
黑肩绿盲蝽外生殖器及其感器的超微结构
2021-06-28 08:31杨韩英张振飞吴伟坚
环境昆虫学报 2021年3期
杨韩英,张振飞,张 扬,吴伟坚*
(1. 华南农业大学昆虫生态研究室,广州 510642;2. 广东省生物农药创制与应用重点实验室,广州 510642;3. 广东省农业科学院植物保护研究所,广州 50640)
黑肩绿盲蝽CyrtorhinuslividipennisReuter,属半翅目盲蝽科,杂食性,广泛分布于水稻栽培区,是水稻主要害虫稻飞虱重要捕食天敌,还可大量捕食叶蝉等的卵(陈建明等, 1992)。室内实验研究,单头黑肩绿盲蝽一生能取食飞虱卵214.9~222.4粒,在一定密度范围内,捕食量随密度提高而增加(吴光荣等, 1987),田间7月下旬至8月下旬是黑肩绿盲蝽的发生高峰期,蝽虱比达到或超过1 ∶1时对褐飞虱的种群发展有一定的控制作用(齐会会等, 2014)。
外生殖器是昆虫的生殖部位,一般雌虫外生殖器结构相对简单且缺乏差异性,而雄虫外生殖器是由不同的、单独进化部分组成混合特征,这使得其相较触角、足、复眼等其他身体部位,结构复杂且种间差异明显,因此,在分类近缘种时尤为重要(Arnqvist, 1998; Huber, 2003; Songaetal., 2010),常用于盲蝽科分类(Konstantinov, 2003)。另一方面,生殖器与性选择密切相关,性选择对生殖器进化有重要促进作用(Hoskenetal., 2004)。
感器是昆虫感知环境的重要门户,由表皮演变而来,连接神经元,分布在足、触角、生殖器等身体部位,具有机械感知、化学感知、嗅觉、味觉、温湿度感受等功能(Schenider, 1964)。感器在种类、数量、超微形态、触角神经叶等方面都具有性二型特征(秦玉川, 2009),且同型结构在不同种可能具有不同功能(Acebesetal., 2003)。关于外生殖器感器,在果蝇、石蛾、豆娘、步甲、寄生蜂、叶蝉等类群中有报道,并表明外生殖器感受器在感知并选择寄主上合适的产卵位点和求偶前期两性感受异性状态过程中起到作用(Acebesetal., 2003; Spänhoffetal., 2003; Kimetal., 2004; Uhíaetal., 2005; Hummeletal., 2006; Dwecketal., 2008)。
本文通过扫描电镜对黑肩绿盲蝽雌雄成虫外生殖器形态进行了观察,对生殖器上感器分类,结合黑肩绿盲蝽交配行为,探讨感器功能,从而为其进一步研究及利用提供参考。
1 材料与方法
1.1 供试昆虫
供试黑肩绿盲蝽来源于本实验室饲养种群,饲养于带有飞虱卵的水稻上,置于养虫笼内,养虫笼置于人工气候室内(温度28±1℃,RH70%±5%,光周期L ∶D=14 ∶10)。收集黑肩绿盲蝽雌雄成虫供试。
1.2 扫描电镜制样
