
尾叶桉与窿缘组树种间杂交种的生长特征
2021-06-29 20:52莫继有兰俊罗建中吴满芬彭智邦
广西植物 2021年4期
关键词:生长
莫继有 兰俊 罗建中 吴满芬 彭智邦


摘 要: 选定组配树种对桉树杂交种的特性起决定作用,但不同种间杂交种的生长表现常常不容易预测。为掌握华南地区桉树重要树种间杂交种的生长特征,促进桉树杂交育种的精准化,该文以父本为混合花粉的人工杂交种为遗传材料,以尾叶桉×巨桉为对照,进行了尾叶桉与桉树窿缘组树种(布拉斯桉、细叶桉、钝盖赤桉、昆北赤桉)的4种组配杂交种的生长特征研究。结果表明:在材积生长量上,尾叶桉与窿缘组树种杂交种低于尾叶桉×巨桉,但其内部树种间差异显著(P<0.05),其中尾叶桉×昆北赤桉具有显著优势;尾叶桉与窿缘组树种杂交种的树高、胸径均显著小于尾叶桉×
巨桉,但其高径比显著大于尾叶桉×巨桉(P<0.05);尾叶桉与窿缘组树种杂交种具有高的和一致的保存率,且在不同组配间、家系间(组配内)均无显著差异,5年生值为84.4%~89.6%;尾叶桉与窿缘组树种杂交种的5年生材积的平均变异系数约为64%,组配间、家系间差异大,都大于尾叶桉×巨桉。尾叶桉与窿缘组树种杂交种的速生性、树形等与尾叶桉×巨桉间存在显著差异,尾叶桉与窿缘组树种杂交种的組配、家系间具有显著的生长差异,可为桉树遗传改良提供丰富的多样性。
关键词: 种间杂交种, 生长, 尾叶桉, 窿缘组, 混合花粉子代
中图分类号: Q945
文献标识码: A
文章编号: 1000-3142(2021)04-0631-09
Abstract: Deciding crossing parental species will determine characteristics of eucalypt hybrids, but the growth performance of hybrids from different parents is hard to predict. Understanding the growth features of hybrids between important eucalypt species in South China will greatly improve hybrid breeding accuracy. We studied the growth characteristics of hybrids between Eucalyptus urophylla and four Section Exsertaria species/subspecies (E. brassiana, E. tereticornis, E. camaldulensis var. obtusa and E. camaldulensis subsp. simulata) with E. urophylla × E. grandis (UG) hybrids serving as a control, of which the male parents were polymix of 10 plus trees were studied. The results were as follows: Stem volume growth in E. urophylla × Section Exsertaria species (UES) hybrids were significantly lower than UG. And there were significant differences between the UES hybrids (P<0.05), among which E. urophylla × E. camaldulensis subsp. simulata demonstrated significant advantages. DBH and Ht of UES were significantly lower than those of UG, but with height-diameter ratios were significantly higher than UGs (P<0.05), with 5-year-old values approximately 150 and 130, respectively. UES had high and uniform preserve rates, and no significant differences due to either species or family (within species) level. Their 5-year-old values were 84.4%-89.6%. The UESs coefficient of variation (CV) of 5-year-old stem volume had an average of 64%, yet significantly differed between species was higher than that of the UGs. UES was significantly different from UG on volume growth and tree shape, and the significant differences between UES species and families could provide substantial diversity to eucalypt genetic improvement.
