines[J].Molecular Breeding, 2014, 177-184.
[11]张 绵. 西藏野生大麦干旱耐性基因等位多态性分析及相关基因的功能鉴定[D].杭州:浙江大学,2017.
[12]史亚兴,卢柏山,宋 伟,等. 基于SNP标记技术的糯玉米种质遗传多样性分析[J]. 华北农学报, 2015,30(3):77-82.
[13]BEL A, ZHENG P, LUCK S, et al. Whole genome scan detects an allelic variant of fad2 associated with increased oleic acid levels in maize[J]. Mol Geneet Genomics, 2008, 279:1-10.
[14]YANG Q, LI Z, LI W Q, et al. CACTA-like transposable element in ZmCCT attenuated photoperiod sensitivity and accelerated the postdomestication spread of maize[J]. Proc Natl Acad Sci USA, 2013, 110:16969-16974.
[15]RASHID Z, SINGH P K, VEMURI H, et al. Genome-wide association study in Asia-adapted tropical maize reveals novel and explored genomic regions for sorghum downy mildew resistance[J].Scientific Reports, 2018(8):366.
[16]李用财. 东北中熟玉米杂交组合主要农艺性状鉴定[D].哈尔滨:黑龙江大学,2018.
[17]赵 檀. 河北省小麦品种遗传多样性及主要农艺性状的关联分析[D].保定:河北农业大学,2014.
[18]么大轩,张 彬,刘松涛,等.基于SNP和SSR对甜玉米种质遗传多样性的评价[J]. 江苏农业科学,2019,47(7):45-50.
[19]FANG C, XING Y, MAO H, et al. GS3,a major QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transnenbrane protein[J].Theoretical & Applied Genetics,2006,112(6):1164-1171.
[20]HU G, ZHAO L, LU Y, et al. Genome wide association study identified multiple genetic loci on chilling resistance during germination in maize[J].Scientific Reports, 2017, 7(1):10840.
(責任编辑:陈海霞)
猜你喜欢
农民致富之友(2020年28期)2020-10-20
吉林蔬菜(2020年4期)2020-09-10
国际种业前沿动态(2020年8期)2020-03-09
农民致富之友(2019年5期)2019-03-13
农家科技下旬刊(2018年12期)2018-02-25
农民致富之友(2017年12期)2018-01-31
农民致富之友(2017年23期)2018-01-02
农业与技术(2016年8期)2016-10-21
农家科技(2009年4期)2009-04-23
