
精氨酸处理对采后冬枣黑斑病的抗病作用
2021-06-19 06:02常璐璐张乐乐于有伟王小佳张少颖
食品工业科技 2021年12期
常璐璐,张乐乐,于有伟,王小佳,张少颖
(山西师范大学食品科学学院,山西临汾 041004)
冬枣果实营养丰富,含有人体所需的19 种氨基酸和钾、铁、铜等多种微量元素,还含有VC、VB1、VB2等多种维生素[1],口感脆甜,鲜食品质极佳,深受人们喜爱。冬枣在贮藏期间最易受互隔交链孢(Alternaria alternata)侵染而产生黑斑病,因此,采后冬枣的防腐保鲜一直是人们研究的热点[2]。目前,国内外研究的保鲜方法主要包括物理、化学和生物保鲜法[3]。物理保鲜方法如气调[4]和减压技术[5]能有效抑制果实的呼吸,减慢果实水分的蒸腾速度和抑制微生物活动,延缓果实的衰老进程,减少腐烂。化学处理方式如1-MCP[4]或CaCl2[6]处理也能有效保持冬枣的硬度,降低冬枣腐烂率,减缓冬枣中可滴定酸和VC含量的下降。常用生物制剂保鲜也能够有效抑制冬枣的腐烂,如壳聚糖[7]、金针菇蛋白酶解肽[8]、银杏叶提取液[9]等处理在诱导抗病方面也有一定效果。目前在产业中应用较多的冬枣防腐保鲜技术包括冷藏[10]、气调包装[4]。虽然,上述保鲜技术都具有一定的效果,但是考虑到经济、便捷等因素,寻求更为实用方便的诱导抗病防腐保鲜技术就显得更为迫切。
精氨酸(Arginine,Arg)是多胺和NO 合成的前体物质,二者在多种果蔬的生物胁迫和非生物胁迫中有着重要作用[11]。目前,Arg 多用于鹤望兰[12]、月季[13]、百合[14]等鲜花的保鲜处理;也有研究发现采前Arg 处理能够诱导番茄果实的抗病性[15],同时发现内源NO 浓度与处理后的苯丙氨酸解氨酶(phenylalamineammonialyase,PAL)、多酚氧化酶(polyphenol oxidase,PPO)、几丁质酶(chitinase,CHI)及β-1,3-葡聚糖酶(β-1,3-glucanase,GLU)活性呈正相关,说明Arg 通过影响NO 生物合成和调控防御酶的活性来诱导果实的抗病性。另外,Arg 处理对番茄灰霉病也有一定的抑制效果[15]。有研究还发现,Arg 处理抑制了白蘑菇贮藏期间PPO 和PAL 活性,诱导了超氧化物歧化酶(superoxide dismutase,SOD)和过氧化物酶(peroxidase,POD)活性,增强了白蘑菇的抗病能力[16−18]。但是关于Arg 处理诱导采后冬枣果实抗病性方面的研究还很少,而且也缺乏系统深入的研究。
本研究以冬枣果实为试验材料,通过不同浓度Arg 溶液浸泡处理,研究损伤接种A.alternata后,冬枣果实病斑直径、抗病相关酶活性(PAL、POD、GLU、CHI)以及抗病物质(黄酮、多酚、木质素)的变化,以期丰富Arg 处理在果实采后抗病保鲜方面的理论研究,并为冬枣的采后防腐保鲜提供新思路。
