
无核玫瑰香味葡萄种质创新及胚挽救技术优化
2021-06-07 09:16陈烁李莎莎骆强伟徐炎王跃进
中外葡萄与葡萄酒 2021年3期
陈烁,李莎莎,骆强伟,徐炎*,王跃进*
(1. 西北农林科技大学园艺学院/旱区作物逆境生物学国家重点实验室/农业部西北地区园艺作物生物学与种质创制重点实验室,陕西杨陵 712100;2. 新疆维吾尔自治区葡萄瓜果研究所,新疆鄯善 838200)
葡萄的质量和营养成分以及无核葡萄新品种的育种一直是重要的研究课题[1-2]。无核葡萄因其滋味甜美、即采即食等原因成为消费者选择鲜食水果和果脯的首选。香味是影响果实品质的重要因素。目前,能在市场上被广泛推广和栽培的无核且具有香味的葡萄品种还非常有限,因此将无核和香味两个性状叠加于一体的新品种是葡萄育种研究中的重要目标。玫瑰香味是葡萄品种独特的气味,该香气与萜类化合物质的存在有关,尤其是由于芳樟醇、香叶醇、橙花醇、香茅醇和a-萜品醇等萜类化合物的成分在香味葡萄中浓度较高,是调控香味的主要物质[3-4]。在植物中已鉴定出两种独立的萜类化合物合成途径:萜胞质的甲羟戊酸途径(Mevalonate pathway, MVA)和质体的2-甲基D-赤藓糖醇-4-磷酸途径(Methyl-erythritol-4-phosphathway,MEP),而MEP途径被认为是葡萄果肉中单萜底物生物合成的主要途径[5]。MEP合成途径前期的关键酶1-脱氧-D-木酮糖-5-磷酸合酶(DXS)、1-脱氧-D-木酮糖-5-磷酸还原异构酶(DXR)和4-羟基-3-甲基丁-2-烯基二磷酸还原酶(HDR)共同作用生成所有萜类化合物的前体,异戊烯基二磷酸(Isopentenyl pyrophosphate, IPP)及其异构体二磷酸二甲基烯丙酯(Imethylallul pyrophosphate,DMAPP)[6]。在合成途径中期,IPP和DMAPP通过异戊二烯基转移酶、香叶基磷酸二氢酯合酶(gernayl diphosphate synthase, GPPS)的作用缩合,生成香叶基磷酸二氢酯(gernayl diphosphate, GPP),不同单萜合酶在晚期催化GPP转化为环状和非环状单萜[7]。对葡萄基因组测序预测到一些与萜类合成相关的酶基因[8-9],其中合成里那醇、香叶醇和a-萜品醇等萜类物质的酶基因功能已被验证。
果树胚挽救的初次成功是在樱桃中进行离体胚培养[10],随后胚挽救技术得到发展。Ramming通过离体胚培养成功培育了无核葡萄植株[11]。为了适应生产要求和育种需求的增加,胚挽救技术也在不断完善。影响该技术的因素主要有:亲本基因型、取样时间、培养基成分等[12]。前人对影响胚珠发育的培养条件进行了研究,成功获得胚挽救杂交苗[13-14]。王跃进等[15]利用无核葡萄胚挽救技术培育‘无核×中国野生葡萄’的胚珠,并研究了采样时间、不同培养基及添加激素、接种方式对胚珠发育的影响。潘学军[16]获得了幼胚发育的最佳培养基为MM3+10 mmol/L甘氨酸。