
小兴安岭不同区域天然次生林下红松种源试验1)
2021-06-26 01:51:02魏志刚
东北林业大学学报 2021年6期
魏志刚
(国家林业与草原局盐碱地研究中心(中国林业科学研究院),北京,100091)
王瑞琪 刘莹莹 夏德安 杨传平
(林木遗传育种国家重点实验室(东北林业大学))
李若林
(丽林实验林场)
商永亮
(带岭林业实验局)
舒华铎
(双丰林业局)
阔叶红松林以红松(Pinuskoraiensis)为主,伴生树种主要有紫椴(Tiliaamurensis)、水曲柳(Fraxinusmandschurica)和胡桃楸(Juglansmandshurica)等珍贵阔叶树种,是小兴安岭地区稳定性最好的生态系统和最重要的木材生产基地[1]。建国58 a内,小兴安岭共生产木材3.28亿m3,每年采伐量为生长量的2.0~2.5倍,加之森林保护措施不力、毁林开荒、森林火灾和森林病虫害等原因[2],红松资源消耗殆尽、绝大部分阔叶红松林已退化为破坏程度不同天然次生林[3]。2004年伊春林区停止采伐时,红松林面积和蓄积比例分别由20世纪50年代的53.8%和74.3%下降到4.51%和4.5%,红松的蓄积量仅存400多万m3[4]。由阔叶红松林退化而成的天然次生林遗传多样性下降、整体生态功能和生产力低下,已无法满足该区域社会经济可持续发展和生态环境安全的基本需求[5]。因此,如何提高现有天然次生林的生产力和生态保护功能,已成为区域内森林经营的首要问题。研究表明,天然次生林是阔叶红松林中建群树种红松遭到毁灭性破坏演化形成,红松在群落中的数量比例与生长情况是次生林恢复成败的关键,因此通过天然次生林下间隙和斑块栽植红松等“栽针保阔”措施已成为该区域天然次生森林经营的主要任务[3,6-7]。
小兴安岭纵贯黑龙江省中北部,全长约450 km、宽度约210 km,区域内北部多台地、宽谷,中部低山丘陵、山势和缓,南部属低山、山势较陡,区域内不同地点的次生林下造林环境(土壤、光照、年积温和植被类型)差异巨大,对红松林下造林效果产生巨大影响。