1 材料与方法
1.1 材料与设备
供试白熟期冬枣果实 于2020 年9 月27 日采自山西省临汾市尧都区尧乡冬枣种植基地,采摘后迅速用纸箱包装,运输至实验室,室温(26±2 ℃)贮藏备用;供试Arg 购自上海阿拉丁试剂公司;A.alternata(BNCC115062) 购自北纳生物有限公司,于马铃薯葡萄糖琼脂培养基(PDA)中保存,使用前于PDA 培养基上28 ℃培养10 d。
H1850R 高速台式冷冻离心机 湘南湘仪实验室仪器开发有限公司;SpectraMax M2/M2e 酶标仪 上海普迪生物科技有限公司;IMS-40 全自动雪花制冰机 常熟市雪科电器有限公司;759S 紫外可见分光光度计 上海棱光技术有限公司;MDF-U53V超低温冰箱 日本三洋集团;OLYMPUS-CX31 生物显微镜 山东博科再生医学有限公司。
1.2 实验方法
1.2.1 孢子悬浮液制备 取28 ℃培养10 d 的链格孢菌,加入无菌水(含0.05% Tween-20)后,刮下培养皿上的孢子,三层纱布过滤后,在显微镜下用血球计数板计数、稀释,配制成1×105个/mL 的孢子悬浮液。
1.2.2 果实浸泡、损伤接种处理 用2%次氯酸钠溶液浸泡枣果实2 min,自来水冲洗后自然晾干。将晾干后的果实分为四组,每组200 个,然后分别在0、20、200、1000 μmol/L Arg 溶液浸泡10 min,取出自然晾干,装于塑料盒中,于室温(26±2 ℃)、相对湿度90%下贮藏24 h。24 h 后用75%的酒精均匀喷洒消毒,之后用直径为3 mm 的打孔器在枣果实中部打一个直径为3 mm,深度为2 mm 的孔,取5 μL 配置好的A.alternata孢子悬浮液(1×105个/mL)接种于孔内。接种完后装入塑料盒中用聚乙烯薄膜封口于室温(26±2 ℃)、相对湿度90%下贮藏。定期观测病斑直径,每次观测60 个果实,重复三次。每隔1 d取10 个枣果实损伤部位皮下0.2~10 mm 范围内的果肉存放于超低温冰箱(−80 ℃)内用于测定后期指标。
1.2.3 测定方法
1.2.3.1 病斑直径测定 利用十字交叉法测量冬枣的病斑直径,每次测量60 个冬枣病斑的横纵径,重复3 次(每20 个为一组),求平均值。
1.2.3.2 PAL、POD、GLU 和CHI 活性的测定 PAL活性的测定参照曹建康等[19]的方法,做适当修改。取4 g 枣粉,加入4 mL 提取液(含有40 g/L 聚乙烯吡咯烷酮(PVPP)、2 mmol/L 乙二胺四乙酸(EDTA)和5 mmol/Lβ-巯基乙醇)。在4 ℃、12000×g 离心30 min,收集上清液。取两支试管,一支试管加入3 mL 硼酸缓冲液(50 mmol/L、pH8.8),0.5 mL L-苯丙氨酸溶液(20 mmol/L)、0.5 mL 酶液;另一支将酶液煮沸6 min后加入。两支试管均于37 ℃下保温10 min,加入0.1 mL HCl(6 mol/L)终止反应,在290 nm 处测定吸光度值。以吸光度值增加0.01 为1 个PAL 活性单位(U),表示为U·h−1·g−1FW。