Key words: interspecific hybrids, growth, Eucalyptus urophylla, Section Exsertaria species, polymix progeny
桉树种间杂交因可以聚合多个树种的特性,甚至获得杂种优势,在桉树育种中广泛应用,特别是在亚热带、热带桉树的遗传改良中尤其成功(Potts & Dungey,2004),是目前世界上桉树人工林大国如巴西、中国、南非等的主要育种方法(罗建中,2012;Kullan et al.,2012;Madhibha et al.,2013)。种间杂交过程是不同树种基因的重组,会对一系列数量、质量等性状产生不同程度的影响,产生不同于亲本的新表现型。虽然杂交种的性状大多介于亲本之间(Potts & Dungey,2004),但在特性不同的树种进行交配时,其子代的生长特点并不容易预测,例如,木材生长量、用材率等经济性状,有很多杂种优势的结论,但也有杂种劣势的结果(Assis,2000;Butcher & Williams,2002)。有研究发现杂交种的生存能力有随着亲本分类学距离的增加而降低的趋势、杂交种往往比纯种更容易遭受虫害(Dungey et al.,2000;Potts & Dungey,2004);一些起初正常的杂交种植株,到2年生后才会表现出异常、陆续死亡(Potts & Dungey,2004);在桉树杂交种叶片特征的研究中发现,尾叶桉×粗皮桉叶片属偏父本型,而尾叶桉×细叶桉(Eucalyptus urophylla × E. tereticornis)的多偏向于母本(齐杰等,2018)。因而有专家认为杂交育种的亲本选择,总体处在“边试边看”的状态,缺乏有力的科学理论依据(Grifin et al.,1988)。
然而,在有着明确目标的杂交育种中,需要对将产生的杂交子代的重要性状有较把握的预期,在还没有完善的理论指导之前,就要求对将选用的亲本及其杂交子代的特性有较充分的了解。
桉树是一个拥有近1 000个树种的大家族,遗传多样性极其丰富,不同树种在生长、适应等多方面存在不同程度的差异。在众多的桉树中,双蒴盖亚属的横脉组(Section Transversaria)和窿缘组(Section Exsertaria)树种具有极高的重要性,它们都是包含数十个树种大属,其中很多是世界重要用材树种。已有研究发现,这两个组的树种都具有独特的性质,横脉组的尾叶桉(Eucalyptus urophylla),是仅有的两个天然分布在澳大利亚以外的树种之一,它对我国华南南部的气候适应性强,速生性和抗病虫害能力优良、无性繁殖能力强;同属横脉组的巨桉(E. grandis),是著名的速生树种,具有树干通直圆满、木材性质优良等优点,但在热带地区容易遭受病害(王豁然,2010;罗建中,2012)。窿缘组的树种,如赤桉(E. camaldulensis)、细叶桉(E. tereticornis)等,抗风能力突出,具有良好的耐旱、耐瘠薄能力强等特点,但它们之间在适生环境、速生性、干形特点上具有一定差异(Madhibha et al.,2013)。这两个组的很多树种,不但作为纯种广泛种植,还是重要的杂交亲本树种,它们的组内、组间杂交还产生了大量优异的杂交种,如尾叶桉×巨桉(E. urophylla × E. grandis)、尾叶桉×赤桉(E. urophylla × E. camaldulensis), 细叶桉×巨桉(E. tereticornis ×E. grandis),是多个国家桉树杂交育种的主要组配类型(Retief & Stanger,2009;罗建中,2012;Kullan et al.,2012; Madhibha et al.,2013)。
然而,虽然桉树的很多种间杂交种得到广泛應用,但却很少见到不同组间树种杂交种生长、适应性特点的系统报道。无论是对桉树组间杂交的遗传变异特性的认识,还是对杂交育种中亲本树种的选择都缺乏系统的依据。这可能主要因为桉树人工杂交难度大、成本高,组间交配效果的研究需要大量平衡设计的杂交子代,传统的育种项目往往难以承受如此大的操作规模(Eldridge,1993)。虽然桉树杂交育种的研究和应用历史已经不短,已有不少关于桉树种间杂交的遗传特点研究的报道(Assis,2000;Verryn,2000;Verry,2000;吴昆明等,2001),但研究的组配常常缺乏系统性,涉及的杂交树种少,参与的亲本数量有限,所得结果的公允性受限。混合花粉授粉对解决这一问题有独特的优势。