田莉莉[17]认为,可胚挽救的母本材料对杂种胚的萌发和成苗影响较大,‘底来特’(Delight)、‘爱莫无核’(Olmo Seedless)、‘红宝石无核’等无核品种适合在杂交中作母本,成苗率相对较低的‘无核紫’‘无核白’‘杨格尔’(Youngle)等品种不宜作母本。唐冬梅[18]通过对比取材时间认为,在一定范围内,较晚取材接种的胚珠成苗率较高,且通过喷施植物生长调节剂可以促进胚珠发育。贾姗姗[19]在胚萌发培养基WPM中添加激动素(KT)和玉米素(ZT),促进了无核香味杂种胚幼苗根的生长。
通过研究不同香味葡萄品种果实成熟期控制香味物质的萜类合成关键酶基因的表达,筛选出具有萜类物质合成酶基因高表达的香味品种作父本,以种子败育型葡萄作母本,利用胚挽救育种技术培育无核玫瑰香味优质品种,为无核香味葡萄育种奠定材料基础。通过比较亲本基因型对胚挽救技术的影响,筛选与优化胚萌发培养基及确定不同母本最适采样时间,提高无核玫瑰香味葡萄胚挽救效率。利用分子标记技术对杂种F1代的无核性状进行早期辅助选择,以提高杂种后代无核选育效率。
1 材料与方法
1.1 试材及取样
试验于2019年5月—2020年6月在新疆维吾尔自治区葡萄瓜果研究所(吐鲁番市鄯善县,北纬42.91°,东经90.30°)、西北农林科技大学葡萄种质资源圃及旱区作物逆境生物学国家重点实验室完成。
用于萜类物质合成关键酶基因实时定量PCR的葡萄品种有:浓香型品种‘莎巴珍珠’‘阳光玫瑰’‘玫瑰香’‘183’,淡香型品种‘巨玫瑰’‘早玫瑰’‘意大利’及无香型品种‘无核白’等,均定植于西北农林科技大学葡萄种质资源圃中。
用于胚挽救杂交的试材中,母本为9个种子败育型无核葡萄品种:‘爱神玫瑰’‘昆香无核’‘京早晶’‘火焰无核’‘美丽无核’‘莫丽莎无核’‘红宝石无核’‘Fresno Seedless’‘Australian Seedless’,父本为5个玫瑰香味品种‘玫瑰香’‘阳光玫瑰’‘巨玫瑰’‘183’‘贵妃玫瑰’。以上杂交试验于新疆维葡萄瓜果研究所进行,供试葡萄品种均采用水平棚架栽培,株距1 m,行距5 m。胚挽救杂交组合配置见表1。
1.2 萜类物质合成关键酶基因表达分析
以‘莎巴珍珠’等8个葡萄品种为试材,根据不同品种成熟情况,2019年7月下旬至9月上旬采集各品种成熟果实。称取采集的成熟果实300 g,剥取果皮0.5~1 g置于2.0 mL无酶离心管后标号并迅速放入液氮中速冻,后放入超低温冰箱备用。全程在冰上操作,防止RNA降解,其中果皮剥离时尽可能不粘连果肉。使用RNAperp Pure多糖多酚植物总RNA提取试剂盒(天根)提取RNA,参照cDNA第一链合成预混试剂盒(Cofitt)使用说明逆转录成cDNA,置于超低温冰箱备用。以cDNA为模板,根据萜类物质合成通路中各阶段关键酶基因序列,利用软件Primer 6设计引物(表2)并进行实时定量PCR检测。引物参照Martin[8]的方法,使用VvActin和VvGAPDH2个看家基因作为内参。实时定量PCR进行3次生物学重复,其数据分析利用IBM SPSS Statistics 22.0软件进行单因素方差分析,显著性分析用Duncan's检验(P<0.05)。