POD 活性的测定参照曹建康等[19]的方法,稍作修改。离心管中加入3.0 g 果蔬组织样品和3.0 mL提取缓冲液,于4 ℃,12000×g 离心30 min,收集上清液,低温保存备用。取一支试管,反应体系包括3.0 mL 愈创木酚溶液(25 mmol/L)和0.5 mL 酶提取液,最后加入200 μL H2O2溶液(0.5 mol/L),同时开始计时。在波长470 nm 处测定吸光度值,15 s 后开始记录,每隔1 min 记录一次,重复三次。以吸光度值增加1 为1 个过氧化物酶活性单位(U),表示为U·min−1·g−1FW。
GLU 活性的测定参照曹建康[19]的方法稍作修改。取2.0 g 果蔬组织样品,加入2.0 mL 经预冷的提取液(含1 mmol/L EDTA-Na2、5 mmol/Lβ-巯基乙醇、1 g/L L-抗坏血酸),于4 ℃,12000×g 离心30 min,收集上清液。准备2 支试管,分别加入100 μL昆布多糖(4 g/L),1 支试管中加入100 μL 上清液、另一支加入100 μL 煮沸上清液。37 ℃保温40 min后加入1.8 mL 蒸馏水和1.5 mL 3,5-二硝基水杨酸(DNS),最后用蒸馏水稀释至25 mL。测定540 nm处的吸光度值,分别记作ODS和ODC。重复三次,以每秒钟每克果蔬组织分解昆布多糖产生 1×10−9mol 葡萄糖为一个 GLU 活性单位(U),表示为U·mol·s−1·g−1FW。
CHI 活性测定参照曹建康等[19]方法并作修改。取3 g 枣粉,加入4 mL 预冷丙酮,于4 ℃、12000×g离心30 min。反应体系为0.5 mL 乙酸-乙酸钠缓冲液(50 mmol/L、pH5.2)、0.5 mL 胶状几丁质悬浮液(10 g/L)和0.5 mL 酶液,对照管煮沸5 min。37 ℃水浴1 h,加0.1 mL 脱盐蜗牛酶(30 g/L)继续水浴1 h,最后加0.2 mL 四硼酸钾溶液(0.6 mol/L),煮沸3 min 后迅速冷却,于585 nm 处测定吸光度值。酶活性单位是U·mol·s−1·g−1FW。
1.2.3.3 黄酮、多酚、木质素含量的测定 黄酮测定参考武继芸[20]的方法,稍作修改。取0.5 g 果蔬组织样品,加入4.0 mL 80%的乙醇,于4 ℃,8000×g 离心20 min,收集上清液待用。取0.2 mL 样液,加入0.6 mL NaNO2试剂,摇匀反应5 min 后加0.6 mL Al2Cl3溶液(10%),混匀后于室温下放置6 min,最后加入2 mL NaOH 溶液(1 mol/L),室温下放置15 min。在510 nm 处测定吸光度值。用芦丁标准品制作标准曲线,所有实验均重复测定3 次。总黄酮含量以样品中所含芦丁的当量(mg·g−1FW)来表示。