混合花粉授粉指将多个单株的花粉混合、向指定的隔离母本授粉,一般认为这种授粉方法可以改善亲本间的亲和力,提高杂交的可孕性,并且因为得到与多个父本的交配机会,可以有效提高育种效率(曾贞等,2008)。这种授粉方式具有高的交配效率,在林木树种中的杨树、海岸松、湿地松等多个树种中,曾通过混合花粉子代测定来研究种子园、杂交种的遗传特性等(Bridgwater et al.,1998;Wheeler et al.,2006;Vidal et al.,2015)。随着低成本分子技术应用于识别混合花粉授粉子代中不同父本,可准确估算不同亲本的育种值,有人甚至认为混合花粉授粉可成为传统的全同胞育种方法的可能替代方法(Lambeth et al.,2001)。应用混合花粉子代的分析,加拿大的科学家还提出了一种称作“BWB”(breeding without breeding)的育种方法(Elkassaby & Lstib,2009)。
为了解华南地区桉树重要树种间杂交种的生长特征,本研究测定了杂交子代林的1年生、2年生、5年生和9年生的生长数据,分析了尾叶桉与窿缘组树种杂交子代在幼龄期、成熟龄期的材积生长性状、环境适应性等特点,旨在加深对桉树不同组树种间杂交种生长特征的理解,为科学制定桉树杂交育种策略的提供理论依据。
1 材料与方法
1.1 遗传材料
所研究的杂交种来自5个亲本树种,其中母本为横脉组的尾叶桉,父本为窿缘组4个树种,分别是细叶桉、钝盖赤桉(E.camaldulensis var. obtusa )、昆北赤桉(E.camaldulensis subsp. simulata)、布拉斯桉(E. brassiana)。同时,以巨桉作为对照父本,因为尾叶桉×巨桉被认为是该地区最优秀的树种组合(Pegg et al.,2006)。所有选作杂交亲本的单株都来自广西国有东门林场,是各树种子代或无性系测定林中的优良单株、优良无性系。用作母本的尾叶桉为7株没有亲缘关系的单株,而每个父本均为各树种10个优良单株花粉的混合体。根据树种杂交的组配方式,所有杂交种被分为5种组配类型,每种组配类型含3~5个家系,共19个家系(表1)。
1.2 试验地与试验林
用于杂交种子代测定试验的植株定植于1991年4月,试验地在国有广西东门林场渠多分场内(22°24′ N、107°54′ E),海拔约150 m,当地属南亚热带季风气候,年平均气温21.7 ℃,极端最高温41 ℃,极端最低温-0.1 ℃,年无霜期346 d,年降雨量 1 000~1 300 mm,相对湿度74%~83%,夏季有台风。
试验采用随机完全区组设计,单行10株小区,4次重复,株行距2.0 m × 3.0 m。试验地较为平坦,坡度2°~5°。土壤为砖红壤性红壤,造林前备耕采用机耕全垦整地,带翅犁松土两次,深度为25~30 mm。造林26 d后施肥,N、P、K施肥量均为50 kg·hm-2。肥料为广西南宁产复合肥,含N、P、K各10%。施肥时,在距离苗木两边各约30 cm处挖两个小穴、施放肥料,然后覆土。
1.3 试验林调查方法
试验林定植后,每年调查生长数据,调查的性状包括存活状况(保存率Survival rate, Sur)、树高(Ht)及胸径(DBH)。本研究选用的是1.5年生、2.0年生、5.0年生和8.8年生数据,为便于叙述在文中分别称其为1年生、2年生、5年生和9年生。胸径的测量工具为钢围尺,数据精确到0.1 cm;树高的测量工具为1年生、2年生用测高杆,5年生后用超声波测高测距仪(Vertex Ⅲ,瑞典生产),数据精确到0.01 m。
1.4 数据统计与分析方法
(1)数据分析模型如下。
(2)高径比=Ht(cm)/DBH(cm)。
(3) 材积计算采用如下公式。
式中:Vol为单株材积(dm3);π为圆周率;DBH为树木胸高处直径(cm);Ht为树高(m)。
数据分析采用SAS软件(The SAS system for Windows, release 8.01);数据处理时,个别残差过大的数据视作植株缺失,未参加分析。
2 结果与分析
为方便起见,本研究5种类型的杂交组配类型及简称分别为尾叶桉×布拉斯桉(UB)、尾叶桉×钝盖赤桉(UCo)、尾叶桉×昆北赤桉(UCs)、尾葉桉×细叶桉(UT)、尾叶桉×巨桉(UG)。下面分别从杂交组配水平、组配内的家系水平对杂交种的生长特点进行分析。