雌雄成虫各20头,体视镜下取其腹部生殖节,部分雌成虫腹部解剖产卵器;样品浸入2.5%戊二醛溶液中,4℃条件下固定24 h,0.1 mol/L磷酸缓冲液(PBS)冲洗3次(40 min/次),而后置于1%锇酸固定2.5 h,0.1 mol/L PBS冲洗3次(10 min/次)。然后乙醇系列脱水,30%(4℃),50%(4℃),75%(4℃),80%,90%,各1次(10 min/次),100%浓度5次(10 min/次)。脱水后的样品置于临界点干燥仪(Bal-Tec CPD 030)中干燥2 h。取出样品,在体视镜下,按腹部正面、腹面、侧面分别有序粘贴在导电胶上,后喷金镀膜。最后,在扫描电镜(荷兰FEI Nova Nano SEM 430)下拍照观察。
1.3 感器分类和鉴定
感器的分类和鉴定参照Schenider(1964)和Zacharuk(1985)的方法。
2 结果与分析
2.1 雌成虫交配孔与产卵器的结构
黑肩绿盲蝽雌成虫交配孔(gonopore, gp; 图1-B)开口于第8、9腹节节间膜上,位于产卵器基部,覆盖于呈倒三角形的下生殖板下,交配孔狭长,孔内2种感器;产卵器(图1-B)由第1产卵瓣(first valvula, VL 1)、第2产卵瓣(second valvula, VL 2)、第3产卵瓣(third valvula, VL 3)构成。第1产卵瓣包围第2产卵瓣呈针状,边缘锯齿状结构反向排列,二者以第1产卵瓣上脊状条、第2产卵瓣上凹槽组成可滑动结构,内分布脊载鳞片(spine-bearing scales, SBS; 图3-F);第3产卵瓣为1对片状,靠虫体腹面和末端边缘密布感觉毛。
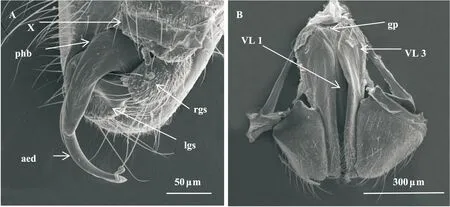
图1 黑肩绿盲蝽雌、雄成虫外生殖器扫描电镜观察Fig.1 SEM observations of genitial of female and male adults of Cyrtorhinus lividipennis注:A,雄虫交配器;B,雌虫产卵器及交配孔;X,第X腹节;VL 1,第1产卵瓣;VL 3,第3产卵瓣;gp,交配孔;phb,阳茎基;aed,阳茎;lgs,左侧突;rgs,右侧突。Note: A, Genitial of male; B, Ovipositor and gonopore of female; X, X abdominal segment; VL 1, first valvula; VL 3, third valvula; gp, gonopore; phb, phallobase; aed, aedeagus; lgs, left genital styles; rgs, right genital styles.
2.2 交配孔与产卵器上感器的类型与超微结构
2.2.1毛形感器TS(triochoid sensilla)
