
温度及种子含水量对剪枝栎实象和青冈象卵发育的影响
2021-06-28 08:31:58肖瑞迪张智英陈小兰
环境昆虫学报 2021年3期
肖瑞迪,张智英*,李 维,杨 柳,陈小兰,
赵倩倩1,周廷予1,苏文华1
(1. 云南大学生态学与环境学院,昆明 650500;2. 云南大学生命科学学院,昆明 650500)
植食性昆虫与植物的相互关系,不仅表现在成虫及幼虫取食阶段,还反映在成虫产卵及卵发育时期。产卵是昆虫维持种群,繁衍后代的关键环节,它关系到后代的发育和存活。大多数植食性昆虫选择在寄主的表面产卵(Hilker and Meiners, 2011),少数产卵于植物组织中(李永福等,2015)。在长期的相互作用共同进化中,植物对产卵于体表的昆虫演化出一系列防御特性,如形成刺、毛及次生代谢物质等,阻碍昆虫产卵(Shapiro,1981;吴文伟等,2002;常金华等,2004;唐宇翀等,2010);着卵部位形成肿瘤或产卵部位周围叶片组织坏死,造成卵脱落(Berdnikovetal., 1992;Dossetal., 2000)。而针对植物对产于组织内的卵产生的防御反应报道较少(崔丽婷,2017)。
象甲属鞘翅目Coleoptera象甲总科Curculionoidea,为植食性昆虫。大部分种类的象甲产卵时用喙在产卵部位钻孔,再利用喙将卵送入植物组织中,其被认为是成功进化的一项关键创新(Anderson,1995;Oberprieleretal., 2007)。将卵产于寄主组织内有助于初孵幼虫获取食物,躲避天敌侵害,减少降雨、风吹、日晒等气候因子的影响,从而提高卵的存活率(Hilker and Meiners, 2011)。在进行剪枝栎实象Mechorisursulu(鞘翅目:卷叶象甲科)雌虫产卵前后剪断产卵果枝的独特产卵行为研究时发现,如果由于阻止雌虫的剪枝行为,卵直接产在活的壳斗科Fagaceae寄主植物种子中,则因产卵损伤寄主植物种子的组织,诱导了寄主的防御反应。最明显的表现就是在卵室部位形成愈伤组织(图1),使大部分的卵被挤压致死。而与剪枝栎实象同寄主且同样危害种子的象甲科Curculionidae青冈象Curculiomegadens,雌虫产卵时没有剪枝行为,可直接将卵产于活的寄主种子中。为了弄清青冈象能在活的寄主植物中产卵的机制,了解它是如何避免寄主植物愈伤组织对卵的影响,有必要对这两个生态位相近的象甲进行生物生态学等特性的比较研究。前期研究发现,剪枝栎实象产卵于寄主植物内是诱导寄主产生防御的关键,而在植物与昆虫卵的相互作用中,卵历期的长短及寄主形成愈伤组织的动态决定着卵的存活情况,而卵发育历期及存活率通常受温度、湿度及寄主含水量的影响。因此,摸清两种象甲卵生长发育与温度等因子的关系,对进一步分析寄主愈伤组织对卵的致死效应及青冈象如何避免寄主植物愈伤组织对卵的影响均有重要的理论意义。此外,由于两种象甲均是危害许多种壳斗科植物种子的重要害虫,摸清生态因子对两种象甲各虫态的影响,均可为有效防治其危害提供科学依据。