长期以来,小兴安岭地区天然次生林下造林用的红松苗木均为皆伐地选育出的良种或品种,没有经过不同地点次生林下遗传稳定性测定,造林后不同地点的红松幼林分化严重,从而对该区域天然次生林向阔叶红松林的恢复进程产生严重影响。针对上述问题,本项研究在前期红松种源试验研究结果的基础上[8-9],通过分析红松各种源在小兴安岭不同地点天然次生林下造林后的生长与适应性状变异,阐明地点对次生林下不同红松种源造林后生长与适应性状变异的影响及规律,从而为不同区域天然次生林林下造林选择出优良红松种源。本项研究将为小兴安岭不同地区天然次生林下红松造林用良种选育提供理论依据和造林良种。
1 材料与方法
1.1 种源材料
根据课题组前期“6.5”转“7.5”科技攻关课题“红松种源试验”结果[8-9],选择红松小兴安岭种源区表现较好的10个种源:黑龙江省带岭、金山屯、铁力、黑河、鹤岗、汤原,吉林省的敦化、白河、汪清与辽宁省本溪种源。
1.2 造林地点与林型
按区域位置与前期工作基础,在小兴安岭天然次生林分布区北部、中部和南部分别选择丽林实验林场(简称丽林)、带岭林业实验局(简称带岭)和双丰林业局(简称双丰)3个试点,基本代表了该区域典型气候和地理特征。上述3个试点的主要次生林类型和具体情况见表1。

表1 不同试点天然次生林的基本信息
1.3 各试验点造林方案
各试验地点均采用随机完全区组设计,10次重复,根据试验地面积大小确定小区树为10~20株,株行距为2 m×2 m,单行或者双行小区。区组间用1行红皮云杉做隔离行,试验地四周用1行红皮云杉做保护行,保护行和隔离行的株行距与红松相同。各种源苗木为均为4年生,2018年春季造林。
1.4 性状调查与统计分析
2020年10月,林木停止生长后,对各试点试验林10个区组内的所有红松植株进行地径、树高、当年高、3年高生长和保存率调查。
在统计分析前,依据拉依达准则去除异常数据[10],并进行以下参数估算:
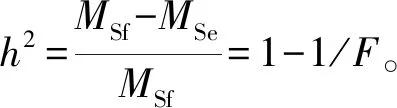

数据计算采用Minitab和SAS软件进行性状基本统计、方差分析和多重比较。
2 结果与分析
2.1 不同地点红松种源生长与适应性状