毛形感器I(triochoid sensilla I, TS I; 图3-E)分布于第3产卵瓣虫体腹面和末端边缘,基部宽1.68 μm,着生于突起的基窝中,感器表面有纵纹,端部尖细,整体细长,略弯曲,平均44.93 μm,最长可达77 μm,数量较多;毛形感器II(triochoid sensilla II, TS II; 图2-B)位于交配孔外缘及内部,短小,基部宽0.53 μm,长1.45 μm,表面光滑,分布密集,组成巢状。
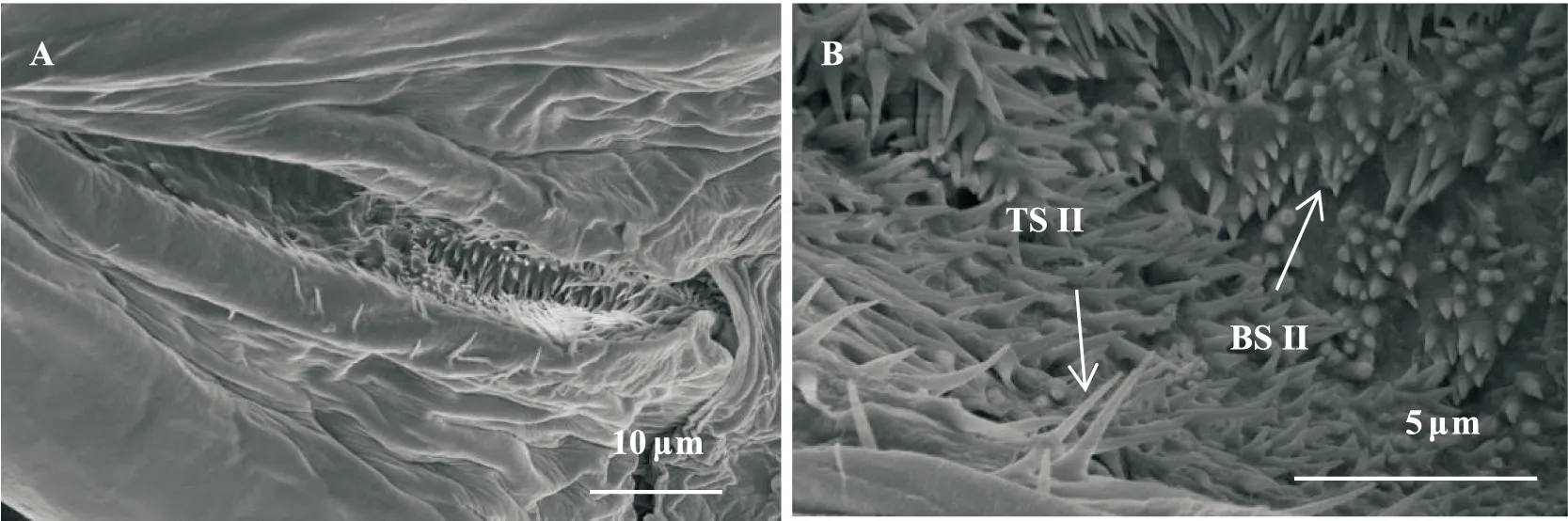
图2 黑肩绿盲蝽雌成虫交配孔中感器扫描电镜观察Fig.2 SEM observations of sensilla in the gonopore of female Cyrtorhinus lividipennis注:A,雌成虫交配孔;B,雌成虫交配孔中感器;TS II,毛形感器II;BS II,锥形感器II。Note: A, Gonopore of female; B, Sensilla in gonopore; TS II, Triochoid sensilla II; BS II, Basiconic sensilla II.
2.2.2刺形感器CHS (chaetica sensilla)
刺形感器(图3-E)在第3产卵瓣外缘密集分布,表面光滑,无孔无基窝,基宽0.51 μm,长6.12 μm,较尖细,多平直,数量众多。
2.2.3凹状感器DS(depression sensilla)
凹状感器(图3-D),为第2产卵瓣锯齿边缘两侧表皮上圆形浅凹,中央略突起或平坦,直径3 μm,单侧8~11个,排列较均匀。
2.2.4锥形感器BS (basiconic sensilla)
锥形感器I(basiconic sensilla I, BS I; 图3-C)仅位于第3产卵瓣中段,表皮光滑,整体笔直,长度20 μm,左右两瓣各一簇,单簇10~12个,整体向背侧倾斜。锥形感器II(basiconic sensilla II, BS II; 图2-B)位于交配孔底部,为表皮圆锥形突起,数量较多,基宽0.31 μm,长0.43 μm。

图3 黑肩绿盲蝽雌虫产卵器感器的扫描电镜观察Fig.3 SEM observations of sensilla on the ovipositor of female Cyrtorhinus lividipennis注:A,第3产卵瓣;B,第1产卵瓣和第2产卵瓣;VL 1,第1产卵瓣;VL 2,第2产卵瓣;TS I,毛形感器;CHS,刺形感器;BS I,锥形感器 I;DS,凹状感器;SBS,脊载鳞片。Note: A, First valvula; B, First valvula and second valvula; VL 1, First valvula; VL 2, Second valvula; TS I, Triochoid sensilla I; CHS, Chaetica sensilla; BS I, Bascionic sensilla I; DS, Depression sensilla; SBS, Spine-bearing scales.