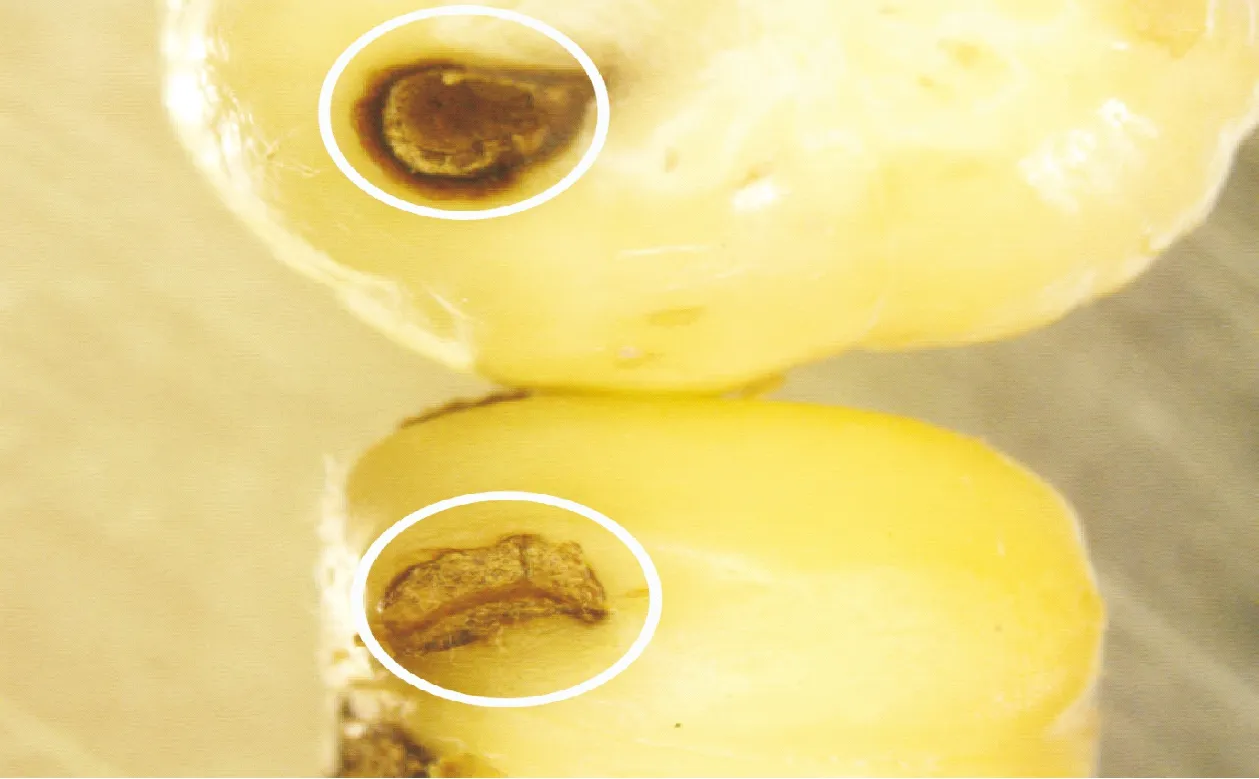
图1 剪枝栎实象雌虫剪枝后在锥连栎坚果上自然产卵(上)与人为阻止剪枝后产卵(下)5 d后卵室对比情况,显示被人为阻止剪枝后的坚果中卵室(下)被愈伤组织所填满Fig.1 The egg chamber (no callus, above)in a acorn of Quercus franchetii 5 days after female of Mechoris ursulus cut off fruit branch in natural oviposition was compared to one (filled callus, below)in same 5 days after the female was artificially prevented from cutting off fruit branch in oviposition
1 材料与方法
1.1 试验地概况
昆明植物园地处东经102°41′,北纬25°1′,海拔1 980 m,属于中亚热带内陆高原气候,干湿季节分明,冬春季低温干燥,晴朗少雨,日照充足,春季多风;夏季高温多湿,雨量集中。年平均温度14.7℃,年平均降雨量为1 006.5 mm,平均相对湿度为73%。土壤为典型的酸性红壤,pH 5.59,有机质含量低(龚洵等,2003)。昆明植物园现有植物近4 000种,设有10个专类植物区,其中的壳斗科植物区,种植有锥连栎Quercusfranchetii、麻栎Q.acutissma、槲栎Q.aliena、柞栎Q.mongolica和滇青冈Cyclobalanopsisglaucoides等众多壳斗科植物。
1.2 供试虫源
由于野外无法判断青冈象是否产卵,本试验采取室内饲养的方法获取青冈象当天所产的卵。剪枝栎实象由于产卵前期长达2-3月,室内饲养获取大量当天产的卵较难,而野外由于成虫产卵前后具有剪断产卵果枝的行为,便于识别。因此,实验所用的卵均为野外采集。青冈象卵采集方法:在昆明植物园收集受青冈象为害较严重的坚果,分别放入盛有细泥土的容器中,适时浇水以保持泥土湿度,用尼龙网罩住容器,待翌年成虫羽化出土后,将成虫放入饲养笼内,以10%蜂蜜水和清水饲养,待雌虫开始产卵后,每天放入新鲜的种子供其产卵,当天解剖种子,取出卵供实验用。剪枝栎实象卵采集方法:从8月下旬剪枝栎实象雌虫开始一定规模产卵后,在昆虫植物园锥连栎种植区,每天收集剪枝栎实象当天产卵掉落地上的果枝,解剖种子,取出卵供实验用。