不同地点各林型下红松造林3 a后生长与适应性状统计分析结果(表2)表明,丽林试点红松各种源地径、树高、当年高、3年高和存活率的性状与变异系数均值分别为0.71 cm和20.52%、0.34 m和25.37%、0.07 m和31.53%、0.18 m和22.86%、28.91%和93.44%;带岭试点各种源上述性状分别为0.73 cm和17.38%、0.3 m和25.17%、0.07 cm和65.85%、0.17 cm和45.48%、41.00%和81.30%;双丰试点各种源上述性状分别为0.76 cm和16.93%、0.39 m和20.56%、0.08 cm和32.03%、0.20 cm和32.79%、57.26%和52.48%。3个试点红松各种源生长与适应性状变异系数的均值分别为38.74%、47.04%和30.96%,表明带岭试点次生林林下环境最复杂,对红松种源各性状的影响最大,其次为丽林试点,最后为双丰。上述结果初步表明,造林地点的不同会对红松幼林的生长与适应性状产生影响。
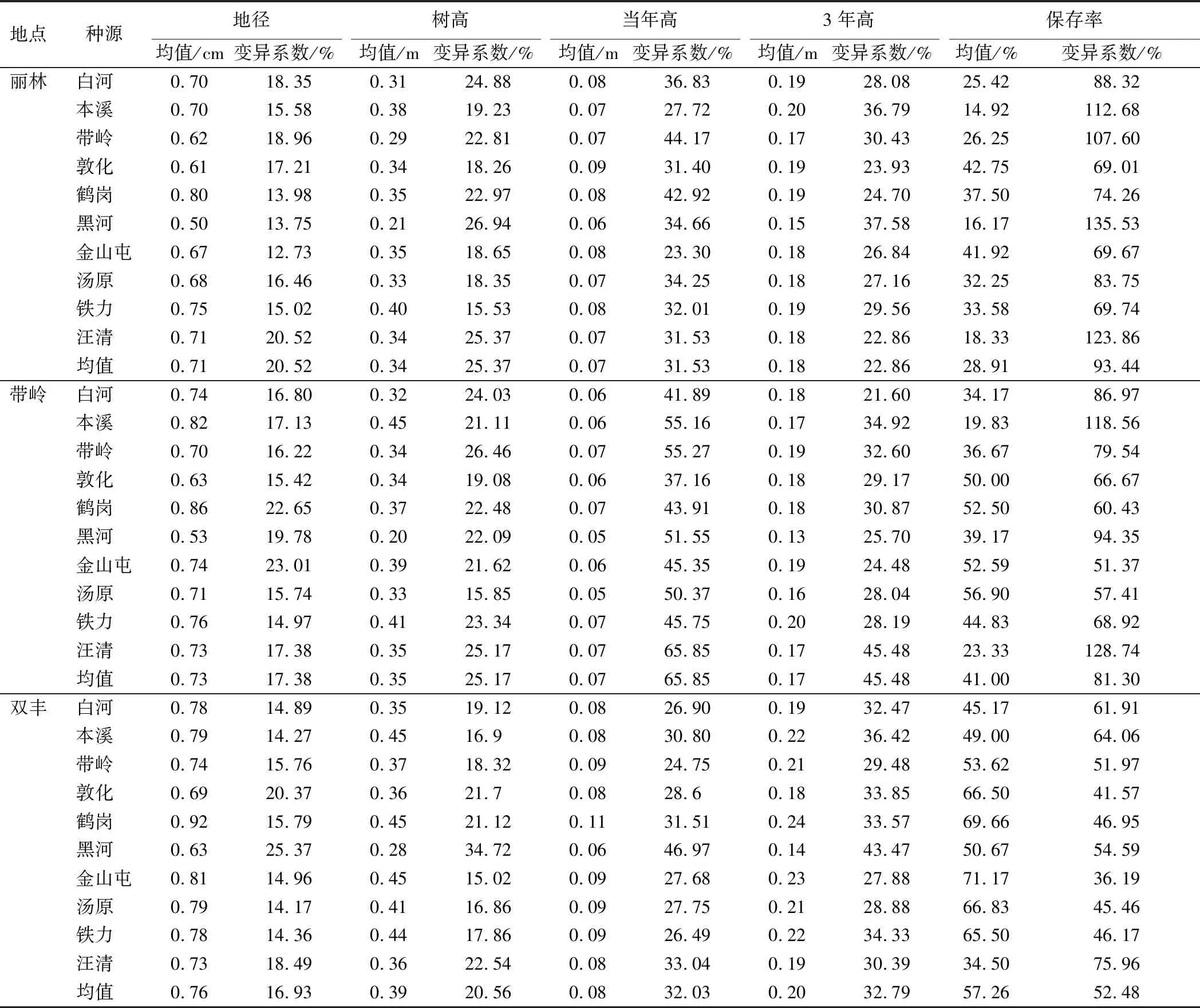
表2 不同地点天然次生林下造林3 a后红松各种源生长与适应性统计与分析结果
2.2 不同地点红松各种源幼林期生长差异性
方差分析发现:不同地点和不同种源间的生长与适应性状存在极显著差异。同时,地点与种源之间在地径、树高和当年高性状上交互作用极显著(表3)。上述结果表明,小兴安岭地区次生林下红松造林时,必须考虑区域位置与种源因素对造林效果的影响。

表3 不同地点和不同种源间生长与适应性状的方差分析结果
2.3 适宜造林地点与各地点优良种源选择
不同地点间红松各性状多重比较发现:双丰试点红松各种源各性状表现最好,是最优的造林地点。其次为带岭试点,除当年高外和3年高外,其余性状均显著优于丽林试点。丽林试点红松各种源生长与适应性状表现最差(表4)。上述结果表明,有必要分不同区域选择红松优良种源。

表4 不同地点红松种源生长与适应性状的多重比较结果
