
Lipid species dependent vesicles clustering caused by alpha-synuclein as revealed by single-vesicle imaging with total internal reflection fluorescence microscopy
2021-06-08 03:21ChintaManiAryalOwenTyoeJiajieDiao
Biophysics Reports 2021年6期
Chinta Mani Aryal,Owen Tyoe,Jiajie Diao ✉
1 Department of Cancer Biology, University of Cincinnati College of Medicine, Cincinnati, OH 45267, USA
2 Department of Physics, University of Cincinnati, Cincinnati, OH 45221, USA
Abstract Single-molecule methods have been applied to study the mechanisms of many biophysical systems that occur on the nanometer scale. To probe the dynamics of such systems including vesicle docking, tethering, fusion, trafficking, protein-membrane interactions, etc., and to obtain reproducible experimental data; proper methodology and framework are crucial. Here, we address this need by developing a protocol for immobilization of vesicles composed of synthetic lipids and measurement using total internal reflection fluorescence (TIRF) microscopy. Furthermore, we demonstrate applications including vesicle clustering mediated by proteins such as alpha-Synuclein (αSyn) and the influence of external ions by using TIRF microscopy. Moreover, we use this method to quantify the dependence of lipid composition and charge on vesicle clustering mediated by αSyn which is based on the methods previously reported.
Keywords Single molecule, Alpha-Synuclein, Lipids, Vesicles, Clustering, Total internal reflection fluorescence microscopy
INTRODUCTION
The series of events with synaptic vesicles in association with proteins are common in synaptic nerve terminals. For example, synaptic vesicles (SV)transportion, fusion, and recycling processes facilitate the release of neurotransmitters (Espositoet al.2011).These overall processes take place in many steps such as the formation of cluster, docking, tethering,exocytosis and endocytosis cycle. This is, however,possible only after the mediation of cytosolic proteins(e.g., alpha-synuclein (αSyn)) (Murphyet al.2000),membrane bound proteinse.g. soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) (Bonifacino and Glick 2004) and other accessory proteins and ions. Specifically, αSyn w/o SNARE or ions plays a critical role for regulation in clustering (Caiet al.2020; Diaoet al.2013), docking(Laiet al.2014), tethering (Caiet al.2019), fusion (Huet al.2019), and endo/exocytosis (Lautenschlageret al.2017) by stabilizing the vesicles (Kaur and Lee 2021).
αSyn is a functional protein found predominantly in the cytosol of neurons with primarily high abundance in the presynaptic terminal (Stefanis 2012) and is associated with both physiology as well as pathology.Whereas the physiological role involves with synaptic vesicles recycling by interaction with the lipid membrane (Lautenschlageretal.2017); the pathological role is relevant in several synucleinopathies such as dementia. Remarkably, its aggregation into the Lewy bodies becomes the hallmark of Parkinson’s disease (PD) (Spillantiniet al.1997).Indeed, the structural variation (Wanget al.2016) or mutation (Polymeropouloset al.1997) or posttranslational modifications (Wuet al.2020; Buet al.2017) are causative for the functioning of αSyn from physiological function to the toxic condition. It is comprised of 140 amino acids (~14 kDa) and is known to be an intrinsically disordered protein. The structural basis shows that it can be divided into three main regions namely: N-terminus, non-amyloid-beta component (NAB) and C-terminus contributing distinct structural, and dynamical properties for the physiochemical regulation relevant to the content of amino acid sequences (Fuscoet al.2014). Of many functionalities of αSyn such as docking, tethering,fusion, endo/exo-cytosis are influenced by lipid species binding as mentioned earlier. Upon membrane binding,αSyn adopts the structural transition (alpha helix)(Pfefferkornet al.2012). Studies have shown that αSyn has a high tendency of binding only with the anionic lipids like phosphatidylserine (PS) (Middleton and Rhoades 2010), and is preferential to the small and highly curved membrane (Liuet al.2021). However,needs a specific combination of polyunsaturated chains which provides the loose packing (Kuboet al.2005).Other studies have shown that the presence of zwitterionic phospholipid particularly phosphatidylethanolamine (PE) causes the elevated binding (Joet al.2000), whereas Jianjun Panet al.shows the inhibitory effect of PE in membrane remodeling (Panet al.2018). So, the distinct role of specific lipid’s head and chains for αSyn binding remain elusive.