2.1 材积生长量分析
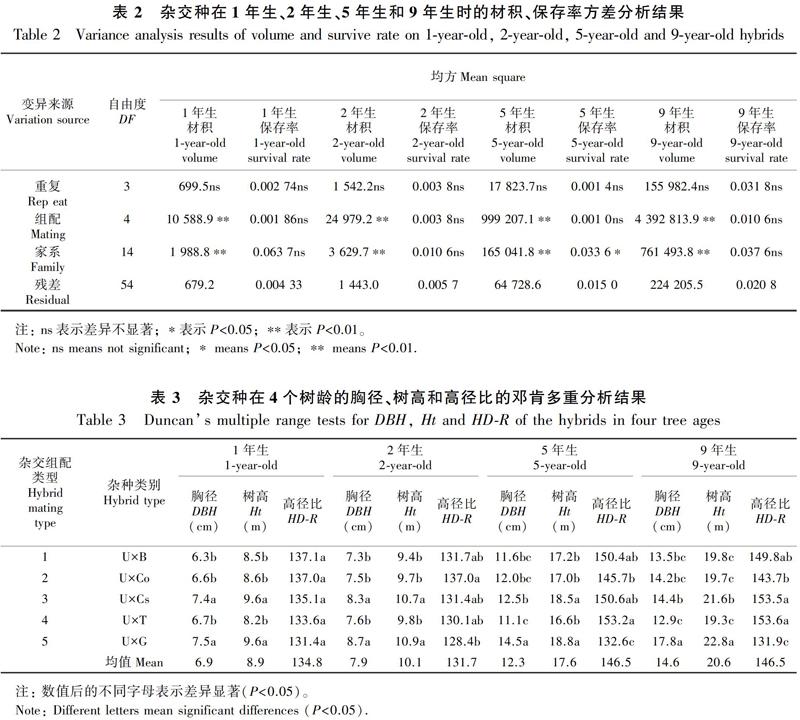
材积常作为林木产量的衡量指标,也是比较品种速生性的最重要指标。方差分析显示(表2),所研究的5类杂交组配在4个林龄的材积生长量均存在极显著差异(P<0.01),表明尾叶桉与不同类型桉树的杂交种间具有显著的材积生长差异,而且这个差异从第1年到第9年一直存在。在杂交种的家系水平(组配内),材积的差异也均达到极显著水平(P<0.01),表明即使在同样的树种组配内,不同个体产生的杂交家系间生长量仍存在极显著的差异。
在树种组配水平,尾叶桉×巨桉(UG)的材积生长量在4个树龄都具有优势,且显著高于尾叶桉与窿缘组树种的所有组配(图1)。尾叶桉与窿缘组树种的4种组配的材积生长量差异特征随树龄有一定变化:在幼林期(1年生和2年生),材积的大小排序为尾叶桉×昆北赤桉(UCs)>尾叶桉×细叶桉(UT)>尾叶桉×钝盖赤桉(UCo)>尾叶桉×布拉斯桉(UB),尾叶桉×昆北赤桉显著大于其他3个组配(P<0.05),但其他3个组配间无显著差异;而在成熟树龄(5年生和9年生),材积的大小依次为UCs>UCo>UB>UT,幼龄期在4个组配中处于第2位的UT,掉到了所有组配的最后一名,其他组配的相对位置没有改变,仍然是UCs具有显著的材积优势(P<0.05,分析结果未列出)。
从幼龄期到成熟龄期,尾叶桉与窿缘组杂交种间的材积差异呈扩大趋势。与尾叶桉×巨桉相比,2年生时,尾叶桉×昆北赤桉、尾叶桉×细叶桉、尾叶桉×钝盖赤桉和尾叶桉×布拉斯桉分别是它的87%、68%、68%和64%;而5年生时则分别是它的70%、51%、65%和60%。相对来看,各尾叶桉×窿缘组杂交种间的生长量差异较为稳定。
分析发现,尾叶桉×窿缘组树种的不同组配内家系间,在各个树龄的材积生长量差异一直达到极显著水平。以5年生材积生长量为例,尾叶桉×布拉斯桉、尾叶桉×钝盖赤桉、尾叶桉×昆北赤桉、尾叶桉×细叶桉中,材积最大家系与最小家系材积比分别是1.25、1.73、1.28和1.52;作为对照的尾叶桉×巨桉组配中,这个比值达1.82。作为混合花粉的家系,表现的是母本的一般配合力,这个结果说明各组配内的不同母本的配合力具有显著差异。
2.2 树高、胸径和高径比的分析
树高、胸径以及它们的比值(高径比)是反映树木生长特点的重要性状,且一起决定树木的材积生长量。高径比的大小还影响树种的稳定性和抗逆能力。表2结果发现,5种杂交种的树高、胸径以及高径比在各树龄都存在显著差异(P<0.05)。在所有4个树龄,尾叶桉与窿缘组间4类杂交种的胸径、树高均小于尾叶桉×巨桉组配,其内部4类杂交种间的大小排序与材积的类似。在1年生和2年生时,胸径、树高排序为尾叶桉×昆北赤桉>尾叶桉×细叶桉>尾叶桉×钝盖赤桉>尾叶桉×布拉斯桉;而到了较大树龄(5年生和9年生),尾叶桉×细叶桉的生长速度降低而排到所有杂交种的最后,其他杂交种间的相对顺序未变。但是,与材积不同的是,在1年生和2年生时尾叶桉与窿缘组杂交种中尾叶桉×昆北赤桉的胸径、树高并不显著低于尾叶桉×巨桉。2年生胸径和树高分别为8.3 cm、10.7 m,和同龄尾叶桉×巨桉的8.7 cm和10.9 m极为相近,它们间没有显著差异。随着树龄的增长,尾叶桉与窿缘组杂交种与尾叶桉×巨桉间的差距扩大,5年生和9年生的树高、胸径都显著小于尾叶桉×巨桉。
尾叶桉与窿缘组杂交种高径比的特点则完全不同于材积生长量等性状(表3)。在1年生和2年生时,4类组配的高径比无显著差异,两树龄分别在133.6~137.1和130.1~137.0之间,其中尾叶桉×钝盖赤桉的两个树龄的高径比均为137.0;而同龄尾叶桉×巨桉的分别为131.4和128.4,略小于尾叶桉与窿缘属杂交种,1年生时它与尾叶桉×窿缘组树种都无显著差异,仅在2年生时显著小于其中的尾叶桉×钝盖赤桉。5年生和9年生时,4种尾叶桉×窿缘组树种间的差异都达到显著水平(P<0.05)。5年生时,高径比的大小和排序为尾叶桉×细叶桉(153.2)>尾叶桉×昆北赤桉(150.6)>尾叶桉×布拉斯桉(150.4)>尾叶桉×钝盖赤桉(145.7)。9年生时的数值与5年生相近,排序也完全相同。尾叶桉×巨桉杂交种的同龄高径比分别为132.6和131.9,都极显著小于尾叶桉与窿缘组杂交种(P<0.01)。另外,也可见高径比从幼龄期到成熟龄期有10%幅度的增长,但在幼龄期相近、成熟龄期也稳定,所有杂交种1年生、2年生、5年生和9年生的平均高径比为134.8、131.7、146.5和146.5。