不同试点红松种源生长与适应性状方差分析发现:丽林试点,除3年高生长外,各种源间地径、树高和保存率均存在极显著差异,当年高性状种源间存在显著差异;带岭试点,除当年高外,各种源间其余性状均存在极显著差异;双丰试点,红松各种源间上述性状均存在极显著差异(表5)。
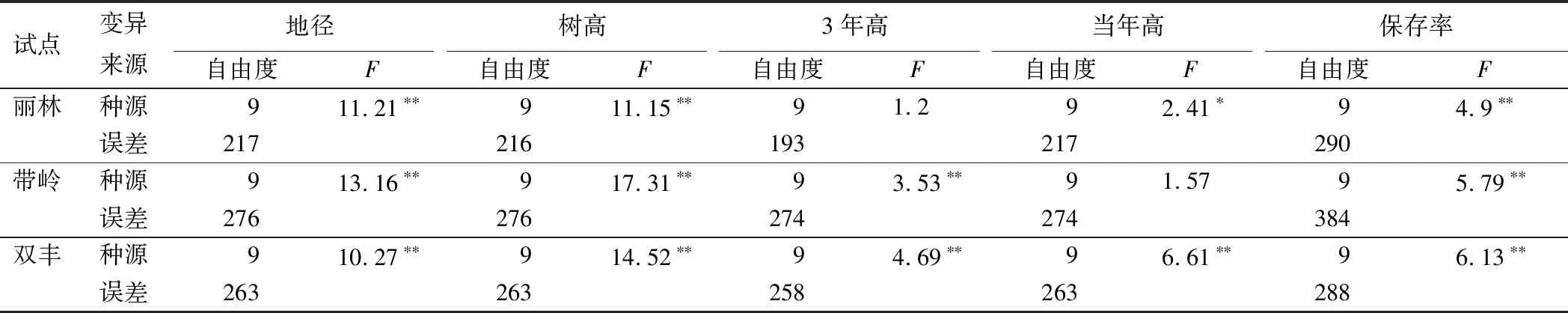
表5 不同试点红松生长与适应性状的方差分析结果
种源间差异显著性状遗传力计算分析发现,丽林试点种源地径、高、当年高性和保存率状遗传力分别为0.91、0.91、0.80和0.59,带岭试点种源地径、高、3年高和保存率遗传力分别为0.92、0.94、0.72和0.83,双丰试点种源各性状分别为0.90、0.93、0.79、0.85和0.84,各种源在不同地点的生长与适应性状属于强度遗传范围,性状变异受遗传因素影响大,通过选择能获得较好的遗传增益。此上,同一性状在不同地点的遗传力存在一定差异,也进一步证明遗传力是特定条件下的测定参数。
丽林试验点种源间存在显著和极显著差异的性状多重比较结果(表6)发现:在地径性状上,鹤岗种源表现最优,但与铁力、汪清和白河等种源之间无显著差异;树高性状上,铁力种源最优,但与本溪、金山屯、鹤岗和汪清种源间无显著差异;当年高性状上,敦化种源表现最优,但与鹤岗、铁力和金山屯等种源间无显著差异;保存率性状,敦化种源最优,但与金山屯、鹤岗和铁力等种源间无显著差异。综合各种源各性状多重比较结果,铁力种源各性状表现最好。该试点选择铁力种源造林,在地径、树高、当年高和保存率性状上可获得的遗传增益分别为10.55%、20.08%、3.04%和10.60%。
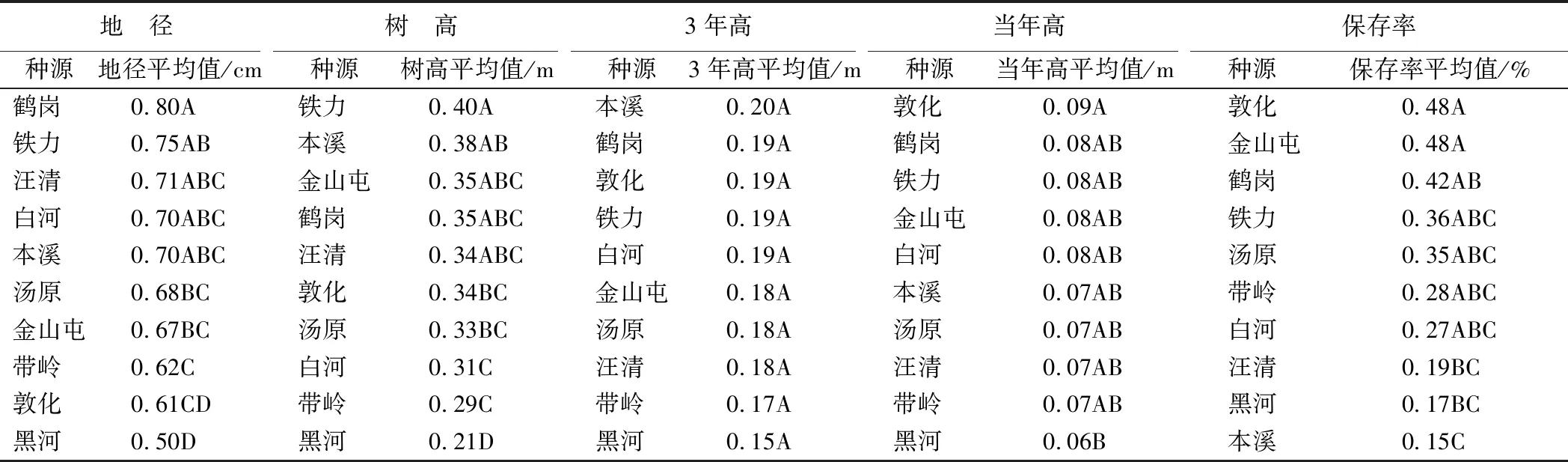
表6 丽林试点种源间的多重比较
带岭试验点种源间存在显著差异的各性状多重比较结果(表7)表明:铁力种源在地径、树高、当年高和保存率上表现最优,为该试点优良种源。该试点选择带岭种源造林,在上述性状上可获得12.06%、15.45%、11.57%和12.18%的遗传增益。

表7 带岭试点种源间的多重比较