2.3 雄成虫交配器的结构
黑肩绿盲蝽雄成虫交配器(图1-A)由阳茎(aedeagus, aed)和抱握器构成,整体呈不对称结构。阳茎单一圆筒状,顶端窄,斜悬于生殖腔孔中,并伸出生殖腔;基部即阳茎基(phallobase, phb),阳茎基着生于第9腹节;抱握器着生于生殖腔开口外后缘处,由左右两部分组成,分别为阳基左侧突(left genital styles, lgs),阳基右侧突(right genital styles, rgs),均为不规则片状,大小和形状均不对称,左侧突U形,右侧突椭圆形,且左侧突大于右侧突。射精管位于阳茎中,在交配时,从阳茎端伸出。阳茎基光滑无感器分布,左侧突外侧、右侧突外侧共有感器8种。
2.4 交配器上感器的类型与超微结构
2.4.1刺形感器CHS (chaetica sensilla)
刺形感器(图4-A)分布于右侧突外侧,左侧突尾端,在交配器上数量最多,密度最高,形态同产卵瓣上刺形感器。
2.4.2毛形感器TS (triochoid sensilla)
毛形感器I(图4-A),同雌虫毛形感器形态上相同,长度明显长于其他感器,分布在左、右侧突外侧面,在交配器上分布数量次于刺形感器,且较刺形感器稀疏。
2.4.3二叉状感器DIS (dichotomous sensilla)
二叉状感器(图4-F)端部分岔成二支,较长一支折向基部,仅在左侧突外侧面观察到2个,位于光滑面与密被感觉毛的交界处(图4-B)。

图4 黑肩绿盲蝽雄成虫交配器感器扫描电镜观察Fig.4 SEM observations of sensilla on the genital of male Cyrtorhinus lividipennis注:A,部分左侧突;B,部分左侧突;TS I,毛形感器;CAS,钟形感器;DS,凹状感器;CHS,刺形感器;SP,感觉孔;DIS,二叉状感器;SS I,栓锥形感器I;SS II,栓锥形感器 II。Note: A, Part of left genital styles; B, Part of left genital styles; TS I, Triochoid sensilla I; CAS, Campaniform sensilla; DS, Depression sensilla; CHS, Chaetica sensilla; SP, Sensory pits; DIS, Dichotomous sensilla; SS I, Styloconic sensilla I; SS II, Styloconic sensilla II.
2.4.4凹状感器DS (depression sensilla)
凹状感器(图4-A)与雌虫产卵器上浅凹状感器形态相似,表面无孔,位于左侧突U形两端。
2.4.5钟形感器CAS (campaniform sensilla)
钟形感器(图4-C)为表皮圆形凹陷,中央圆顶突起,表面无孔,感器数量很少,仅在左侧突U形端发现。
2.4.6栓锥形感器SS (styloconic sensilla)
栓锥形感器I(styloconic sensilla I, SS I; 图4-D)位于左侧突U形两端外侧,圆形深坑中央锥形突起,突起高0.8 μm,无纹,2个;栓锥形感器II(styloconic sensilla II, SS II; 图4-G)位于左侧突U形两端内外侧,形态与栓锥形感器I相似,表面光滑无孔,突起更高,2~3个。
2.4.7感觉孔SP (sensory pits)
感觉孔(图4-E)直径不到1 μm,表皮深陷成孔洞,孔周围具突起。感器数量较少,见于左、右侧突密被感觉毛区域。
3 结论与讨论
3.1 黑肩绿盲蝽外生殖器结构与生殖行为
黑肩绿盲蝽外生殖器为雌虫交配孔、产卵器和雄虫交配器。雌虫产卵器由第1、第2、第3产卵瓣构成,第1、2产卵瓣末端具锯齿,与其产卵与水稻组织的习性相适应,产卵时第2、3产卵瓣组成的产卵鞘通过肌肉活动,当产卵器扩大,产卵鞘打开,直到产卵器竖立,产卵鞘停在后方固定产卵器(Davis, 1955);雄成虫交配器由阳基、阳茎和侧突构成不对称结构,侧突的不对称伴有腹部后端的不对称,属于昆虫生殖器不对称现象(Huber, 2010),该结构有利于其灵活调整交配位置,交配时,雄虫与雌虫反向,腹部左右贴近(Liquidoetal., 1985),阳茎插入雌性生殖孔开口于第8、9腹节之间节间膜上的生殖孔,左右侧突抱握起固定作用(章士美, 1985)。