多酚测定参考Fan 等[21]的方法,取0.5 g 果蔬组织样品,加入4.0 mL 80%的乙醇,于4 ℃,8000×g 离心20 min,收集上清液待用。取0.2 mL样液,加入2.0 mL Folin-Ciocalteau 试剂(稀释10 倍),反应5 min后加入2.0 mL Na2CO3(10%),混匀后于室温下放置1.5 h,在765 nm 处测定吸光度值。用没食子酸溶液制作标准曲线,所有实验均重复3 次。总酚含量以样品中所含没食子酸的当量(mg·g−1FW)来表示。
木质素测定参照常晨等[22]的方法,取1 g 果蔬组织样品,加入4.0 mL 预冷的95%的乙醇混匀,于4 ℃,12000×g 离心10 min,分别用95%的乙醇、乙醇:正己烷=1:2(v/v)冲洗沉淀三次。收集沉淀后加入1 mL 的溴乙酰(25%),70 ℃恒温水浴30 min。加入1 mL NaOH 溶液(2 mol/L)终止反应,再加入2 mL 冰醋酸和0.1 mL 盐酸羟胺(7.5 mol/L)。离心后取上清液0.25 mL,用冰醋酸定容至5 mL。在280 nm 处测定吸光度值,所有实验均重复测定3 次。木质素含量以OD280·g−1FW 表示。
1.3 数据处理
使用SPSS 处理数据并进行Duncan’s 多重比较分析,Origin 2019 绘制图表。
2 结果与分析
2.1 Arg 处理对接种A.alternata 后冬枣果实病斑直径的影响
冬枣采后接种A.alternata后病斑直径在贮藏期间呈上升趋势,由图1 可知,在贮藏的20 d 中,200 μmol/L Arg 处理组的病斑直径最小,对照组的病斑直径最大。在贮藏的第8 d,对照组的病斑直径分别比20、200、1000 μmol/L Arg 处理高57.40%、60.04%、38.28%。到第20 d 时,对照组的病斑直径比处理组分别高出52.14%、66.77%、41.50%。说明Arg 处理可以有效抑制接种A.alternata后枣果实病斑直径的增长。
2.2 Arg 处理对冬枣果实中抗病相关酶活性的影响
PAL 是冬枣抗病过程中的重要酶类,能参与植物抗病过程中多种抗病物质的合成。如图2A,在贮藏期间,20、200 μmol/L Arg 处理组PAL 活性在0~12 d 呈上升趋势,在12~20 d 呈下降趋势,在第12 d时,两个处理组酶活性分别为96.7 和123.4 U·h−1·g−1,对 照 组PAL 活 性 为59.9 U·h−1·g−1,比 对 照 组 高61.4%、106%。1000 μmol/L Arg 处理组和对照组的PAL 活性在0~16 d 呈现上升趋势,16 d 后下降,第16 d 两 组 酶 活 性 分 别 为56.9、70.2 U·h−1·g−1,1000 μmol/L 处理组比20 和200 μmol/L 处理组的活性低21.5%、42.4%;对照组比20 和200 μmol/L处理组的活性低3.4%、29%。