在组配内的家系水平,树高、胸径的差异特征与组配水平极相似。虽然在各树龄都达到显著水平,但在1年生和2年生时的差异显著低于5年生和9年生;同时,组配内家系水平的变异幅度均低于组配水平(结果未列出),这表明组配水平变异是变异的主要来源。组配内家系的高径比不同,在所有4个树龄,组配内家系间差异均未达到显著水平,表明组配内具有高度的一致性。
2.3 保存率分析
本研究的试验地气候具有冬季温暖干燥、夏季高温高湿的特点,夏季台风较频繁,桉树病害、风害较为常见(罗建中等,2009),保存率可作为品种的环境适应性、抗逆性的重要判断指标。从表2可以看出,保存率在杂交组配水平、家系水平几乎均无显著差异(仅5年生家系间差异显著)。这表明尾叶桉与窿缘组树种的杂交种、尾叶桉×巨桉,无论在组配水平、组配内家系水平,对试验地点的幼龄期、成熟龄期的环境适应性都极其相近。
4种尾叶桉与窿缘组树种杂交种的1年生、2年生、5年生和9年生平均保存率分别为96.0%、94.5%、87.0%和79.5%。雖然随着树龄的增长引起保存率会下降,但是都属类似地区相应树龄的较高保存率,说明这4种杂交种具有良好的环境适应性。同龄的尾叶桉×巨桉的保存率依次为93.8%、93.1%、83.1%和77.5%,都略低于尾叶桉×窿缘组树种,每个年度的差异都未达到显著水平。从图2可以看到,这4种杂交种及尾叶桉×巨桉的保存率间的差异随着树龄的增大也略有增大。值得一提的是,成熟龄期的(5年生、9年生)尾叶桉×昆北赤桉的保存率分别为89.6%和83.3%,都是同龄期的最高值。
虽然家系间(组配内)保存率的差异小,但是随着树龄增大,组配内的家系间保存率差异幅度也变大。在5年生、9年生时,平均保存率最高的尾叶桉×昆北赤桉的家系水平变异范围为81.3%~96.9%和71.9%~90.6%(结果未列出),家系间最大差异分别达到15%和20%,存在着值得关注的遗传变异。
2.4 变异系数分析
从表3可以看出,4种尾叶桉×窿缘组树种的变异系数总体上随着树龄增大而增大,在幼龄期、成熟龄期相对稳定,1年生、2年生、5年生和9年生的平均值分别为48.9%、49.5%、64.2%和68.3%。4种杂交组配的变异系数差异极大,以5年生为例,最大值和最小值分别为53.2%和78.1%,它们分别来自尾叶桉与不同赤桉的组配,其中最小值为尾叶桉×昆北赤桉,最大值为尾叶桉×钝盖赤桉;尾叶桉×巨桉的变异系数为59.6%,略小于4种尾叶桉×窿缘树种的平均值。
混合花粉授粉可能带来丰富的变异,更应反映在家系水平的变异系数上。混合花粉子代的多样性会直接体现在家系内单株的多样性上。对各组配内家系水平材积变异系数差异进行分析(表4),结果发现,组配内的家系间差异也极大(结果未列出)。以5年生材积为例,尾叶桉×窿缘组树种的家系水平变异系数分布在47.5%~96.7%之间。其中,平均变异系数最大的组配(尾叶桉×钝盖赤桉)的家系间差异也最大,分布在59.2%~96.7%之间;平均变异系数最小的组配(尾叶桉×昆北赤桉)的家系间差异也最小,其变异系数在47.5%~56.4%之间。而同龄的尾叶桉×巨桉组配的平均值为59.6%,家系的分布范围在47.4%~80.5%之间,比4种尾叶桉×窿缘组树种的平均值低约5%。
同一试验中的单父本尾叶桉×巨桉家系(Fam-U×G)1年生、2年生材积变异系数比同龄的低约5%和10%,而5年生和9年生的变异系数分别低约20%和15%,也略小于尾叶桉×窿缘组树种中变异系数最小的家系(表4)。同时,它的变异系数也小于尾叶桉×巨桉混合花粉组配的平均变异系数,与混合花粉尾叶桉×巨桉的最小家系值相近。可见,尾叶桉×窿缘组树种的混合花粉子代材积变异都高于尾叶桉×巨桉单父本家系。
3 讨论与结论
3.1 材积生长量
尾叶桉与窿缘组树种杂交种的材积生长量总体显著低于广为应用的尾叶桉×巨桉,使其在单纯作为木材生产上的竞争力受到限制。尾叶桉×昆北赤桉是在尾叶桉与窿缘组树种杂交种内具有显著优势的杂交组配,且在环境适应性上也具有一定优势,可在以后加大对其应用力度。其他组配间的材积生长量差异不显著,但所有组配内家系间均具有显著差异。家系水平的丰富变异使得尾叶桉与窿缘组树种杂交种可能产生大量材积生长量大的家系、个体,可在此水平上加大对其选择利用。从幼龄期(1年生和2年生)到成熟龄期过程中,尾叶桉与窿缘组树种杂交种间材积差异会发生一定变化,特别是尾叶桉×细叶桉的相对表现差异大,所以优良品种的选定须在幼龄期后进行,以增加可靠性。