1.3 试验方法
1.3.1温度对剪枝栎实象和青冈象卵发育的影响
试验温度分别设置为15、20、25、30和35℃,无光照。将青冈象和剪枝栎实象当日产的卵分别置于装有滤纸直径为9 cm的培养皿中,加灭菌水使滤纸相对含水量保持在50%~60%,每个培养皿放30粒卵,分别置于不同温度的培养箱内,各处理3个重复。每天9 ∶00和16 ∶00观察2次,并添加适量的水保持滤纸湿度。同时记录卵孵化日期和存活情况,统计不同温度下两种象甲卵的发育历期、发育起点温度、有效积温及卵的存活率。
1.3.2含水量对剪枝栎实象和青冈象卵发育的影响
剪枝栎实象和青冈象的主要寄主植物是壳斗科的柞栎和锥连栎。两种植物坚果的含水量明显不同,前期实验高温烘干测两种坚果的含水量,110.36 g新鲜柞栎测得绝对含水量为49.02 g,130.13 g新鲜锥连栎测得绝对含水量为21.23 g,其寄主的含水量是否会影响卵的发育,试验通过设置不同含水量的滤纸,模拟不同的种子含水量,研究对两种象甲卵发育的影响。试验含水量分别设置为20%、40%、60%和80%,温度为25℃,无光照。在直径为9 cm的培养皿中分别置入不同含水量的滤纸,然后将当日产的青冈象和剪枝栎实象卵放于滤纸上,每个培养皿放30粒卵,置于温度为25℃的培养箱内,各处理3个重复。每天观察记录卵孵化及存活情况。统计不同含水量下两种象甲卵的发育历期、发育起点温度、有效积温及卵的存活率。
1.3.3剪枝栎实象和青冈象卵和卵室体积的测量
实验样地收集大量受两种象甲危害严重的坚果,在解剖镜下解剖受虫害的坚果,然后取出卵和卵室,并在电镜下测量卵和卵室的体积,各处理30个重复。按照公式计算:体积V1=4/3*πr3(r为半径)或体积V1=4/3*πabh(a为长,b为宽,h为高)。统计和比较两种象甲卵和卵室的差异,并从两种象甲卵和卵室的形态特征分析其适应和进化的意义。
1.4 数据处理
加权平均法求出各剪枝栎实象和青冈象卵的发育历期(N=孵化日期-产卵日期),用发育历期的倒数表示两虫卵的发育速率V,根据有效积温法则,采用回归直线法计算虫卵的发育起点温度和有效积温,其公式如下:
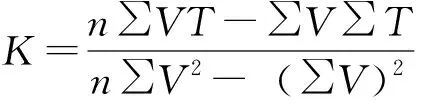
式中,n为试验中温度处理的组数;V为发育速率,N为象甲完成生长发育期所需的时间(d);T为该期的平均温度;T'为理论日均温度,可由T′=C+KV计算而得;K为有效积温,Sk为有效积温的标准误;C为发育起点温度,Sc为发育起点温度的标准误。
采用Excel进行实验数据的整理,然后用SPSS 13.0软件进行分析和显著性检验。采用单因素方差分析不同温度和含水量对卵的发育历期的显著差异性,运用Duncan氏新复极差法进行多重比较分析。