POD 主要参与植物体内酚类化合物的氧化和植物细胞壁中木质素的形成过程,有助于加强细胞壁的屏障作用以限制病害的发展[22],增强植物的抗病性。如图2B 所示,POD 活性在贮藏期间呈现先上升后下降的趋势,各组均在第4 d 出现活性高峰。贮藏期间,200 μmol/L 处理组POD 活性始终高于对照组,而1000 μmol/L 处理组POD 活性始终低于对照组。第16 d 时,20、200 μmol/L 处理组分别比对照组的POD 活性高6.8%、18.9%。
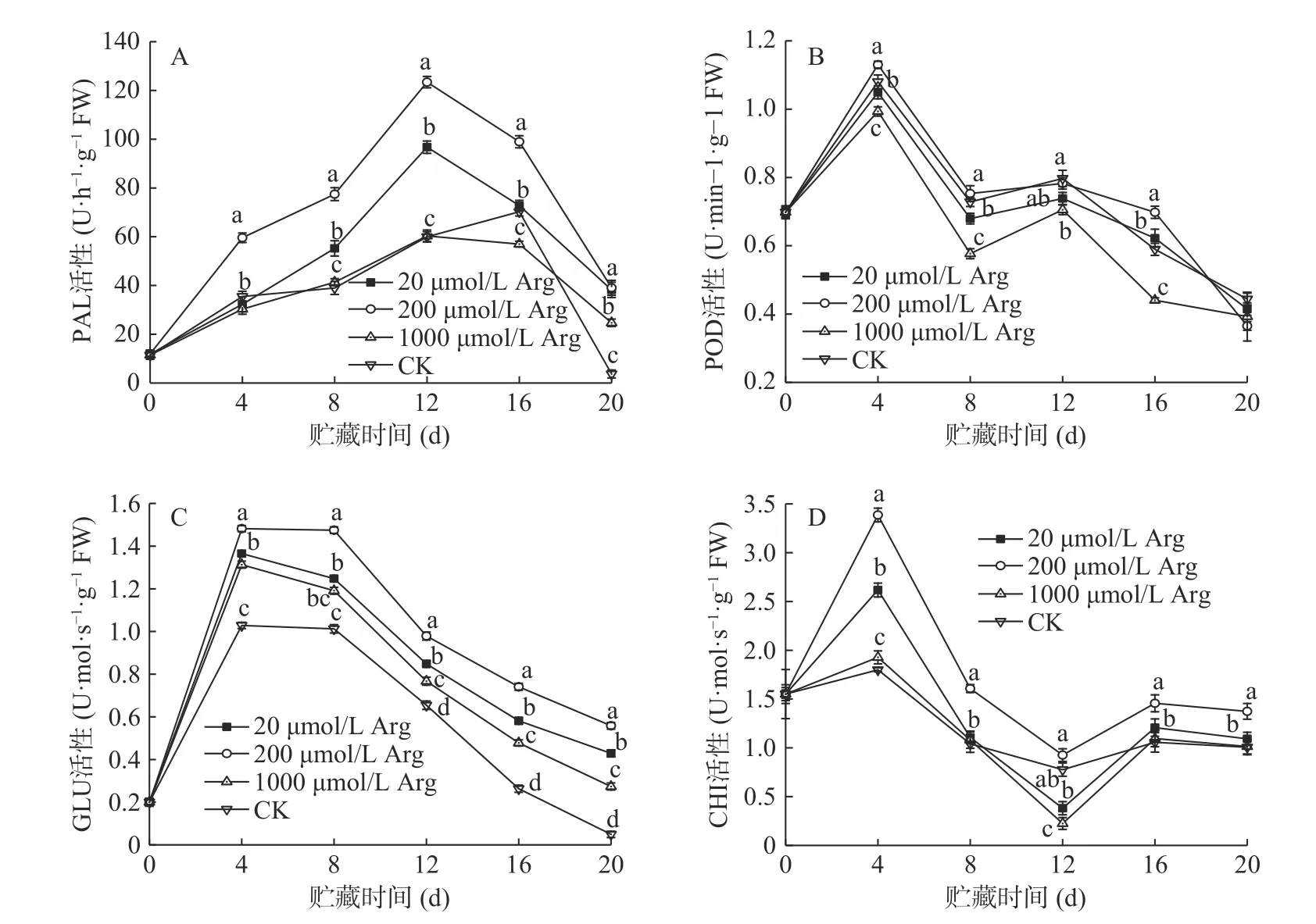
图2 Arg 处理对冬枣果实中PAL(A)、POD(B)、GLU(C)、CHI(D)活性的影响Fig.2 Effect of arginine treatment on the activities of PAL (A),POD (B),GLU (C) and CHI (D) in winter jujube
GLU 是参与果实抗病作用的重要病程相关蛋白,它可以通过水解真菌细胞壁中的葡聚糖来参与植物的防御反应[23−24]。如图2C 所示,在贮藏0~4 d时,GLU 活性急剧上升,在第4 d 时各组活性均达到最大值,之后则呈现缓慢下降的趋势。在第4 d 时,20、200、1000 μmol/L 处理组比对照组的GLU 活性分别高25%、31%、22%。直到20 d 时,各处理组和对照组的GLU 活性仍然有较大差异,各处理组的酶活性分别是对照组的10.5 倍、13.75 倍、6.75 倍。
CHI 直接作用于真菌细胞壁的主要组成成分几丁质,通过破坏真菌的细胞壁结构增强植物的抗病性。如图2D 所示,在处理后0~4 d,酶活性快速上升,4 d 之后,呈现逐渐下降的趋势。在处理前8 d,20、200、1000 μmol/L 处理组的CHI 活性均高于对照组。在第4 d 时,各处理组的酶活性分别比对照组的高45%、87%和6%。总体来看,贮藏期间200 μmol/L处理组的GLU 活性一直高于其他处理组和对照组。
2.3 Arg 处理对冬枣果实中黄酮、多酚和木质素含量的影响
黄酮是植物中重要的抗氧化和抗病物质,黄酮含量的测定在判定抗病作用方面有重要意义。在贮藏期间,黄酮的含量先快速上升,而后变化相对平稳。由图3A 可知,各处理组的黄酮含量均高于对照。到第12 d 时,20、200、1000 μmol/L 处理组和对照组的差异较大,各处理组分别比对照组高37.5%、41.2%、25%。到20 d 时,各处理组的黄酮含量分别为1.18、1.25、1.14 mg·g−1,分别比对照组高6.3%、12.6%、2.7%。