Synaptic vesicles of size ~40 nm contain a significant amount of phospholipids with 12 mole% PS, 23 mole% PE, and several other lipids (cholesterol, spingomyline,phosphatidylinositol, hexylceramide, ceramide,etc.)covering ~50% surface, and is asymmetric in terms of lipid distribution (Takamoriet al.2006). Every lipid in synaptic vesicles has a specific role in association with the αSyn binding (Shvadchaket al.2011). Biophysical studies of mimic synaptic vesicles (Croweet al.2017) or membranes mediated by αSynin vitro(for detail refer to review Candace M Pfefferkornet al.(Pfefferkornet al.2012)) have been studied by using different kinds of model vesicles and bilayer using variable lipids (Panet al.2018). So having an insight into how lipid species and their head or tail are affected by αSyn binding is important to understand the functionalities of synaptic vesicles recycling and pathogenicity caused by this protein.
Several techniques have been employed to study the membrane association of αSyn such as nuclear magnetic resonance spectroscopy (NMR) (Eliezeret al.2001),atomic force microscopy (AFM) and electron paramagnetic resonance spectroscopy (EPR) (Panet al.2018), computer simulation (Westet al.2016), neutron reflectometry (NR) (Hellstrandet al.2013), electron microscopy (EM) (Liet al.2019; Madineet al.2009),etc.Here we study the interaction of αSyn and membrane for lipid specificity at the single-molecule level (Tianet al.2019; Ferreonet al.2009; Gonget al.2016) which overcomes the limitation of ensemble average (Denizet al.2007) by using total internal reflection fluorescence microscopy (TIRFM or TIRF) (Laiet al.2016; Huet al.2017; Duet al.2020). TIRF is often known as evanescent wave or field microscopy providing a high contrast imaging of dynamics at or at the proximity of cellular membrane (Mattheyseset al.2010). The physical basis of this technique is that it utilizes the evanescent field as an excitation field which decays exponentially with distance when the incoming bean passes through the medium with a high refractive index (usually glass) to a low refractive index (buffer or sample) at an angle above the critical angle (Fish 2009). This technique is often compared with epifluorescence microscopy. Unlike epifluorescence microscopy that fits for bulk imaging, it is capable to provide information about the process that happens within ~100 nm (Steyer and Almers 2001). So,utilizing this technique for selective excitation using two solid state lasers (green (532 nm)-Nd:YAG and red(640 nm)-HeNe) has been particularly useful to study the vesicles docking as induced by protein (Caiet al.2019). Moreover, it has been established as a powerful technique to probe several modern processes in cell biology (Mattheyseset al.2010). For instance,peptide—lipid interaction (Foxet al.2009), actin nucleation and elongation (Jiang and Huang 2017),vesicles clustering as mediated by αSyn and regulated by calcium ions (Caiet al.2020), translocation of signaling molecule (Tengholmet al.2003), and docking and priming of vesicles (Bechereret al.2007) have been successfully studied using TIRF. Figure 1 shows the schematic of measurement by prism based TIRF.
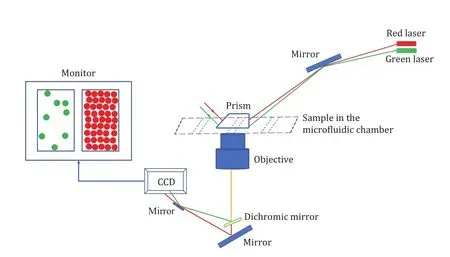
Fig. 1 Schematic illustration of an imaging and detection system of the prism based TIRF
Small unilamellar vesicles (SUV) provide an excellent platform to monitor several biological phenomenain vitro(Huet al.2016; Aryalet al.2020; Panet al.2017;Khadkaet al.2018, 2021). To monitor and quantify the lipid species dependent clustering synaptic mimicking SUV and effectiveness of the technique; in this paper, we report a protocol of small unilamellar vesicles immobilization followed by interaction with free vesicles (Diaoet al.2009) as mediated by synaptic protein αSyn as measured by TIRF. Specifically, vesicles of size ~45 nm with DiD labeled vesicles (DiIC18(5);DiD-vesicles onward) are immobilized and vesicles with DiI labelled (DiIC18(3); DiI-vesicles onward) are floating. The floating vesicles depending upon the lipid species it contains in the presence or absence of divalent cations show the distinct quantitative clustering in the presence of αSyn. Clustering as mediated by αSyn and the influence of divalent cations (Caiet al.2019)particularly the concentration dependent non-linear role of Ca2+has been already studied by Diao’s group(Caiet al.2020). By following a similar protocol, it may be relevant to study the influence of lipid species such as PS at different concentration levels in the clustering of vesicles as mediated by αSyn and modulated Ca2+.Understanding the influence of particular lipid species on the protein induced process in a single vesicles level will provide the kinetic and dynamic insight into the mechanism of action at molecular level in cell membranes (Manet al.2021). The method described here is particularly designed for those who wish to perform the imaging and analysis of protein mediated action at single-molecule level, but could be applied to study the other macromolecule-macromolecule interactions and endocytosis/exocytosis, budding,fusion/fission process,etc. A similar assay has been already discussed to study the conformational dynamics of αSyn during the interaction with the membrane (Maet al.2019) and SNARE (Sunet al.2019) for its functional relevance.