2 结果与分析
2.1 不同温度和种子含水量下剪枝栎实象和青冈象卵的发育历期和发育速率
温度对剪枝栎实象和青冈象卵的发育历期有显著影响(表1)。在15~30℃温度范围内,两象甲卵发育速率随温度的升高而明显加快,发育历期缩短。但在35℃时,剪枝栎实象卵发育历期延长,发育速率减慢,而青冈象卵发育历期和发育速率均为0,其卵无法正常发育。当温度低于30℃,剪枝栎实象和青冈象卵的发育速率随温度降低而延长发育历期,当温度高于30℃,剪枝栎实象和青冈象卵的发育速率随温度升高而延长发育历期,甚至卵死亡。30℃为其最适发育温度,两种象甲的卵发育速率最快,发育历期最短。说明温度过高或过低,对两种象甲的卵发育有抑制作用;当温度适宜时,对两种象甲的卵发育有促进作用。两种象甲在相同温度下,发育速率和发育历期不同。除了20℃外,相同温度下剪枝栎实象的卵发育速率大于青冈象的卵发育速率,剪枝栎实象卵的发育历期短于青冈象卵的发育历期。

表1 不同温度下剪枝栎实象和青冈象卵的发育历期和发育速率
根据表1中不同温度下剪枝栎实象和青冈象卵的发育速率,可求出发育速率V和温度T之间的线性回归方程:剪枝栎实象的卵V=0.008T-0.052,r=0.957;青冈象的卵V=0.0056T-0.06,r=0.967。在适宜的温度条件下,剪枝栎实象和青冈象的发育速率和温度之间呈显著正相关(表2)。

表2 剪枝栎实象和青冈象卵的发育速率与温度的回归方程
不同含水量处理对剪枝栎实象和青冈象卵的发育历期无显著差异(表3)。在含水量为20%~40%内,剪枝栎实象卵的发育历期随含水量的增大而缩短,发育速率增大;在含水量为40%~80%内,剪枝栎实象卵的发育历期随含水量的增大而延长,发育速率减小。当含水量为40%时,剪枝栎实象卵的发育历期最短,发育速率最快,故40%为其卵最适发育的种子含水量。

表3 模拟不同含水量下剪枝栎实象和青冈象卵的发育历期和发育速率
在含水量为40%~60%内,青冈象卵的发育历期随含水量增大而延长,发育速率减慢。在含水量为60%~80%时,青冈象卵的发育历期随含水量增大而缩短,发育速率加快。在含水量为20%时,青冈象卵的发育历期最短,发育速率最快,故20%为其卵最适发育的种子含水量。在相同含水量时,剪枝栎实象卵的发育历期均长于青冈象卵的发育历期,其卵发育速率均小于青冈象卵的发育速率。实验结果显示,含水量过高或过低,剪枝栎实象卵的发育历期延长,发育速率减慢,而青冈象卵的发育随含水量的降低历期缩短,发育速率加快。
2.2 剪枝栎实象和青冈象卵的发育起点温度和有效积温