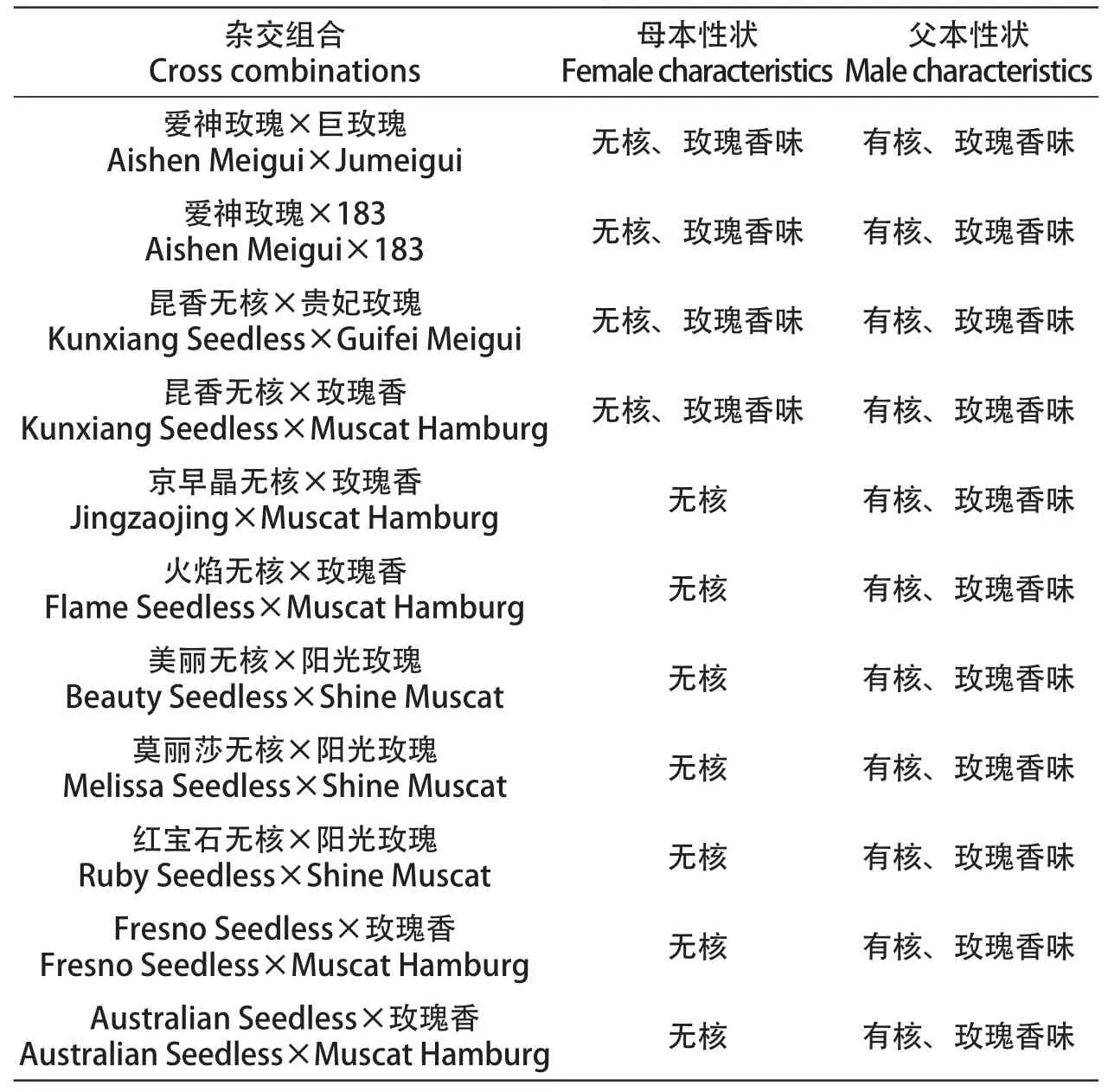
表1 无核香味胚挽救杂交组合配置Table1 Configuration of hybrid combinations
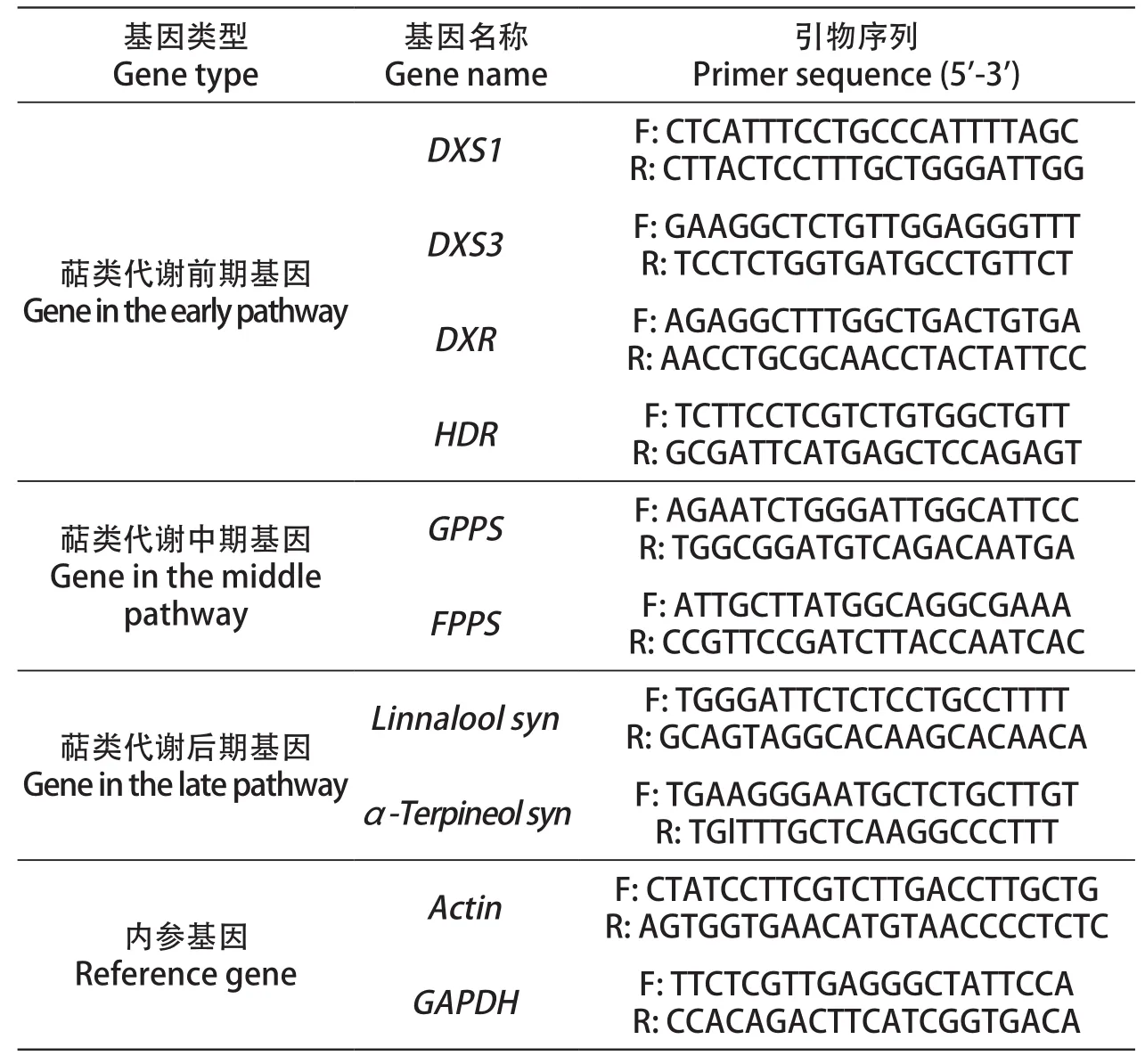
表2 试验所需引物序列Table 2 Primers used in the experiment
1.3 胚挽救流程
采集并研磨花粉,密封保存并放于4 ℃冰箱备用。选择长势强壮的葡萄植株,采集粗壮结果母枝上发育良好的花序,用指尖轻轻掐去花冠顶盖部,使花药脱落,露出柱头,避免伤害柱头。去雄完成后,清理花穗上残余花粉并套袋标记。去雄2~3 d 后,开始授粉,且有黏液时授粉最好。授粉时间以晴天上午8:00—10:00最佳,授粉结束应立即套袋。连续授粉3~4 d。不同杂交组合以最后一次授粉时间为准,在最适采样期将整穗果实采回实验室。从果柄处将果粒分别剪下,装入网兜内置于流水下冲洗3~4 h。在无菌条件下,将消毒干净的果粒在玻璃培养皿中剥取胚珠,将剥离的胚珠接种在MM3固体胚发育培养基中培养。暗培养8~10周后,无菌条件下利用解剖镜剥取杂种胚,将胚平铺在WPM胚萌发培养基中光照培养,温度为25±2 ℃。1个月后统计胚萌发数,2个月后统计成苗数,进行数据统计,胚挽救具体程序参照本课题组前期研究方法[18,27-28]。胚发育率(%)=发育胚数/接种胚珠数×100,成苗率(%)=成苗数/接种胚数×100。
1.4 不同母本最佳采样时间的确定
对‘京早晶’和‘Fresno Seedless’为母本的无核香味胚挽救杂交组合进行连续取样,采样时间为授粉后36~44 d,取样间隔一天。将采下的果穗做好取样时间及组合名称标记并放在冰盒中保存,带到实验室于4 ℃冰箱备用。按照胚挽救流程统计数据并计算不同取样时间下各组合的胚发育率和成苗率。
1.5 胚萌发培养基最适芸苔素内酯浓度筛选
试验共设置了26种不同的胚萌发培养基,以WPM为基础培养基(WPM 2.41 g/L+蔗糖20 g/L+琼脂7 g/L+活性炭1.5 g/L),分别添加不同浓度梯度的芸苔素内酯(brassinolide, BR)作胚萌发培养基并进行优化。比较不同培养基中胚的萌发率,筛选最适胚萌发的BR添加浓度,统计胚萌发率。
1.6 分子标记辅助选择无核葡萄胚挽救杂交子代
杂交亲本及子代基因组DNA的提取,无核探针GSLP1-569及无核标记SCF27-2000的引物序列及反应程序参考李志瑛[24]的方法。
2 结果与分析
2.1 果实中单萜类物质生物合成相关基因表达水平