EXPERIMENTAL SECTION
Materials
List of reagents
Reagents used in experiments are listed in Table 1.
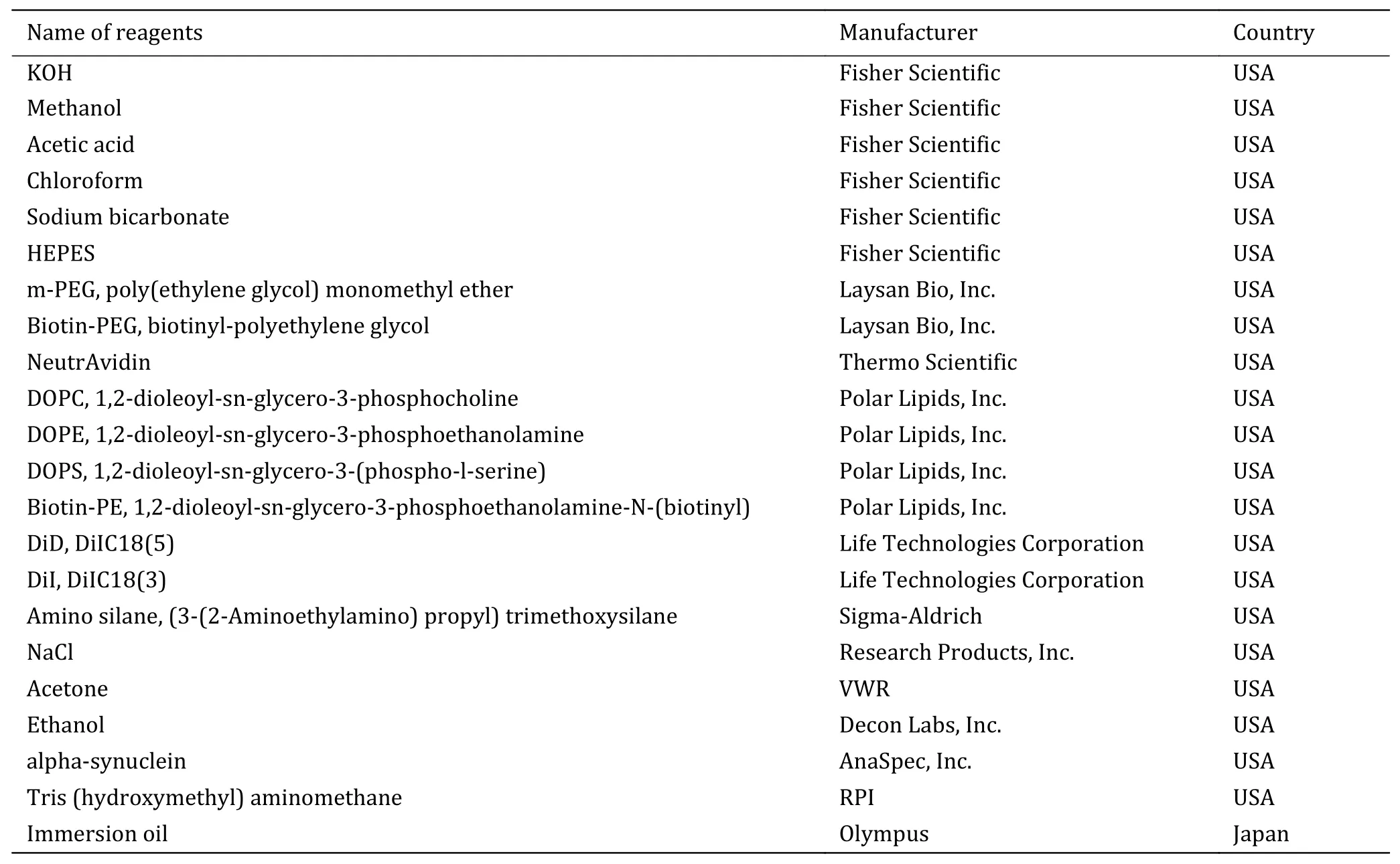
Table 1 List of reagents
List of equipment
Equipment that are used for experiment are tabulated in Table 2.