双丰试验点种源间存在极显著差异的各性状多重比较结果(表8)表明:鹤岗种源所有性状综合表现最优,为该试点优良种源。选择该种源造林后,在上述性状上可获得遗传增益分别为18.61%、13.49%、15.40%、20.25%和18.19%。
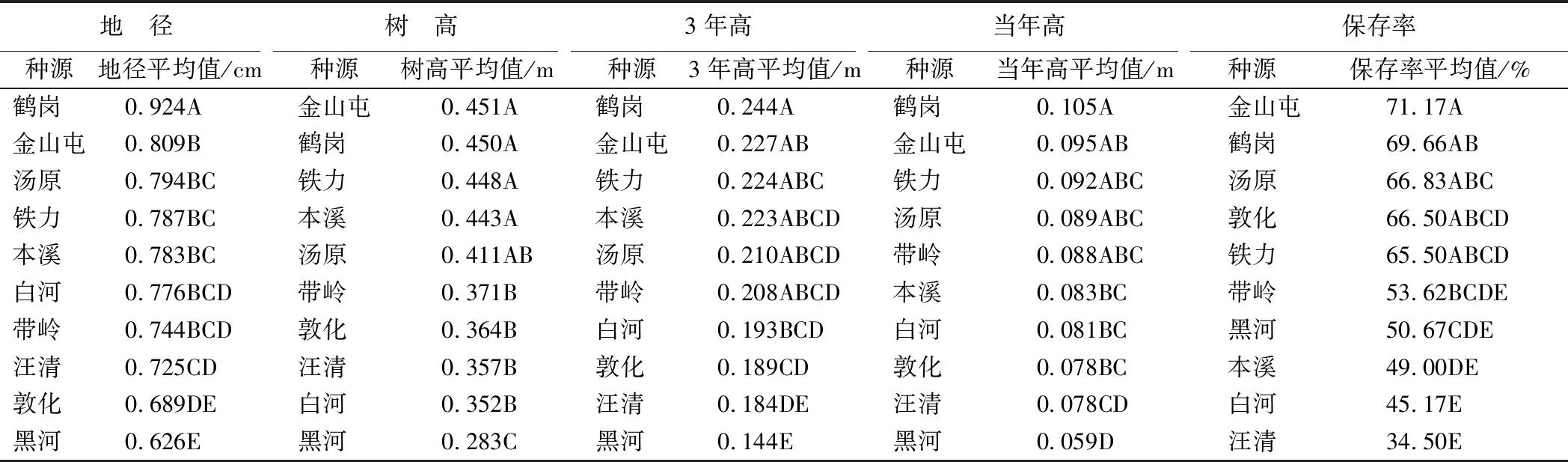
表8 双丰试点红松种种源性状多重比较结果
3 结论与讨论
小兴安岭纵贯黑龙江省中北部,区域内地形复杂,不同地点的次生林下造林环境(土壤、光照和植被类型)差异巨大[4,12]。因此,该区域次生林下进行红松造林时,应考虑地点因素对林下红松的生长与适应性的影响。红松分布范围较,在我国东北地区主要分布于长白山到小兴安岭一带、并呈新月型走向,并且分布高度随纬度增高而下降,分布区内不同群体间存在极为广泛的遗传变异[9,13-14],为不同地点天然次生林下红松优良种源选择提供了丰富的育种群体。
不同试点红松各性状的变异系数不同,表明红松生长与适应性状在不同地点会产生一定的分化。其中,带岭试点红松各性状变异系数均值最大,表明该地点次生林林下环境最复杂,对红松种源各性状的影响最大,而双丰试点次生林下环境相对变异较小,红松各性状的变异幅度较小。方差分析结果表明,地点对红松性状影响达到极显著水平。上述结果,进一步实证不同地点的同一林型,由于林隙大小、小气候特征和土壤的物理性状等环境因子不同会对红松生长产生显著影响[15]。不同地点红松生长与适应性状多重比较进一步发现,双丰试点红松种源各性状表现最好,是最优造林地点,其次为带岭试点,最后为丽林试点。上述结果完全符合红松分布区不同地点种群的生长规律。虽然小兴安岭地区为红松天然分布区,然而,分布区不同地点气候存在较大差异[9]。南部(双丰)由于水热条件要优于北部(丽林),因此红松生长速度快,北部则相反,而中部(带岭)其生长性状表现介于南部与北部性状之间。上述结果进一步证明,天然次生林下,红松造林时,必须做到“适地点适种源”。