对8个葡萄品种成熟果实中萜类物质合成关键酶基因进行实时荧光定量PCR检测(图1),其中7个为玫瑰香味品种,1个无香味品种作为对照。定量结果显示,调控萜类物质合成途径前期的关键酶基因DXS1、DXS3、DXR及HDR均在浓香型品种中表达量最高。其中,DXS1及DXS3在‘阳光玫瑰’中表达量最高,DXS1在‘阳光玫瑰’中的表达量是其它品种的3~5倍;DXS3除在‘阳光玫瑰’中高表达外,在‘玫瑰香’中也有较高表达;DXR在‘183’中表达量最高,而在淡香型‘意大利’中和无香型‘无核白’中表达量较低;HDR在‘莎巴珍珠’及‘玫瑰香’成熟果实中表达量都较高。调控萜类物质合成途径中期的关键酶基因GPPS及FPPS在不同品种中表达呈现差异,GPPS在‘183’‘玫瑰香’‘阳光玫瑰’等浓香型品种中都有较高表达,而在‘意大利’中的表达量最低;FPPS在‘阳光玫瑰’及‘早玫瑰’中表达量最高,在‘阳光玫瑰’的表达量是其它品种的1~2.5倍,但其在部分浓香品种如‘玫瑰香’与‘莎巴珍珠’果实中的表达量并不高。调控萜类物质合成途径晚期的关键酶基因Linnalool syn和α-Terpineol syn在不同品种间的表达差异较大,Linnalool syn在‘阳光玫瑰’中的表达量最高,在‘早玫瑰’中的表达量次之;α-Terpineol syn在‘183’中的表达量显著高于其它品种。α-Terpineol syn相较于其它酶基因在各成熟葡萄果实中的表达量也是最高的,是其它酶基因表达量的10~65倍。
2.2 亲本基因型对无核香味葡萄胚挽救的影响
2.2.1 不同母本基因型对胚挽救的影响
试验设置11个无核香味胚挽救杂交组合,共获得杂种胚珠9731个,杂种苗511株。胚发育率在3.39%~55.63%,成苗率在6.88%~38.82%。
以‘阳光玫瑰’作父本与以‘红宝石无核’及‘莫丽莎无核’作母本的杂种胚成苗率均高于以‘美丽无核’作母本的杂种胚,其成苗率分别达到38.82% 和18.92%。此外,在以‘玫瑰香’作父本配置的香味无核杂交组合中,‘Fresno Seedless’及‘Australian Seedless’作母本的组合成苗率高于以‘昆香无核’‘京早晶’及‘火焰无核’为母本的杂交组合,二者成苗率最高,分别为22.03%和27.60%。父本相同时,以‘红宝石无核’作母本的杂种胚成苗率最高,可达到38.82%(表3)。
2.2.2 不同父本基因型对胚挽救的影响
以‘爱神玫瑰’作为母本与不同香味父本的组合中,‘183’作父本的组合成苗率略高于以‘巨玫瑰’作父本的组合,成苗率为17.07%。以‘昆香无核’作母本与不同香味父本杂交,‘玫瑰香’作父本的组合成苗率(18.75%)明显高于以‘贵妃玫瑰’作父本的杂交组合。母本相同时,以‘玫瑰香’作父本的杂种胚成苗率最高,可达到18.75%(表3)。
2.3 取样时间对无核香味葡萄胚挽救的影响
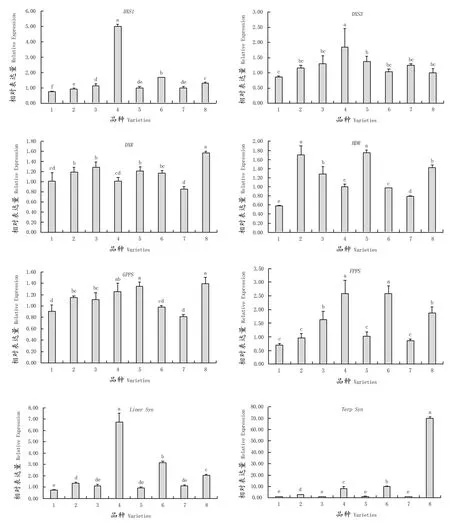
图1 成熟果实中萜类物质合成酶相关基因表达Figure 1 Expression of monoterpene synthase related genes in mature berries