Table 2 List of equipment
Table of % (mole) lipids in the vesicles
Lipids compositions for both DiD and DiI-vesicles are presented in Table 3.
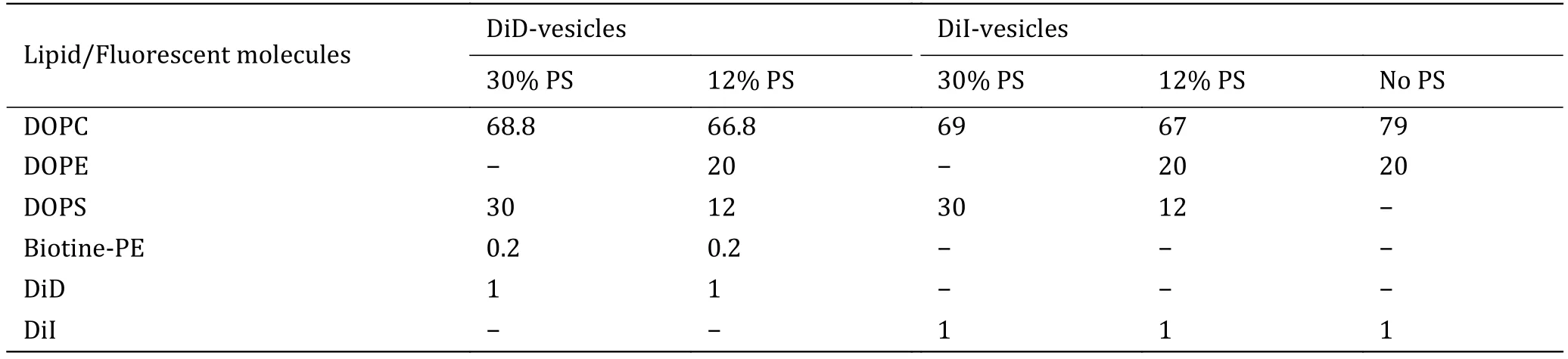
Table 3 Lipid compositions (mol%) for DiD and DiI-vesicles
Procedures
Research design
Based on the initial hypothesis that lipids species influence the αSyn binding (particularly PS, and PE),while the role of Ca2+has been studied (Diaoet al.2009), it will change the ionic strength, suppressing the binding of αSyn at low concentration as a competitive binding. Acknowledging this idea, we experimentally validate the αSyn binding as caused by PE and PS, since both are presented in the synaptic vesicles.Furthermore, it has been reported that whereas phosphatidylcholine (PC) lipid plays little role in binding, PE has relatively higher; this study can be extended to understand the actual role of PE for αSyn interaction in a single molecule level. The experiment covers several tests for a given αSyn binding by identifying the optimized working condition satisfying the physiological relevance. Since experimental evidence shows the presence of anionic lipid enhance the binding of αSyn with vesicles, the project is being initialized with the extreme content of PS (Rhoadeset al.2006; Diaoet al.2009) for example using 30 mole% on both DiD and DiI-vesicles and or reproducing the earlier reported results from our lab on the effect of different concentrations of calcium ions. Below, we outline the methodology and preliminary results.