尾葉桉和巨桉间杂交种材积生长的优异性,已经被多个国家研究报道(Retief & Stanger,2009;Kullan et al.,2012;Madhibha et al.,2013)。尾叶桉与窿缘组树种间的杂交种,以往研究相对较多的为尾叶桉×赤桉,在材积上也低于尾叶桉×巨桉(Assis,2000;Richard et al.,2006;李梅等,2007),这与本研究的结
果相似。虽然材积生长量没有优势,但以尾叶桉或巨桉作母本,赤桉、细叶桉为父本的杂交种在抗风、耐旱等特殊要求高的地区利用广泛(Assis,2000;Madhibha et al.,2013)。因此,在我国抗逆要求高的地区,应加大对尾叶桉与窿缘组树种间杂交种的测定,以充分认识它们抗逆特点、合理利用。
3.2 树高、胸径及高径比
虽然尾叶桉与窿缘组树种间杂交种的树高、胸径都显著小于尾叶桉×巨桉,但它们的高径比都显著高于尾叶桉×巨桉。同时,高径比在尾叶桉与窿缘组树种杂交种的组配间也存在显著差异,可以通过杂交亲本树种选择得到改良。高径比在抗逆性上的特殊意义,可能使尾叶桉与窿缘组树种间的杂交种具有特殊的优势,它们在我国台风高发地区的突出表现,已显示了它们的特殊价值(罗建中等,2009;何国华等,2014)。但是,它们5年生时高达近150的高径比与抗风的优异表现,却与欧洲树木的高径比在80以上则抗逆性差的结论不一致(Wonn & O′Hara,2001)。这可能是由于树种不同、灾害类型的不同所导致,但具体的原因还有待深入研究。高径比还是决定树木干形的关键值,是树木材积计算的关键参数,对树木的出材率、木材价值等有重要影响(Luo et al.,2013)。高径比对尾叶桉与窿缘组树种间杂交种木材价值的影响值得深入研究,以便于制定合理的改良、利用计划。
3.3 环境适应性
保存率的分析表明,尾叶桉×窿缘组树种都具有良好的环境适应性,且和尾叶桉×巨桉、尾叶桉×窿缘组树种均无显著差异。这说明尾叶桉×窿缘组树种的4类杂交种和尾叶桉×巨桉一样,都适合在试验地的环境中生长。本研究的试验地为典型的夏季湿热气候,夏季台风较频繁,这与之前的研究发现尾叶桉与赤桉、细叶桉等窿缘属树种杂交种抗风能力强,具有一定的适应性优势相一致(罗建中等,2009;何国华等,2014)。本研究4个树龄的研究结果发现,所有杂交组配类型的环境适应性表现较相近,未出现显著差异。不同地点、年份的气候条件会有较大的差异,可能是导致这种结果的主要原因,可综合多年、多地点的测定结果以得到更加全面的评价。
3.4 变异系数
目前在世界主要桉树生产国,热带桉树的杂交育种主要采用“有性改良,无性利用”的育种策略,即应用人工杂交、自由授粉等有性的方法创造变异,改良目标性状,再通过无性繁殖优异杂交子代个体的方法应用变异,形成优良品种。在这样的策略中,遗传变异往往比平均值更重要(Potts & Dungey,2004)。尾叶桉与窿缘组树种杂交子代的材积变异系数略高于尾叶桉×巨桉的子代,且均大于尾叶桉×巨桉的单父本子代,说明混合花粉子代的变异幅度得到显著提高,具有更大的应用价值。4类尾叶桉×窿缘组树种的材积变异系数有极大差异,其中与钝盖赤桉的最大、与昆北赤桉的最小,需根据不同组配的变异特点加以利用。本研究的尾叶桉与窿缘组树种杂交种5年生材积变异系数约63%,与李梅等(2007)开展的相似杂交种研究结果相近,可能变异系数的大小也受环境、遗传材料的影响。
本研究中的杂交种作为林木重要树种,在这里仅评价了部分生长性状,而对反映树木重要特征的树冠特点、反映树木重要价值的木材性质等性状均未涉及,对这些树种特征的理解仍有较大的局限性。希望以后的研究能弥补这些缺憾,形成对这些树种更加全面、深入的认识。
参考文献:
ASSIS T, 2000. Production and use of eucalyptus hybrids for industrial purposes[C]. Noosa, Queensland, Australia:Proceedings of QFRI/CRC-SPF Symposium: 63-74.