胚挽救杂交组合‘京早晶×玫瑰香’在授粉后第42天胚发育率最高,达到13.33%;授粉后的36 d,成苗率最高,为18.18%;而在授粉后第38天,胚发育率为0。在不同取样时间成活的杂交苗长势均健康,根系发达。以‘Fresno Seedless’作为母本的杂交组合‘Fresno Seedless×玫瑰香’在授粉后的36 d胚发育率达到最高,为31.25%;同一时期的成苗率也最高,为33.33%;授粉后的40 d胚发育率最低,为13.68%(表4)。
2.4 不同浓度BR对无核香味葡萄胚挽救的影响
以‘红宝石无核×阳光玫瑰’的杂种胚为材料,通过添加不同浓度的BR和6-BA对杂种胚萌发率的影响(表5)。与CK1相比,T2、T3处理对胚萌发有促进作用,分别高15.71%、21.59%,以T3处理效果最显著,即生长调节剂的最适浓度为:0.1 mg/L BR+0.2 mg/L 6-BA。随着BR浓度升高,胚发育率有下降趋势。当将6-BA替换为IAA,对比不同浓度梯度的BR和IAA对胚挽救育种的影响时,T7处理组对杂种胚萌发率的促进作用最明显,比CK2高出14.30个百分点,即生长调节剂的最适浓度为BR 0.05 mg/L。而在改变BR与IAA的浓度进行不同处理时,其对应胚萌发率均低于对照组,没有起到促进作用(表5)。T3与T7处理下长成的幼苗长势均健壮,没有出现畸形苗现象。
2.5 利用分子标记鉴定胚挽救杂交子代的无核性状
2.5.1 无核探针GSLP1-569检测胚挽救亲本及杂交子代的无核性状
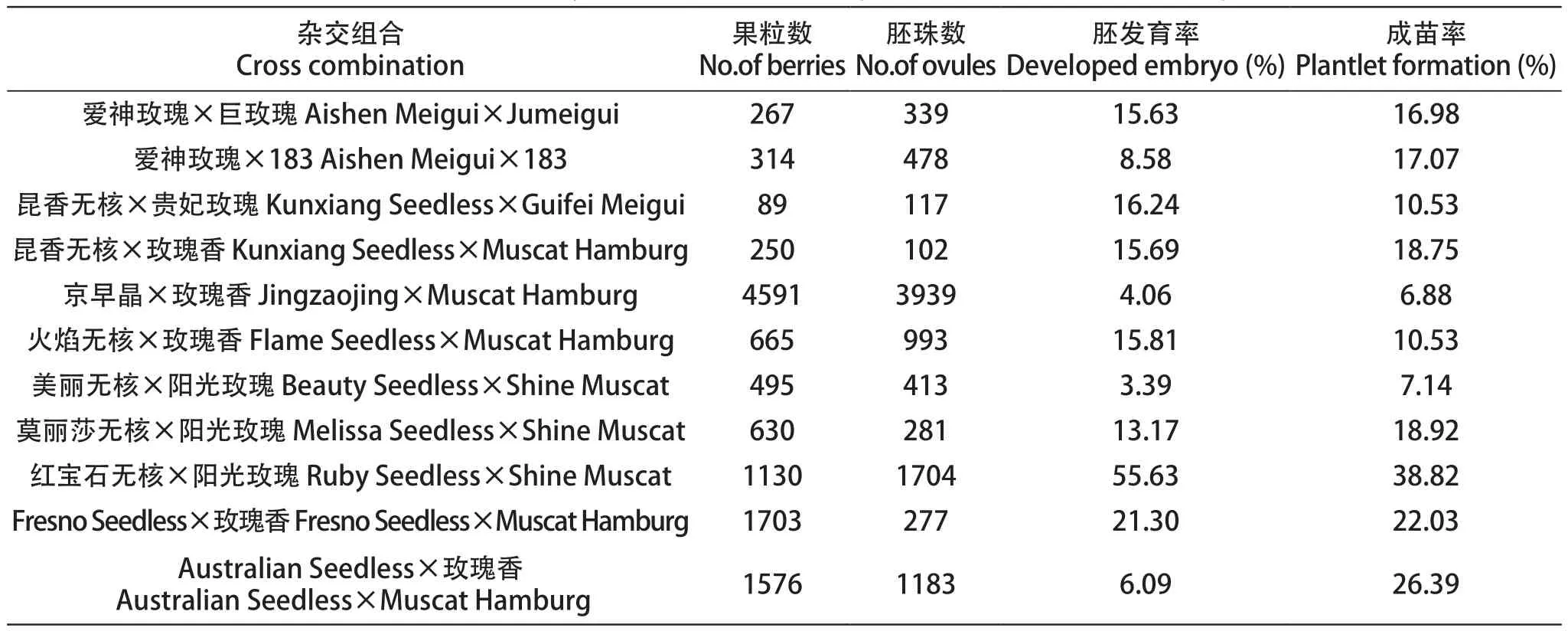
表3 无核香味葡萄胚挽救发育成苗情况Table 3 The result of embryo rescue for breeding seedlessness with aroma grapevine
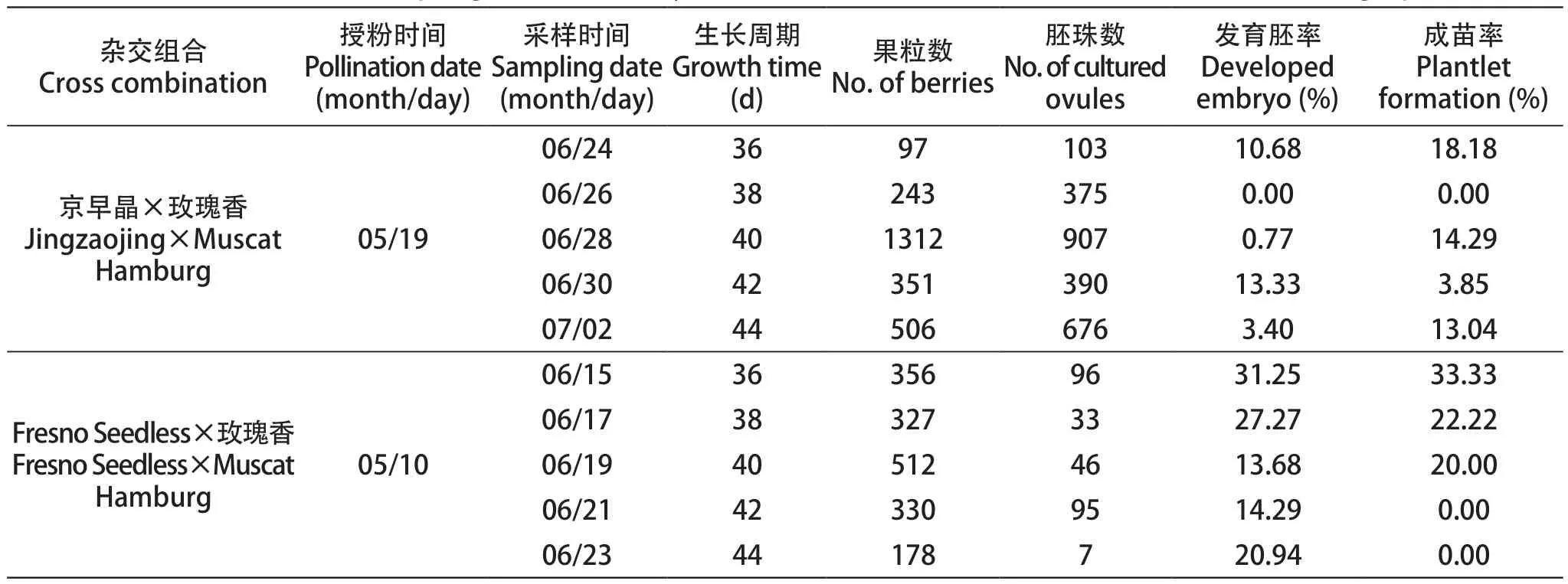
表4 采样时间对无核香味葡萄胚挽救育种结果的影响Table 4 The effect of sampling time on embryo rescue for innovation of seedlessness and flavor grapevine
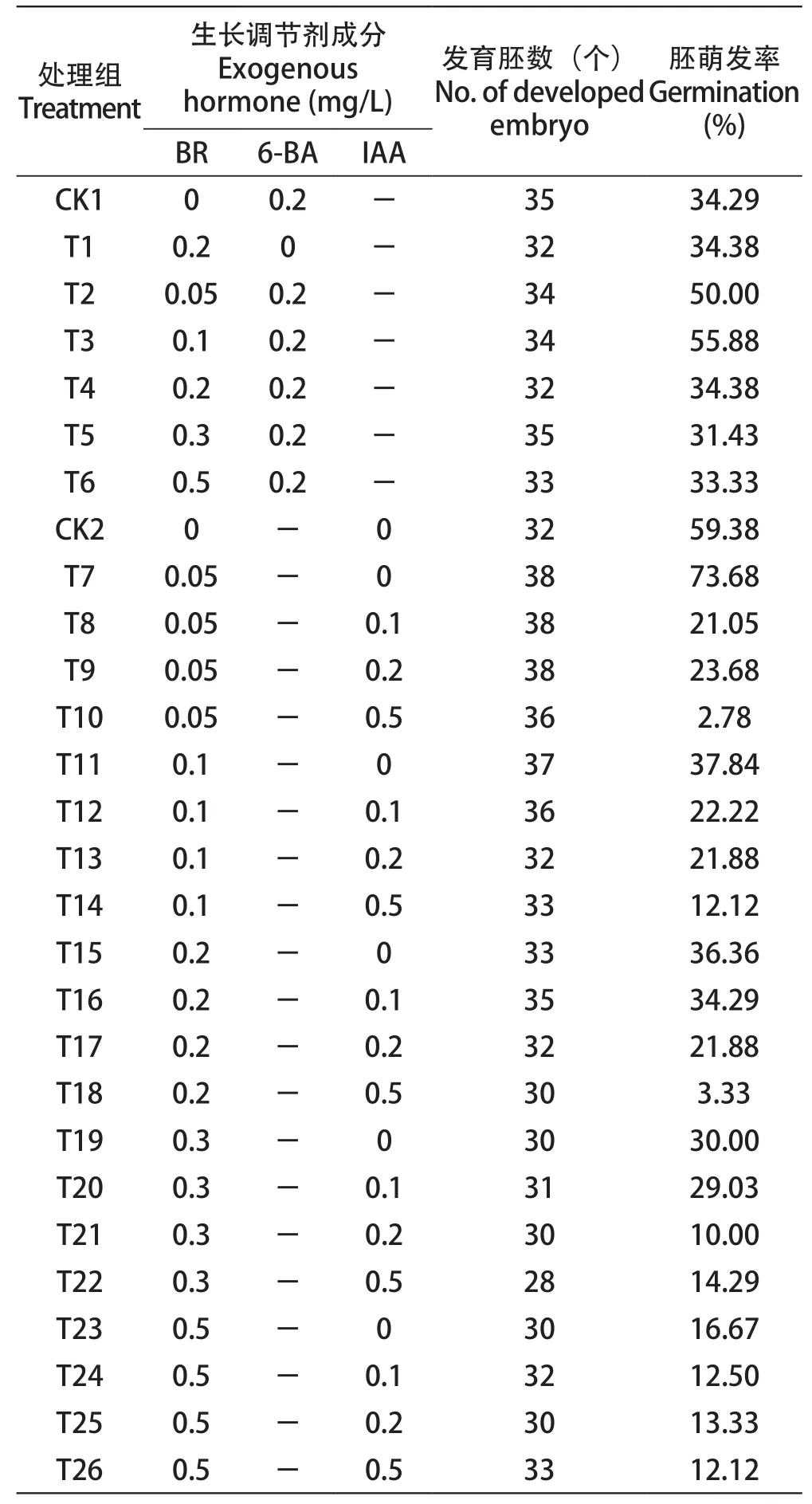
表5 不同浓度BR对胚挽救育种的影响Table 5 Effects of BR at different concentrations on embryo rescue breeding
利用无核探针GSLP1-569对杂交组合中14个双亲进行无核性状筛选。其中可扩增出569 bp条带的母本品种有:‘火焰无核’‘美丽无核’‘Fresno Seedless’和‘Australian Seedless’4个品种;未出现特异性条带的父本品种有:‘巨玫瑰’‘玫瑰香’‘贵妃玫瑰’和‘183’4个品种(图2A)。即无核探针GSLP1-569可用于检测‘火焰无核×玫瑰香’‘Fresno Seedless×玫瑰香’及‘Australian Seedless×玫瑰香’杂交后代无核性状。