Sample preparation
1 Surface cleaning and PEGylation
(A) Surface cleaning
The pre-drilled quartz slides and the glass coverslips obtained from vendors must first be thoroughly cleaned. Here, we have described our standard cleaning procedure, but others have used slightly different protocols (Duet al.2021; Lamichhaneet al.2010).Scrub and thoroughly rinse slides with ethanol (100%)and then with Milli-Q water. Since the presence of impurities on the surface increase background fluorescence, scrubbing is required for removing debris from previous experiments. Place slides and fresh coverslips in the clean and dry Coplin staining jars separately. Rinse with Milli-Q water. Sonicate with acetone for 20 min to remove any remaining organic material and non-specifically interacting debris. Again rinse with Milli-Q water three times. Fill jars with 1 mol/L KOH. Sonicate for 20 min followed by rinsing with Milli-Q water three times. Fill jars with methanol and sonicate for 20 min. Burn one side of each quartz slide for at least two mins using a propane torch, then rinse both slides and cover slips with fresh methanol,three times.
(B) Silanization
The next step is silanization which is achieved by aminosilanization with amino silane ((3-(2-aminoethylamino) propyl) trimethoxysilane (APTS)). In aminosilanization reaction, methanol is used as a solvent and acetic acid as a catalyst (Chandradosset al.2014). Mix 100 mL methanol, 5 mL acetic acid and 1 mL APTS reagent in a clean, dry beaker. Pour into the jars and make sure it fully covers slides and coverslips.Incubate for 10 min. Sonicate for 1 min and again incubate for 10 min. Rinse slides and coverslips with methanol, and then at least three times with Milli-Q water. Completely dry the slides and coverslips with blowing air. Note that, amino silane solution should be freshly prepared.
(C) PEGylation
Polyethylene glycol (PEG) is commonly used for surface passivation to prevent non-specific interactions with protein, lipids (Ha and Joo 2002; Joo and Ha 2012a) or vesicles (Wanget al.2019). This is achieved by adding 100 μL of freshly prepared reaction solution (120 mg of mPEG, 4 mg of biotin—PEG, and 700 μL of 0.1 mol/L sodium bicarbonate solution) onto the imaging surface of the slide. Each slide was covered by one clean coverslip to be a set, which was incubated overnight and then rinsed with Milli-Q water, dried, used immediately or stored at 20oC for further use.
(D) Microfluidic chamber assembly
Prepare 4—8 mm wide and ~150 μm deep flow channels. For this place a quartz slide on a flat surface with the PEGylated side facing up. Make a channel on the PEGylated surface by putting double-sided tape over the quartz slide in such a way that the holes are positioned at the center of the channel. Place a coverslip facing the PEGylated surface at the top of taped quartz slide. Remove the extra tape by using a razor blade. Seal the chamber by pressing the coverslip over the area where double-sided tapes are placed using properly mixed epoxy (5 min) and wait at least 20 min for the epoxy to set.
2 Vesicles preparation
SUV were prepared by using the extrusion method discussed elsewhere (Aryalet al.2020). Briefly; Mix the lipids in the appropriate ratio (Table 3) from stock kept at —20oC in a glass tube. For each consecutive lipid pipetation, the syring should be cleaned at least five times. Vacuum the mixture for 4 h to remove the organic solvents (chloroform/ethanol). Rehydrate the obtained dry film with HEPES (25 mmol/L HEPES,100 mmol/L NaCl, pH 7.4 (with NaOH)) buffer. Vertex and sonicate the suspension until the film is fully dissolved. Repeat seven cycles of freezing and thawing by submerging into the water at 50oC and introducing into the dry ice (—79oC) alternatively to obtain the polydispersed unilamellar vesicles solution. Finally,obtain the monodispersed vesicles (size ~50 nm) by extruding the solution (21 times) through the 50 nm polycarbonate membrane. Use the obtained vesicles immediately or keep them at —80oC for future use. Note that, chloroform is a carcinogenic agent, lipid mixing is advised to be conducted under a fume hood. Vesicles preparation steps are summarized in Fig. 2.