不同地点具有显著和极显著差异的性状遗传力不同,且多为中强度遗传性状,如地径、树高和保存率的遗传力于均大于0.70,通过选择能获得较高的遗传增益。此外,同一性状在不同地点遗传力不同,如树高在杨桦林、硬阔林和杂木林0.91、0.94和0.93。在红松半同胞家系选择研究中,也发现红松地径和树高性状遗传力均超过0.5,但具体值与本项研究结果不同[16]。上述结果不仅表明红松种源不同性状在不同林型下遗传力不同,同时也进一步证明遗传力是特定条件下的估算参数。
由于红松地径、树高和当年高性状存在地点与种源间极显著交互影响,因此有必要为不同地点天然次生林选择适宜种源。而且,同一地点,红松种源间生长与适应性状存在显著与极显著差异,这与皆伐迹地上红松各种源生长性状存在极显著差异结果具有一致性[14]。其中,丽林试点铁力种源为优良种源,其造林后在地径、树高、当年高和保存率性状上的遗传增益分别为10.55%、20.08%、3.04%和10.60%;带岭试验点铁力种源表现最优,其造林后在地径、树高、当年高和保存率上可获得12.06%、15.45%、11.57%和12.18%的遗传增益;双丰试点鹤岗种源表现最好,造林后在地径、树高、3年高、当年高和保存率性状上的遗传增益分别为18.61%、13.49%、15.40%、20.25%和18.19%的。
本项研究结果将为小兴安岭不同地区天然次生林恢复顶级群落-阔叶红松林用红松优良种源选择提供理论依据和物质基础,对于提高该区域天然次生林的经营水平具有积极意义。
猜你喜欢
种子(2023年7期)2023-09-24 07:40:54
特产研究(2022年6期)2023-01-17 05:05:02
格言·校园版(2022年17期)2022-07-06 22:01:48
小哥白尼(野生动物)(2021年9期)2022-01-17 02:31:00
辽宁林业科技(2021年1期)2021-03-16 09:17:08
现代农业研究(2017年11期)2018-01-12 21:03:03
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
现代园艺(2017年20期)2017-02-03 18:49:29
现代营销(创富信息版)(2016年6期)2016-11-21 06:00:03