使用无核探针GSLP1-569对‘火焰无核×玫瑰香’的8个杂交子代的检测结果中,扩增出569 bp特异性条带的共5个株系(图2B);对‘Fresno Seedless×玫瑰香’的8个杂交子代的检测结果中,其8个子代中全部扩增出569 bp特异性条带(图2C);对‘Australian Seedless×玫瑰香’的8个杂交子代的检测结果中,全部扩增出569 bp特异性条带(图2D)。
2.5.2 无核标记SCF27-2000检测胚挽救亲本及杂交子代的无核性状
利用无核标记SCF27-2000对杂交组合中14个双亲进行无核性状筛选,可扩增出2000 bp条带的母本品种有:‘美丽无核’‘红宝石无核’和‘Fresno Seedless’3个品种;‘巨玫瑰’‘183’‘贵妃玫瑰’‘阳光玫瑰’和‘玫瑰香’5个父本品种均未扩增出2000 bp的特异性条带(图3A),即无核探针SCF27-2000可用于检测‘美丽无核×阳光玫瑰’‘Fresno Seedless×玫瑰香’及‘红宝石无核×阳光玫瑰’杂交后代无核性状。
使用无核分子标记SCF27-2000对‘美丽无核×阳光玫瑰’7个杂交子代的检测结果中,扩增出2000 bp特异性条带的共5个株系(图3B);对‘Fresno Seedless×玫瑰香’8个杂交子代的检测结果中,扩增出2000 bp特异性条带的共6个株系(图3C);对‘红宝石无核×阳光玫瑰’子代的检测结果中,其17个子代中11个株系扩增出2000 bp特异性条带(图3D)。
3 讨论与结论
无核香味葡萄的育种已经开展了很多年,但少有新品种被推出,其主要原因是对香味形成机理的研究较少。目前,已知的成果是萜烯类物质是玫瑰香味的主要呈香物质。前期贾姗姗已经对葡萄品种的果实香味物质含量进行了测定,筛选出‘亚历山大’‘阳光玫瑰’‘玫瑰香’等玫瑰香味物质含量相对较高的品种[19]。在此基础上,分别对不同香型的葡萄品种中控制萜类物质合成关键酶基因进行表达分析,8个酶基因在不同香型葡萄品种中的表达具有差异,总体呈现为在浓香型品种中表达量高于淡香型品种,无香品种基因表达量最低,其中以‘阳光玫瑰’‘玫瑰香’‘183’等品种较为突出。在后期胚挽救杂交组合配置中可优先选择这些品种作为香味父本,而各萜类物质关键酶基因在‘意大利’中的表达均不高,不建议选择其作为亲本。值得注意的是,MEP代谢通路晚期的酶基因Linnalool syn和α-Terpineol syn的表达量显著高于代谢途径前、中期的酶基因。Linnalool syn和α-Terpineol syn分别在萜类合成通路中调控里那醇和萜品醇的合成,果实在成熟期香味最为浓郁,果实内积累大量萜类物质,故而相关酶基因的表达量显著高于通路前、中期的相关酶基因,可由此作为开发香味基因分子标记的参考依据,对胚挽救获得的杂种后代进行香味基因分子标记辅助选择,提高优质品种的选育效率。
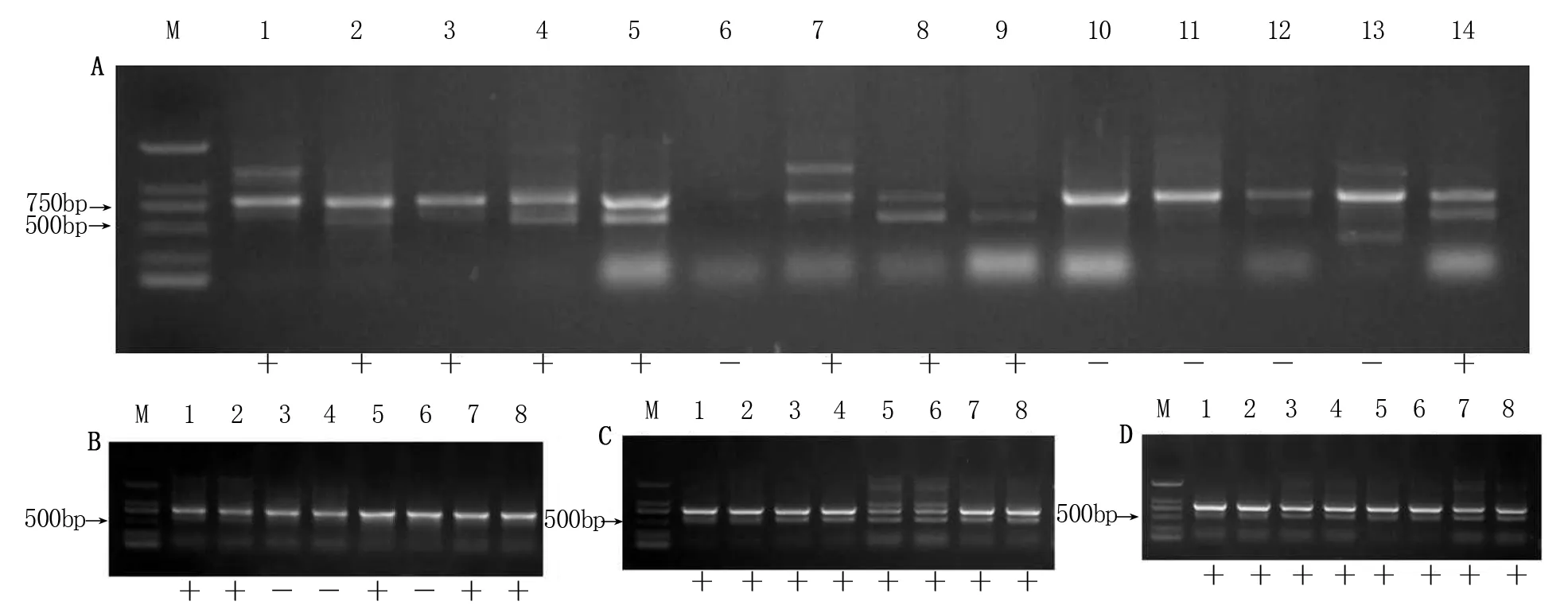