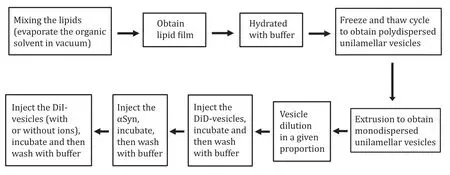
Fig. 2 Flow diagram to show the steps of vesicles and sample preparation
3 Vesicles immobilization and slide preparation
For the measurement by using TIRF, inject 40 μL(two shots of 20 μL) of 0.1 mg/mL NeutrAvidin(Thermo Scientific, 0.1 mg/mL in 10 mmol/L Tris-HCL,pH 7.5, 50 mmol/L NaCl) per channel and incubate for 15 min. This will create a binding surface for biotin labeled samples (DiD-vesicles). Inject the 100 μL of DiD-vesicles with appropriate dilution and incubate for 30 min. This is the basis framework for many types of experiments, but for us, we are interested in vesicle clustering and the interaction with αSyn. Inject 50 μL of αSyn (appropriate concentration) and incubate for 30 min. Inject 100 μL DiI-vesicles with appropriate dilution and incubate for 30 min. The channel without αSyn is considered as the control channel. Note, in each consecutive step, HEPES buffer exchange is required at least five times (200 μL each time) to remove unbound vesicles/protein. Thus, prepared slides were imaged using TIRF under identical intensity.
DiD-vesicles which contain biotin—PE moiety bounds to the neutravidin and hence get immobilized. Since an optimized number of DiD-vesicles provides the homogenous distribution, it can be visualized using the red laser under TIRF. After the confirmation of DiD-vesicles coverage, the protein was injected for the affected channel whereas only buffer was injected in the control channel. Control and affected channel should be prepared within a slide leaving the consecutive channel without any sample to save it if there is any tape to tape leakage, and for better batch to batch comparison. Replication was also done within the same slide for the comparison. Since αSyn has a definite binding ability, the optimal DiI-vesicle concentration depends upon protein. The imaging sequence is tabulated in Table 4.

Table 4 Imaging sequence
After the experiment, the slides can be reused. For recycling, soak the slide in acetone overnight to soften any residue of epoxy or tape from the previous experiment. Use a razor blade to scrape off the residue.Rinse the slides several times with Milli-Q water and dry them.
4 Optimization of working condition
There are many variables to consider to obtain consistent and reproducible results and optimization relies on fixing many of the variables to be as uniform as possible. Beyond the laser alignment, one must optimize the surface density of bound substrate vesicles (DiD-vesicles)e.g., full coverage, homogenous and isotropic distribution in the channel, number(ratio) of DiD and DiI-vesicles for proper concentration of αSyn (pmol/L to μmol/L) binding (Diaoet al.2012),choice of background (high or low), time of incubation(e.g., few minutes to many hours), temperature (about 0oC or room temperature (RT) or 37oC), DiD or DiI-vesicles as substrate, mole% of DiD and DiI fluorescent in a given composition and any surface imperfections on the quartz surface of oil-prism interface which results in the formation of diffraction patterns in the images which in turn make them useless. It is to be noted that, the vesicles count differs from channel to channel, sample to sample and batch to batch, so direct comparison of the count may not be possible.
5 Buffer preparation
PEG buffer: this is prepared freshly for immediate use.It contains 0.1 mol/L sodium bicarbonate (pH 8.5).
HEPES buffer: this can be used for up to one month if stored at 4oC. It contains 25 mmol/L HEPES and 100 mmol/L NaCl (pH 7.4 with NaOH).
T50 buffer: this can be stored at 4oC to be used for a month. It contains 10 mmol/L Tris-HCl and 50 mmol/L NaCl (pH 8.0 with NaOH).
TIRF imaging
The sample is mounted in the following sequence for the imaging: DI water drop at the top of objective,sample coverslip down in contact with DI water,immersion oil on the surface of quartz slide, and finally the prism at the top of the oil. The prism is held in place with a frame and screw. The photograph of the TIRF system used for this experiment is as shown in Fig. 3.

Fig. 3 Photograph of the different parts of the TIRF system in the lab
For reproducibility, at least ten different randomly selected sites were scanned for each sample flow channel and at least three channels a day were scanned.The experiment was conducted at least three different days with freshly prepared vesicles.
Image processing and quantification
Fluorescence signals or images, as well as real time recording, can be taken and analyzed using separate software, but we do both using the custom-built software obtained from Dr. Taekjip Ha’s group (John’s Hopkins University, USA). Statistical results and plots are typically obtained in external programs, such as Excel, MATLAB, Python, Mathematica,etc. Protein induced changes are analyzed by using the studentttest. Ifp< 0.05, the change is considered statistically significant.