多酚也是植物体内主要的抗氧化物质,在果实对抗采后病原菌中起着重要作用,贮藏期间呈现先上升后下降的趋势,各处理的多酚含量在接种后的整个贮藏期间均高于对照。由图3B 可知,贮藏第8 d时,各组的多酚含量最高,20、200、1000 μmol/L 处理组和对照组的多酚含量分别为2.93、2.99、2.83、2.67 mg·g−1,各处理组分别比对照组高9.7%、11.9%、5.9%。
木质素的沉积可以降低病原菌致病酶对寄主细胞的降解,延缓病原菌对寄主植物的侵染。如图3C 所示,20、200 μmol/L 处理组木质素含量在0~8 d 时缓慢上升,8 d 后逐渐下降,1000 μmol/L 处理组木质素含量在4~8 d 时缓慢上升,8 d 后逐渐下降,而对照组的木质素含量在处理后前16 d 一直缓慢下降,到20 d 时略有升高。贮藏期间,各处理组的木质素含量均高于对照,其中,200 μmol/L 处理组枣果实的木质素含量最高。各处理组均在贮藏第8 d时木质素含量最高,分别为3.68、3.72、3.64 OD280·g−1,分别比对照组高2.5%、3.6%、1.3%。
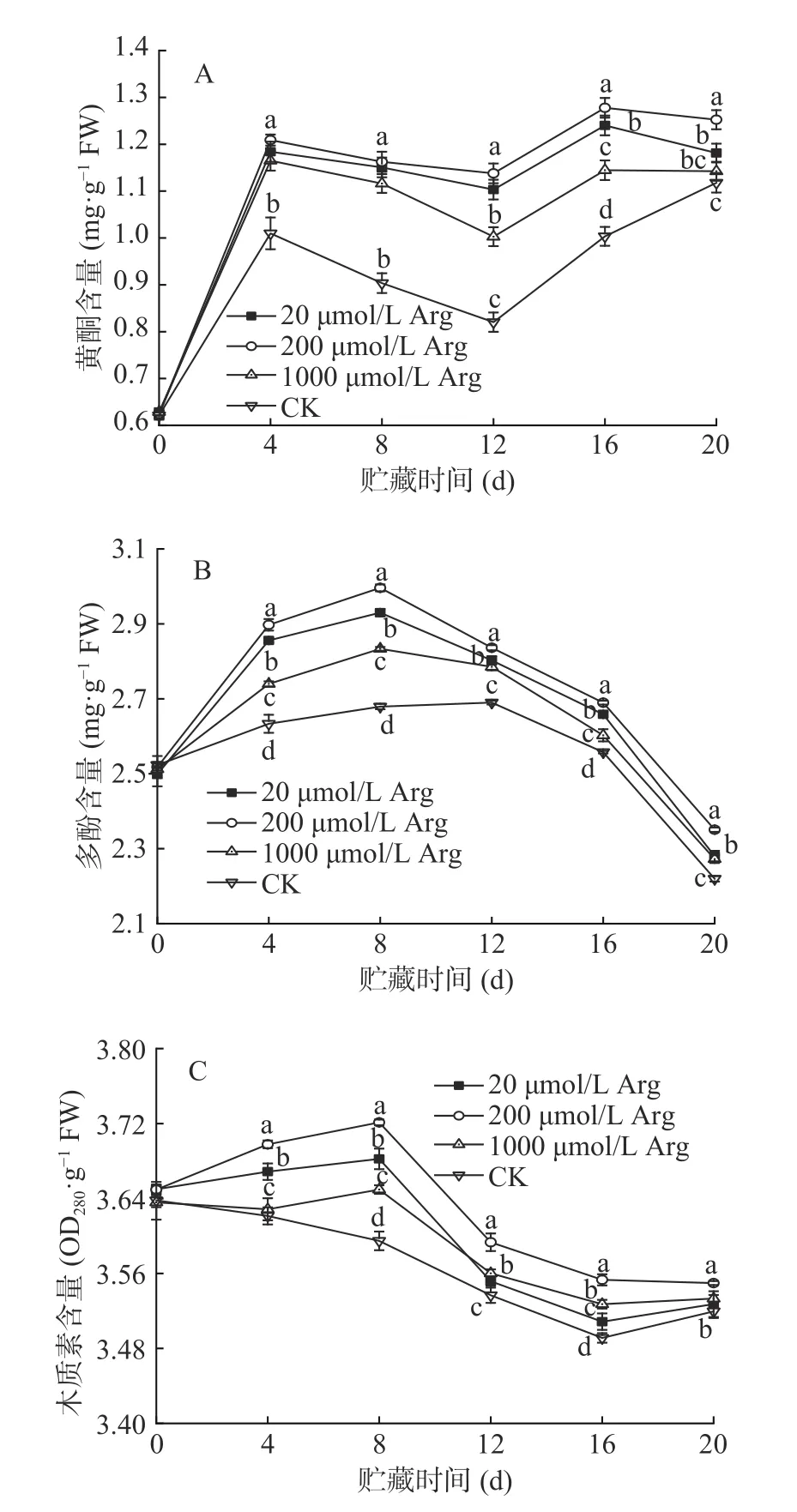
图3 Arg 处理对冬枣果实中黄酮(A)、多酚(B)、木质素(C)含量的影响Fig.3 Effect of arginine treatment on the contents of flavones(A),polyphenols (B) and lignin (C) in winter jujube
3 讨论
Arg 作为人体必需氨基酸,同时是植物体内一种重要的氮素储藏营养物,也是多胺(PA)和NO 这两个重要信使分子的前体物质,与果蔬采后抗性密切相关[22]。在应对不同的生理病害时,植物细胞积聚的PA 通过调控一系列转运过程和生理生化反应来抵抗病害,如PA 作为抗氧化剂和自由基清除剂能减少细胞内的活性氧,防止细胞氧化损伤。NO 作为抗病机制中的信号分子,已被证实与PAL、GLU 和CHI等抗病酶的活性呈正相关[15]。研究表明,Arg 处理可以延缓番茄[15]、白蘑菇[16]的腐烂速度,但是关于Arg 处理诱导采后果蔬抗病性的研究还不深入。
病斑直径能够直观反映果实的抗病作用效果。我们的研究发现,Arg 处理对接种A.alternata后冬枣的病斑直径扩展有明显的抑制作用,各处理组的病斑直径均小于对照组,这与季娜娜在研究精氨酸处理诱导番茄果实抗病性时的发现一致[24]。因此,Arg 浸泡处理对抑制枣果实的黑斑病是有效的。此外,体外实验结果显示Arg 对A.alternata无直接抑制作用(未发表),由此推测Arg 可能通过诱导作用来增强枣果实的抗病性,并进行后续研究。