图2 无核探针GSLP1-569对亲本及杂交子代的检测Figure 2 The detection of grape parents and hybrid progeny with seedlessness gene using GSLP1-569 probe
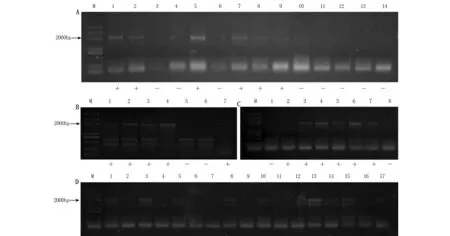
图3 无核分子标记SCF27-2000对亲本及杂交子代的检测Figure 3 The detection of grape parents and hybrid progeny with seedlessness gene using the molecular marker of SCF27-2000
通过胚挽救育种技术成功实现将香味和无核性状的叠加,创制无核玫瑰香味新种质。当亲本基因型不同时,胚挽救杂交组合的成苗率差异较大。研究中‘红宝石无核×阳光玫瑰’杂交组合的胚萌发率和成苗率都最高,与前人研究得到‘红宝石无核’适宜用来配置胚挽救组合结果一致[19]。其原因可能是‘红宝石无核’与其他葡萄亲本的亲和性强,有利于受精胚的发育和生长。此外,‘Fresno Seedless’作为母本时成苗率也较高,且以其作为母本的杂交幼苗长势较其他组合更为健壮,具有生根快的特点,在胚挽救组合的配置中也可作为优良母本供选。不宜选择‘昆香无核’‘美丽无核’‘京早晶’等品种配置胚挽救杂交组合,以上品种作母本的杂交组合成苗率都较低,有可能是由于胚败育发生过早导致胚珠活力低,挽救效果较差。基于前期萜类物质合成关键酶基因的表达分析,筛选香味基因表达较高的品种配置杂交组合,筛选出的材料有:‘阳光玫瑰’‘玫瑰香’‘183’。结合本课题组前期研究结果,还增选‘贵妃玫瑰’‘巨玫瑰’作为胚挽救杂交组合香味父本。在母本相同的情况下,以‘玫瑰香’‘183’作为父本配置的杂交组合成苗率较其他组合更高,因此适合将其作为父本选育香味种质,父本的基因型对胚挽救育种效率也具有影响。
在胚挽救育种技术中,果实的取样时间是重要的影响因素[20],其主要原因是取样时间不同导致杂种胚的发育程度不同[21-22]。研究中的供试母本‘京早晶’是优质早熟无核葡萄品种,在北京地区5月下旬开花,7月下旬果实成熟[23]。研究配置的‘京早晶×玫瑰香’杂交组合是在位于新疆的鄯善进行,在授粉后的36 d和42 d取样都具有较高的胚发育率,42 d时取样胚发育率最高,达到13.33%。可能由于此时胚的发育程度较高,也可能与新疆当地气候有关,授粉后42 d时已为6月末,鄯善当地的日均气温为22~37 ℃,日照时间充足,降水量少,气候干燥,利于杂种胚在果实内生长发育,因此在葡萄生长环境与新疆当地气候相近时,建议在授粉后42 d采样为宜。本研究的另一供试母本‘Fresno Seedless’是种子败育型无核葡萄品种,李志瑛[24]研究得出‘Fresno Seedless’葡萄的胚挽救最佳取样时期是花后32~36 d。研究配置的‘Fresno Seedless×玫瑰香’杂交组合是在陕西杨凌进行授粉作业,在授粉后36 d杂种胚的发育率和成苗率都最高,这与前人研究结果一致[24],建议将授粉后36 d作为‘Fresno Seedless’最佳采样时间。