Imaging sequences
The following sequence (Table 4) was implemented in the imaging process.
Imaging troubleshoot
Here are some issues experienced and solutions associated with the imaging by TIRF and samples during the experiments (Table 5). For more details,please refer to our previous publication (Diaoet al.2012).
EXPECTED RESULTS
Figure 4 shows the representative images of clustering of vesicles as an effect of αSyn or ion binding under the excitation of green laser. We quantify the observation by measuring the number of DiI-vesicles as a result of αSyn binding as a function of lipid head charge and ion concentration. As the measurement is done with various kinds of samples in the optimization process with different αSyn concentrations, we present the representative data here. Figure 5A shows the change in the clustering count of DiI-vesicles on the top of DiD-vesicles (both contain 12 mol% PS) as the effect of αSyn. Where-as an enhanced count can be observed for the same set of vesicles with the calcium ion as compared to both “without αSyn” and “with αSyn” as shown in Fig. 5B. Experiments were also performed in high background. Figure 5C and 5D show the significantly increased count as a result of αSyn binding in 30 mol% PS containing DiI-vesicles and the 12 mol% PS DiI-vesicles respectively, where, 30 mol% PS containing DiD-vesicle was the substrate. Furthermore,No PS DiI-vesicles (on 30 mol% PS DiD-vesicle substrate) show less enhanced count even in the presence of αSyn (Fig. 5E). However, No PS DiI-vesicles on 12 mol% PS DiD-vesicles show a pretty much similar count as background (Fig. 5F). With these preliminary results, it can be said that; αSyn shows a strong binding affinity with the lipids having charged head specifically PS, and highly enhanced in the presence of a higher concentration of calcium ion is in good agreement with the previous finding (Caiet al.2020).
SUMMARY
Surface passivation and immobilization strategy and measurement by TRIF have been widely accepted technique to study the several biophysical interactions in a single molecule level (Joo and Ha 2012b;Lamichhaneet al.2010). This technique and protocol can be generalized in different ways including to study the interacting behavior of protein and lipid in real time and to probe the complex system of proteins and lipids relevant to physiological conditions by using multicolor fluorophore/excitation. Furthermore, TIRF coupled with other high-resolution techniques and single level spectroscopies to probe molecular mechanisms and dynamics in the qualitative as well as quantitative manner. Here, we have described the detailed protocol in the quantification of the role of lipid’s charge or ion on clustering behavior caused by αSyn by using TIRF aiming to extend our study in αSyn’s binding with several other lipids that are physiologically relevant for synaptic vesicles. Since this assay has been already proven to be useful in elucidating the conformational dynamics of a protein in protein—membrane interaction and RNA/DNA—protein interaction in conjugation with single-molecule forster resonance energy transfer, it can be applied to study other protein—lipids, protein—protein, and protein—ion interactions. One immediate and potential application of this protocol could be to distinguish between the forces such as charges versus hydrophobicity and imperfection that govern the protein interaction with lipids species. Furthermore, this protocol could be extended in thein vitrobiophysical studies of protein reconstituted system of vesicles (e.g., segment of SNARE) for their relevancy of docking, fusionetc. with the membrane.
AcknowledgementsThis work was supported by the National Institute of Heath (R35GM128837, R21AG061600,R01NS121077) to Jiajie Diao.
Compliance with Ethical Standards
Conflict of interestChinta Mani Aryal, Owen Tyoe and Jiajie Diao declare that they have no conflict of interest.
Human and animal rights and informed consentThis article does not contain any studies with human or animal subjects performed by any of the authors.Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use,sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.

- Biophysics Reports的其它文章
- Quantitation of nucleoprotein complexes by UV absorbance and Bradford assay
- Optical tweezer and TIRF microscopy for single molecule manipulation of RNA/DNA nanostructures including their rubbery property and single molecule counting
- Molecular mechanisms of Streptococcus pyogenes Cas9:a single-molecule perspective
- Real-time imaging of structure and dynamics of transmembrane biomolecules by FRET-induced single-molecule fluorescence attenuation
- A review for cell-based screening methods in drug discovery