根据有效积温法则,可算出剪枝栎实象和青冈象卵的发育起点温度和有效积温(表4)。剪枝栎实象卵的发育起点温度是7.41±0.09℃,有效积温是119.33±20.25日·度。青冈象卵的发育起点温度是3.83±0.08℃,有效积温是154.45±26.93日·度。剪枝栎实象卵的发育起点温度大于青冈象卵的发育起点温度,青冈象卵的发育起点温度较低,说明其对温度的适应能力强。剪枝栎实象卵的有效积温小于青冈象卵的有效积温。根据发育起点温度和有效积温,可建立发育历期预测式N=K/(T-C)。
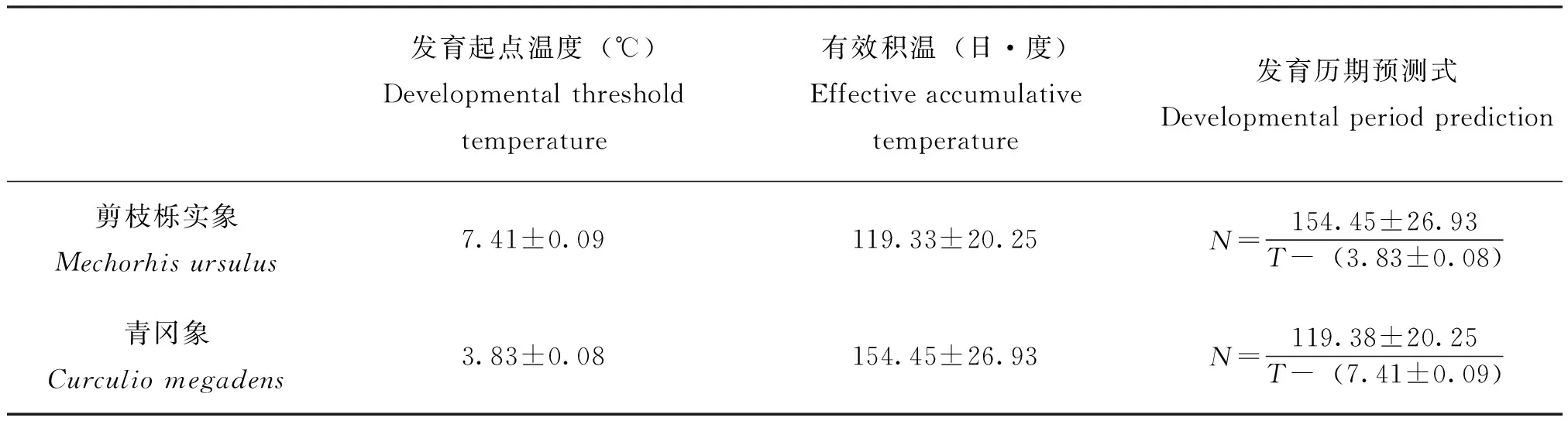
表4 剪枝栎实象和青冈象卵的发育起点温度和有效积温
2.3 不同温度和种子不同含水量下剪枝栎实象和青冈象卵的存活率
温度对剪枝栎实象和青冈象卵的存活率有显著影响(表5)。在15~25℃时,随着温度的上升,剪枝栎实象和青冈象卵的存活率逐渐增大;温度为25℃时,剪枝栎实象和青冈象卵的存活率最高。在30~35℃时,随着温度的上升,剪枝栎实象和青冈象卵的存活率逐渐减小。35℃时,青冈象的卵无法正常发育,其存活率为0,说明青冈象的卵在35℃下不能孵化。实验结果显示,温度过高或过低,对剪枝栎实象和青冈象卵的生长和发育有抑制作用,在相同温度下,剪枝栎实象卵的存活率高于青冈象卵的存活率。
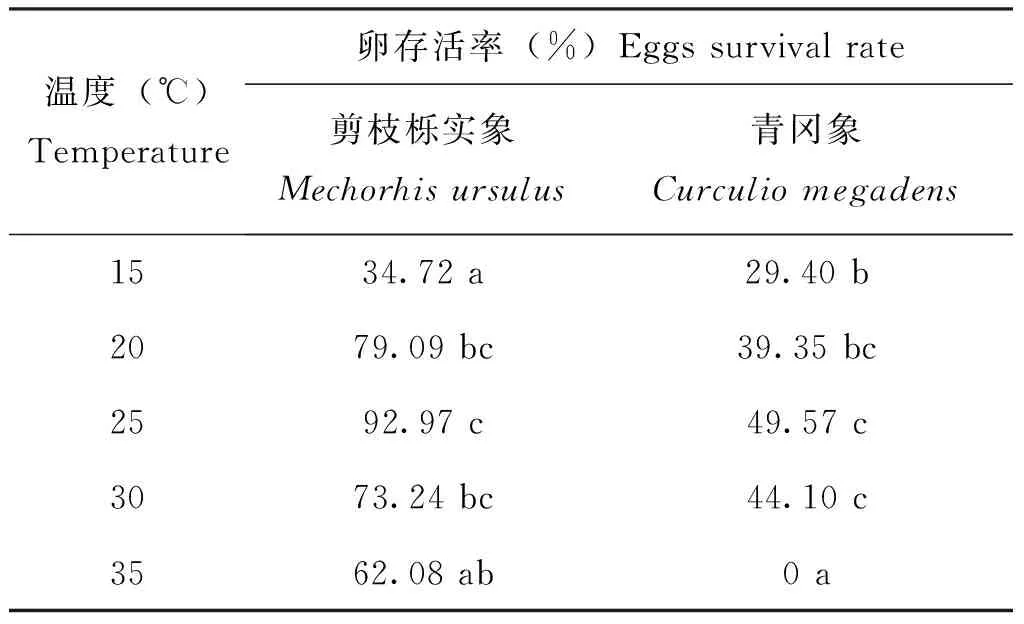
表5 不同温度下剪枝栎实象和青冈象卵的存活率
模拟种子不同含水量对剪枝栎实象和青冈象卵的存活率无显著影响(表6)。在含水量20%~80%,剪枝栎实象卵的存活率分别为63.33%,62.67%,71.59%,90.93%。结果显示除含水量40%外,剪枝栎实象卵的存活率随含水量增大而增大。在含水量为80%时,剪枝栎实象卵的存活率高达90.93%。说明寄主含水量较高,有利于剪枝栎实象卵的存活。在含水量20%~80%,青冈象卵的存活率分别为22.18%,22.32%,16.37%,29.13%。结果显示在含水量为20%~40%时,青冈象卵的存活率相近。在含水量为60%~80%时,随含水量增大,青冈象卵的存活率逐渐增大。说明寄主含水量较高,有助于提高青冈象卵的存活率。在相同含水量下,剪枝栎实象卵的存活率均大于青冈象卵的存活率。在含水量为80%时,剪枝栎实象和青冈象卵的存活率达到最大。说明寄主含水量越高,越有利于提高剪枝栎实象和青冈象卵的存活率。
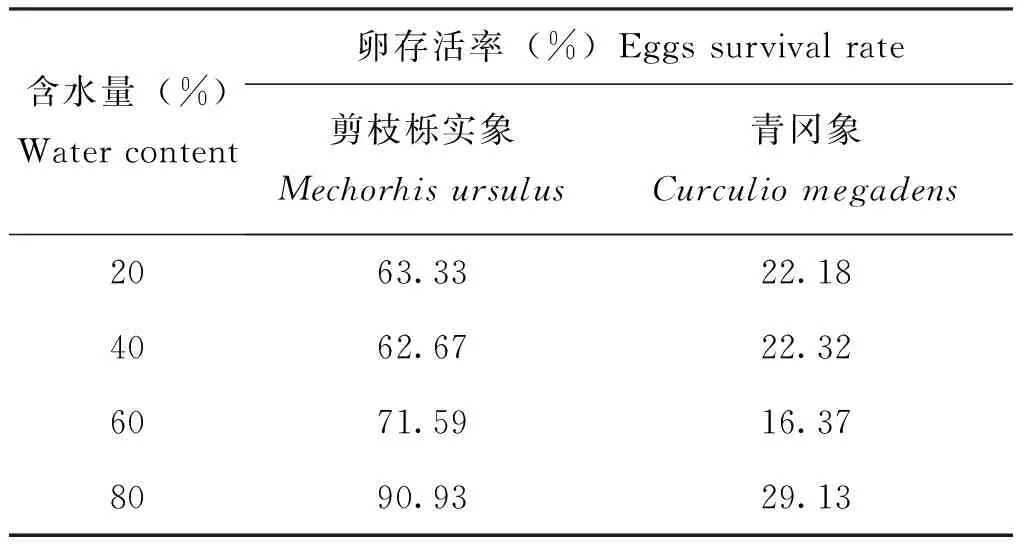
表6 模拟不同含水量下剪枝栎实象和青冈象卵的存活率
2.4 剪枝栎实象和青冈象卵和卵室体积
在电镜下测得剪枝栎实象和青冈象的卵和卵室大小有显著差异(表7)。30粒剪枝栎实象卵的平均体积为0.2368 mm3,范围为0.0704~0.9502 mm3;30粒青冈象卵的平均体积为0.0836 mm3,范围为0.0454 ~0.2439 mm3。30粒剪枝栎实象卵室的平均体积为3.9589 mm3,范围为0.6770 ~9.4118 mm3;30粒青冈象卵室的平均体积为0.4650 mm3,范围为0.0633~1.6268 mm3。结果说明青冈象卵和卵室明显小于剪枝栎实象卵和卵室。