BOSELA M, KONOPKA B, SEBE VLADIMR, et al.,2014. Modelling height to diameter ratio—An opportunity to increase Norway spruce stand stability in the Western Carpathians[J]. Lesn Cas For J,60(1): 71-80.
BRIDGWATER FE, BRAMLETT DL, BYRAM TD, et al.,1998. Controlled mass pollination in loblolly pine to increase genetic gains[J]. For Chron, 74(2): 185-189.
BUTCHER PA, WILLIAMS ER, 2002. Variation in outcrossing rates and growth in Eucalyptus camaldulensis from the Petford Region, Queensland; Evidence of outbreeding depression[J]. Silv Gen, 512(21):485-488.
DEAN N, REBECCA J, 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae)[J]. Telopea, 21: 129-145.
DUAN MX, SONG TM, FAN HW, et al.,2000. Utilizing genetic markers mixing pollen to estimate heterosis value and xenia effect value of high oil corn[J]. J Chin Agric Univ, 5(3): 45-50.[段民孝, 宋同明, 范弘伟, 等, 2002. 用遗传标记混合花粉估计玉米杂交当代优势和油分花粉直感效应[J]. 中国农业大学学报, 5(3): 45-50.]
DUNGEY HS, POTTS BM, LI WF, 2000. Plant genetics affects arthropod community richness and composition: Evidence from a synthetic eucalypt hybrid population[J]. Evolution, 54(6): 1938-1946.
ELDRIDGE KG, DAVID J, HARWOOD CE, et al., 1993. Eucalypt domestication and breeding[M]. Oxford: Clarendon Press: 207-208.
ELKASSABY YA, LSTIBREK M, 2009. Breeding without breeding[J]. Tree Genet Genomes, 91(2): 111-120.
HE GH, CHEN WP, JIAN M, et al., 2014. Growth of Eucalyptus urophylla hybrid progeny in Leizhou Peninsula[J]. Eucalypt Sci & Technol, 31(2): 10-16.[何国华, 陈文平, 简明, 等, 2014. 尾叶桉杂交子代在雷州半岛的生长分析[J]. 桉树科技, 31(2): 10-16.]
HENRIK R, HALLINGBACK, GUNNAR J, 2013. Genetic information from progeny trials: A comparison between progenies generated by open pollination and by controlled crosses[J]. Tree Genet Genomes, 9(3): 731-740.
KULLAN ARK, DYK MMV, HEFER CA, et al., 2012. Genetic dissection of growth, wood basic density and gene expression in interspecific backcrosses of Eucalyptus grandis and E. urophylla[J]. BMC Genet, 13(1): 60-71.
LAMBETH C, LEE BD, WHEELER N, 2001. Polymix breeding with parental analysis of progeny: An alternative to full-sib breeding and testing[J]. Theor Appl Genet, 103(6-7): 930-943.
LI M, GAN SM, LI FG, et al., 2007. Selection of growth and bacterial wilt resistance in Eucalyptus interspecific hybrids[J]. J Nanjing For Univ (Nat Sci Ed), 31(6): 25-28.[李梅, 甘四明, 李發根, 等, 2007. 桉属种间杂种生长和抗青枯病的联合选择[J]. 南京林业大学学报(自然科学版), 31(6): 25-28.]
LIU CH, LIU Y, 2018. Phenotype analysis of pineapple hybrid line obtained by mixed-pollen cross and its paternal origin analysis[J]. J Agric, (4): 39-45.[刘传和, 刘岩, 2018. 菠萝混合授粉杂交品系表型研究及其父本来源分析[J]. 农学学报, (4): 39-45.]
LUO JZ, XIE YJ, CAO JG, et al., 2009. Genetic variation in 2-year Eucalypt hybrids growth and typhoon resistance[J]. Acta Pratacult Sin, 18(6): 91-97.[罗建中, 谢耀坚, 曹加光, 等, 2009. 2年生桉树杂交种生长与抗风的遗传变异研究[J]. 草业学报, 18(6): 91-97.]