枣果实的诱导抗病性与果实中抗病相关酶活性的变化密切相关。PAL、GLU、CHI 是植物抗病相关的重要酶类[25]。PAL、GLU、CHI 活性的增加在诱导抗病性中起关键作用,有助于增强果实抵御病原菌入侵的能力,进而抑制病害的发生[26−27]。PAL 是苯丙烷途径的初始通道酶,其活性的增加不仅与抗病有密切关系的黄酮、多酚等物质的合成有关,还参与植物细胞壁木质化,从而直接限制病原菌的生长。GLU 和CHI 是植物组织中两种关键的抗病相关蛋白,二者分别是真菌细胞壁中的几丁质和葡聚糖的水解酶,具有降解真菌细胞壁的能力[28−29]。POD 参与植物细胞壁的构建过程,如酚类化合物的氧化、亚甲基化和木质化,这有助于加强细胞壁结构以限制病原菌的发展[30]。本研究表明,Arg 浸泡处理枣果实后,其PAL、GLU、CHI 活性都有明显上升,表明Arg 处理通过提高抗病相关酶的活性来诱导果实产生抵抗病原菌的能力,这与季娜娜的研究结果一致[24]。而整个贮藏期间对POD 的活性研究表明,适宜浓度的Arg 处理(200 μmol/L)能诱导POD 活性上升。因此,PAL、GLU、CHI 三种酶均参与冬枣果实对A.alternata感染引发的黑斑病的防御反应。
黄酮、多酚、木质素,是植物苯丙烷代谢的重要产物,它们可以保护植物免受病原菌的侵害。黄酮和多酚不但可以直接作为抗菌剂,还可以氧化成醌类物质,进一步抑制病原菌的生长。木质素可以加固细胞壁,形成阻碍病原菌入侵的屏障,阻止病原菌的入侵[31]。本实验结果表明,Arg 处理可以明显提高果实中黄酮、多酚、木质素的含量,增加枣果实对黑斑病的抗性。这与前人在番茄上的研究结果相似。当然,Arg 在果实抗病中的分子作用机制以及具体的信号通路还有待进一步研究。
4 结论
不同浓度的Arg 处理均能有效抑制病原菌的扩展,提高冬枣果实的抗病能力,其中,200 μmol/L Arg 处理组的病斑直径增长最慢,效果最显著。进一步研究显示,Arg 处理提高了枣果实中PAL、GLU、CHI 的活性,其中200 μmol/L Arg 处理组的效果最好,说明Arg 处理通过对枣果实中抗病相关酶活性的诱导,显著增加了果实组织中黄酮、多酚、木质素等抗病相关物质的含量。因此,Arg 能有效提高枣果实对A.alternata感染引发的黑斑病的抵抗能力。
猜你喜欢
热带作物学报(2022年7期)2022-08-06
今日农业(2022年4期)2022-06-01
作物学报(2022年4期)2022-02-10
作物研究(2020年2期)2020-04-04
农产品市场周刊(2020年23期)2020-01-25
今日农业(2019年12期)2019-08-13
发明与创新·大科技(2019年5期)2019-07-31
电子制作(2019年10期)2019-06-17
长江蔬菜·学术版(2016年12期)2017-01-12
营销界(2015年25期)2015-08-21