3.2 黑肩绿盲蝽外生殖器感器与功能
毛形感器是黑肩绿盲蝽与寄主或配偶接触时首先接触到的感器。昆虫触角感受器显微结构观察和电生理实验表明,毛形感器可感知机械压力和机械振动(余海忠, 2007),还可能感受空气流速功能、重力、声波等(Schenider, 1964);果蝇Drosophilamelanogaster雄成虫单侧抱器上毛形感器消除,该果蝇交配位置不再对称(Acebesetal., 2003),表明生殖器上毛形感器的机械功能;而昆虫产卵器鞘上的毛形感器在寄主选择或在产卵检测中可能具有定位和刺探位点的作用(Le Ralecetal., 1990)。可推测毛形感器同时具备机械感知和化学感知两种功能。
两种栓锥感器于雄虫左侧突上分布,且在U形前端较光滑处,无孔。Dweck等(Dwecketal., 2008)在Habrobraconhebetor(Say)第2产卵瓣上,以及第1、2产卵瓣连接处分别观察到这类感器,认为是机械感受器,以探测第1、2产卵瓣之间的运动,在霍氏啮小蜂阳茎外侧面(郑溢华等, 2016),鞘翅目步甲科阳茎上都发现此种感器(Kimetal., 1999; Kimetal., 2004),故推测栓锥感器会在阳茎插入交配孔时起机械功能。

锥形感器内有一个或几个神经元,表皮感觉迟钝,是机械或化学感受器(Shields, 2008),而寄生蜂产卵器的侧瓣端部上锥形感受器,常是化学感受器(Gutierrez, 1970),本文锥形感器I位于第3产卵瓣与腹部嵌合处,锥形感器II位于交配孔内,推测仅行使机械功能。
凹形感器在ArgepullataZadd(Zhangetal., 2014)产卵瓣上观察到,被认为是感知压力,从而在产卵器刺入寄主过程中对继续前进还是中止做出决定,而在盲蝽雌虫产卵瓣和雄虫左侧突上均有分布,可能在雌虫产卵和雌雄雄虫交配过程中起到机械感知作用。
钟形感器在白蛾周氏啮小蜂Chouioiacunea产卵器上有分布,表面无孔,可能对温湿度敏感(徐晓蕊等, 2012),则在黑肩绿盲蝽雄虫左侧突上,该感器有助于交配期间感知环境。二叉状感器少见报道,仅有Damaster(Coptolabrus)smaragdinusbranickii和D.blaptoides阳茎底部观察到形态近似感器(Kimetal., 1999),其功能有待进一步研究。
外生殖器感器还在两性求偶前期性选择中起到感受异性状态的作用。对豆娘生殖器感器研究表明,其雄性生殖器上存在与化学感器相似的感器,这类感器可以探测雌性豆娘的状态(Uhíaetal., 2005);观察鞘翅目多食亚目9个种交配过程,雄虫在雌虫周围通过中茎顶端感知雌虫生殖器是否打开,推测中茎顶端感器可获取雌虫位置和雌虫生殖器开放形状的信息(Düngelhoefetal., 2010)。黑肩绿盲蝽在求偶期,雄虫在追逐雌虫的过程中弯起腹部触碰雌虫腹部,尝试交配,而雌虫会选择是否接受交配。综上,黑肩绿盲蝽生殖器上感器是其与环境和自身生物学特性相适应的结果,利于其感知并选择寄主上合适的产卵位点,感知异性的交配状态信息等。本研究仅进行了扫描电镜观察,感器的功能有待进一步研究以验证。
猜你喜欢
汽车实用技术(2022年14期)2022-07-30
广西林业科学(2022年1期)2022-03-03
口腔医学(2021年10期)2021-12-02
环境昆虫学报(2021年2期)2021-05-14
四川蚕业(2021年1期)2021-02-12
昆明医科大学学报(2021年1期)2021-02-07
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
江西农业学报(2019年10期)2019-10-29
江苏农业科学(2014年3期)2014-07-16