LUO JZ, 2012. Genetic and environmental effect of eucalypt hybrid clones on pulpwood traits[D]. Nanjing: Nanjing Forestry University.[罗建中, 2012. 桉树杂种无性系纸浆材性状的遗传与环境效应研究[D]. 南京: 南京林业大学.]
LUO JZ, ARNOLD R, REN SQ, et al., 2013. Veneer grades, recoveries, and values from 5-year-old eucalypt clones[J]. Ann For Sci, 70(4): 417-428.
MADHIBHA T, MUREPA R, MUSOKONYI C, et al., 2013. Genetic parameter estimates for interspecific Eucalyptus hybrids and implications for hybrid breeding strategy[J]. New For, 44(1): 63-84.
NICHOLAS W, PEGGY P, VALERIE H, et al., 2006. Polymix breeding with paternity analysis in Populus: A test for differential reproductive success (DRS) among pollen donors[J]. Tree Genet Genom, 2(1): 53-60.
POTTS BM, DUNGEY HS, 2004. Interspecific hybridization of Eucalyptus: Key issues for breeders and geneticists[J]. New For, 27(2): 115-138.
QI J, LU WH, LI P, et al., 2018. Genetic variation patterns in leaf morphology on eucalypts and their hybrids[J]. J Trop Subtrop Bot, 26(6): 589-596.[齐杰, 卢万鸿, 李鹏, 等, 2018. 桉树及其杂交种叶片形态的遗传变异特征[J]. 热带亚热带植物学报, 26(6): 589-596.]
RETIEF E, STANGER TK, 2009. Genetic parameters of pure and hybrid populations of Eucalyptus grandis and E. urophylla and implications for hybrid breeding strategy[J]. S For, (2): 133-140.
RICHARD P, LI HJ, LI HW, et al., 2006. Eucalypt artificial hybridization research in Guangxi[J]. Guangxi For Sci, 35(4): 238-242.[RICHARD P, 李海佳, 李宏伟, 等, 2006. 广西桉树人工杂交育种研究[J]. 广西林业科学, 35(4): 238-242.]
TOLOZA AC, LUCIA A, ZERBA E, et al., 2008. Interspecific hybridization of eucalyptus as a potential tool to improve the bioactivity of essential oils against permethrin-resistant head lice from Argentina[J]. Bioresour Technol, 99(15): 7341-7347.
VERRYN SD, 2000. Eucalyptus hybrid breeding in South Africa[J]. Noosa, Queensland, Australia: Proceedings of QFRI/CRC-SPF Symposium: 191-199.
VIDAL M, PLOMION C, HARVENGTL, et al., 2015. Paternity recovery in two maritime pine polycross mating designs and consequences for breeding[J]. Tree Genet Genomes, 11(5): 105.
VERRY SD, 2000. Eucalyptus hybrid breeding in Congo[C]. Noosa, Queensland, Australia: Proceedings of QFRI/CRC-SPF Symposium: 14-26.
WANG HR, 2010. Introduction to eucalypt biology[M]. Beijing: Science Press.[王豁然, 2010. 桉树生物学概论[M]. 北京: 科学出版社.]
WHEELER N, PAYNE P, HIPKINS V, et al., 2006. Polymix breeding with paternity analysis in Populus: a test for differential reproductive success (DRS) among pollen donors[J]. Tree Gen Genomes, 2(1): 53-60.
WONN HT, O′HARA KL, 2001. Height: Diameter ratios and stability relationships for four northern rocky mountain tree species[J]. West J Appl For, 16(2): 87-94.
WU KM, WU JY, GAN SM, 2001. Hybridization and heterosis in eucalyptus[J]. Guangdong For Sci Technol, 17(4): 10-15.[吴坤明, 吴菊英, 甘四明, 2001. 桉树杂交育种及杂种优势的利用简介[J]. 广东林业科技, 17(4): 10-15.]
ZENG Z, LUO JW, YANG Y, et al., 2008. AFLP analyses of the Fl progenies of tea from a mixed-pollen cross[J]. J Hunan Agric Univ (Nat Sci Ed), 34(6): 719-723.[曾貞, 罗军武, 杨阳, 等, 2008. 混合花粉授粉茶树杂交F1代AFLP分析[J]. 湖南农业大学学报(自然科学版), 34(6): 719-723.]
(责任编辑 何永艳)
猜你喜欢
睿士(2020年11期)2020-11-16
中学生天地(A版)(2020年3期)2020-04-10
故事作文·高年级(2020年3期)2020-03-17
小资CHIC!ELEGANCE(2019年33期)2019-11-22
小资CHIC!ELEGANCE(2019年15期)2019-05-21
财经(2017年6期)2017-03-29
红领巾·萌芽(2016年1期)2016-09-10
课堂内外(高中版)(2016年5期)2016-07-11
课堂内外(小学版)(2016年4期)2016-07-04
英语学习·新锐空间(2013年3期)2013-05-23