表7 剪枝栎实象和青冈象卵和卵室的测量
3 结论与讨论
温度是昆虫生长发育的重要影响因素,对其研究的应用主要涉及对昆虫种群消长的评估,利用有效积温法则预测昆虫在某一地区的发生代数及其在地理上的分布界限等(沈建平等,2003;雷仲仁等,2007;何莉梅等,2019)。本文通过研究温度等因子对剪枝栎实象及青冈象卵生长发育的影响,为进一步探讨植物与产卵在其组织内的昆虫相互作用及昆虫的适应进化奠定基础。研究结果显示剪枝栎实象和青冈象卵的发育和存活均受温度的显著影响。两种象甲卵的发育速率与温度呈显著正相关。在15~30℃的温度范围内,卵的发育历期随温度的升高而缩短,发育速率加快。30℃时卵的发育历期最短。35℃时剪枝栎实象发育速率减慢,历期延长,而青冈象的卵则不能完成生长发育。在相同温度下,青冈象卵的发育历期比剪枝栎实象长。由此可见,青冈象并不是通过缩短卵期来避免寄主愈伤组织对卵的防御。
人为阻止剪枝栎实象剪枝行为后,寄主锥连栎形成愈伤组织充满卵室并致卵死亡,该过程通常需要5 d,而剪枝栎实象在昆明的产卵期为8-10月,期间室内变温情况下,剪枝栎实象卵的发育历期约7 d,与本研究恒温25℃处理较为一致。因此,寄主形成的愈伤组织明显快于卵的发育,这是愈伤组织能对卵形成较高致死率的原因之一。野外人为阻止剪枝栎实象雌虫剪枝行为后,产在活的寄主种子中的卵,死亡率为82.1%(n=28),而孵化出幼虫的卵通常是卵室中种子形成的愈伤组织较少或没有,原因可能是种子损伤后个体间形成愈伤组织的能力存在差异。
本实验在室内恒温条件下,剪枝栎实象卵的发育起点温度为7.41±0.09℃,卵的有效积温为119.33±20.25日·度;青冈象卵的发育起点温度为3.83±0.08℃,卵的有效积温为154.45±26.93日·度。室内变温条件下,欧洲栗象C.elephas卵的发育起点温度为6.5℃,卵的有效积温为108.9日·度(Manel and Debouzie,1995);豆象科的豌豆象Bruchuspisorum卵的发育起点温度为11.76±0.71℃,卵的有效积温为74.0±6.89日·度(谢成君,2003);蚕豆象B.rufimanus卵的发育起点温度为11.45℃,卵的有效积温为79.68日·度(郑兴国等,1991)。对比这两类亲缘关系较近的昆虫,象甲总科的这3种昆虫卵的发育起点温度比叶甲总科的这两种昆虫要低,而卵发育所需的有效积温却更多。而从温度对两种象甲卵的发育影响可以看出,剪枝栎实象卵发育适宜的温度范围比青冈象广,特别对高温的适应能力更强,这从剪枝栎实象分布区比青冈象广得以佐证。青冈象是滇中地区特有种,而夏季的云南中部地区气温不高,与剪枝栎实象相比,青冈象更适应夏季温度相对较低的地区。因此,愈伤组织致卵死亡的情况是否存在地区间差异,或者愈伤组织形成快的地区,卵的历期也较短,这些需进一步的研究才能摸清寄主愈伤组织与卵的相互关系。