胚萌发培养基中的激素成分对杂种胚的存活与发育有重要影响[25]。BR是一种广谱高效的植物生长调节剂,可以促进细胞伸长和分裂,在葡萄上也有应用研究[26]。在试验中,选取不同浓度梯度的BR分别与6-BA、IAA组合添加至胚萌发WPM基础培养基中,在同时添加适宜浓度的BR和6-BA时,‘红宝石无核×阳光玫瑰’杂交组合的后代胚萌发率显著高于对照组,推测BR与6-BA在适宜浓度范围内对杂种胚的萌发具有协同作用,以0.1 mg/L BR+0.2 mg/L 6-BA处理效果最好。而在同时添加不同浓度梯度的BR和IAA两种激素时,胚萌发率均低于对照组,可能是由于IAA与BR均为生长素,作用机理相近,重复添加致使胚萌发培养基中生长素浓度过高从而抑制幼胚的生长。在添加了BR的胚挽救杂种胚萌发成苗后,根系均较未添加BR的幼苗健壮发达,长势良好,未出现畸形苗,推测BR还具有促根生长的作用,可在继代培养基中可以添加使用。
研究利用GSLP1-569及SCF27-2000两种无核分子标记对杂交组合的亲本及子代进行检测,在56个杂种子代中进行无核基因分子标记检测,筛选出14个同时携带两种不同无核分子标记的特异条带。检测结果表明,‘Fresno Seedless×玫瑰香’杂交组合后代进行无核基因分子标记检测时,其F1代无核率达到87.5%,说明‘Fresno Seedless’作母本时杂交后代无核率高,可将其作为配置胚挽救杂交组合的优势亲本。
综上,通过研究萜类物质合成途径关键酶基因表达分析筛选出适合无核香味胚挽救育种的父本品种有‘玫瑰香’‘183’‘阳光玫瑰’。利用胚挽救技术创制无核香味葡萄新种质,以‘红宝石无核’‘Fresno Seedless’作母本和以‘玫瑰香’‘183’作父本的杂交组合胚挽救成苗率较高。‘京早晶’及‘Fresno Seedless’为母本的杂交组合最佳采样时间分别为授粉后42 d和36 d。WPM培养基+0.1 mg/L BR+0.2 mg/L 6-BA可以促进胚挽救的幼胚生根。在56个杂种后代中进行无核基因标记检测,筛选出14个杂种后代同时携带不同无核分子标记的特异条带。今后还要对这些杂种植株的无核性状和香味进行田间调查,为无核香味葡萄育种提供新种质。
猜你喜欢
今日农业(2021年19期)2022-01-12
今日农业(2021年10期)2021-11-27
福建基础教育研究(2019年2期)2019-09-10
人文杂志(2017年9期)2018-03-10
现代农业科技(2016年22期)2017-03-24
江苏农业科学(2015年5期)2015-10-20
科技资讯(2014年26期)2014-12-03
小说月刊(2014年11期)2014-04-18
现代农业科技(2009年1期)2009-04-09
现代农业科技(2009年17期)2009-03-08