从模拟种子含水量对剪枝栎实象和青冈象卵的发育看,种子不同的含水量对两象甲的卵历期无显著影响。但不同含水量的种子对愈伤组织的形成是否有影响有待进一步研究。如果愈伤组织受含水量影响,那么在卵历期不变的情况下,愈伤组织生长越快,可能对卵的致死能力就越强。
青冈象在含水量实验中卵的存活率比温度实验的结果低,发育历期也缩短。这可能是因为青冈象的卵是利用室内饲养的种群进行,而该实验是在雌虫产卵后期进行,雌虫后期所产的卵可能生活力降低。而剪枝栎实象是野外种群,实验所用的卵来自不同的种群,卵的活力未受到影响。
在相同的温度和含水量下,青冈象卵的存活率明显低于剪枝栎实象,这可能与两者卵及卵室的大小有关。青冈象开始产卵的时期比剪枝栎实象晚,产卵期寄主果实更大,然而电镜测量结果显示青冈象的卵和卵室明显小于剪枝栎实象,这可能是青冈象的一种适应进化,而青冈象卵的低存活率可能是其进化不完善性的表现。此外,本次实验中将青冈象的卵从卵室中取出培养,卵周围没有了寄主组织的包围,也可能是导致青冈象卵存活率降低的原因之一。当青冈象的卵连同卵室周围的组织一起置于培养皿中观察时,又因实验材料发霉而没有得到最终结果。故青冈象卵存活率低的原因还有待进一步探究。
除象甲外,鞘翅目天牛科、双翅目实蝇科等昆虫也将卵产于植物组织中,这些昆虫各自有一些自身的产卵特性(李焕存,2000;胡陇生等,2012;李永福等,2015),但它们是如何避免产卵引起的寄主防御反应也待进一步研究。
致谢:本研究野外调查取样得到中国科学院昆明植物园的大力支持,特此致谢!
猜你喜欢
莫愁(2023年9期)2023-03-17 09:37:12
莫愁·家教与成才(2023年3期)2023-03-15 00:55:23
中国农业科学(2022年15期)2022-08-09 02:23:48
中国马铃薯(2022年2期)2022-07-05 00:12:08
湖北农业科学(2021年22期)2021-12-13 13:00:36
四川农业科技(2019年10期)2020-01-06 03:02:02
黑龙江粮食(2019年10期)2019-11-23 02:09:32
中国稻米(2017年2期)2017-04-28 08:00:06
西藏科技(2016年9期)2016-09-26 12:21:38
江苏农业科学(2016年6期)2016-07-25 00:16:35
