
近40 年来对南海化学海洋学研究的新认知
2021-06-03 09:25:40宋金明王启栋
热带海洋学报 2021年3期
宋金明, 王启栋
1. 中国科学院海洋生态与环境科学重点实验室, 中国科学院海洋研究所, 山东 青岛 266071;
2. 青岛海洋科学与技术试点国家实验室海洋生态与环境科学功能实验室, 山东 青岛 266237;
3. 中国科学院大学, 北京 100049;
4. 中国科学院海洋大科学研究中心, 山东 青岛 266071
南海由中国华南大陆、中南半岛、马来半岛和马来群岛所围绕, 周边分布中国、越南、柬埔寨、泰国、马来西亚、新加坡、印度尼西亚、文莱和菲律宾等9 个国家, 其面积约3.56×106km2, 平均水深1212m, 最大水深 5559m。南海地处热带—亚热带,其航运、油气、渔业等海洋资源丰富, 位于北部和西部的珠江、红河、湄公河和昭披耶河4 条重要河流输入南海, 周边国家沿岸区域人口稠密, 经济发展迅速。南海作为我国最大和全球重要的陆架边缘海, 在全球特别是东亚区域社会经济发展中所起的作用巨大, 更直接关系我国的国家安全, 南海海洋学研究在国际海洋学中占据重要地位, 所以, 南海及其海洋学研究也一直是全球关注的热点。
南海的化学海洋学研究与我国近海研究几乎同步, 新中国成立后, 特别是近40 年来, 南海化学海洋学经历了从大面上的广谱调查, 再到持续的定点观测, 发展到近年的系统深入过程研究, 从发现南海环境化学的自然现象和规律, 到探究南海化学过程的变化机理, 产出了大量原创新的论著, 据中国知网的统计, 至今发表的涉及南海的学术论文约有5.89 万篇(占全部海洋学论文86 万篇的6.85%),其中涉及南海博士学位论文994 篇(1990~1999 年间4 篇, 2000 年后990 篇; 占海洋学博士学位论文6887 篇的14.4%)、硕士学位论文4431 篇(占海洋学硕士学位论文3.21 万篇的13.8%, 均为2000 年后)。这其中涉及南海化学的论文309 篇(1999 年以前82 篇, 2000 年及以后227 篇;其中博士学位论文30 篇、硕士学位论文41 篇, 均为2000 年后发表, 从这些数据足以看出南海及其化学海洋学在我国海洋学研究中的重要性。
近40 年来, 南海的化学海洋学研究取得了系统性的新认识, 既包括自然现象和规律的发现, 也包括过程机理机制解析研究的新论点。本文从南海次表层跃层生态系的发现、南海碳源汇研究发展、珠江口低氧脆弱生态区系统演变、南沙珊瑚礁维持高生产力新论点以及南海生态环境历史演变探析等五个方面, 系统地总结并归纳了南海近40 年来化学海洋学研究的重要进展。在此基础上, 分析展望了南海化学海洋学应关注的重点领域, 以期为进一步探明南海的生态环境变化特征, 持续利用南海资源环境奠定基础。
1 南海次表层海水存在着以亚硝酸盐为代表的生态环境参数极值现象, 不同参数出现极值的水层深度范围不同, 由此形成的跃层生态系统与其他生态系统显著不同
20 世纪80—90 年代, 中国科学院南海海洋研究所韩舞鹰研究团队, 通过对南海北部、南沙群岛海域等不同南海区域的多次调查, 系统研究了南海水体理化参数的分布特征(图1), 发现亚硝酸盐等在次表层出现最大值, 提出了“跃层生态系”的概念(林洪瑛 等, 2001)。自20 世纪80 年代初开展的多次调查发现, 在南海的不同区域海水的次表层存在亚硝酸盐的最大值“薄层”, 这些薄层平均厚度一般为20~35m, 处于40~100m 的水层, 其亚硝酸盐浓度平均为0.19~0.23μmol·dm-3。而在薄层上下的水体中, 海水亚硝酸盐含量几乎检测不出。同时, 亚硝酸盐最大值的上层存在叶绿素最大值和光衰减系数最大值, 再向上(海表面方向)则依次是溶解氧最大值和密度跃层。薄层中的亚硝酸盐可能是铵硝化或厌氧铵氧化过程的中间产物, 铵则是来自生物代谢及残体含氮有机物的分解。亚硝酸盐薄层存在的另一个重要原因是浮游生物在跃层下形成的高密度层与下部的水体形成分层, 阻碍了代谢有机物的下沉和向上传递。所以,初级生产水平、生物代谢沉降与分解、铵向硝酸盐的转化等多因素的综合作用决定了亚硝酸盐薄层形成的厚度和所在的深度(韩舞鹰 等, 1991)。
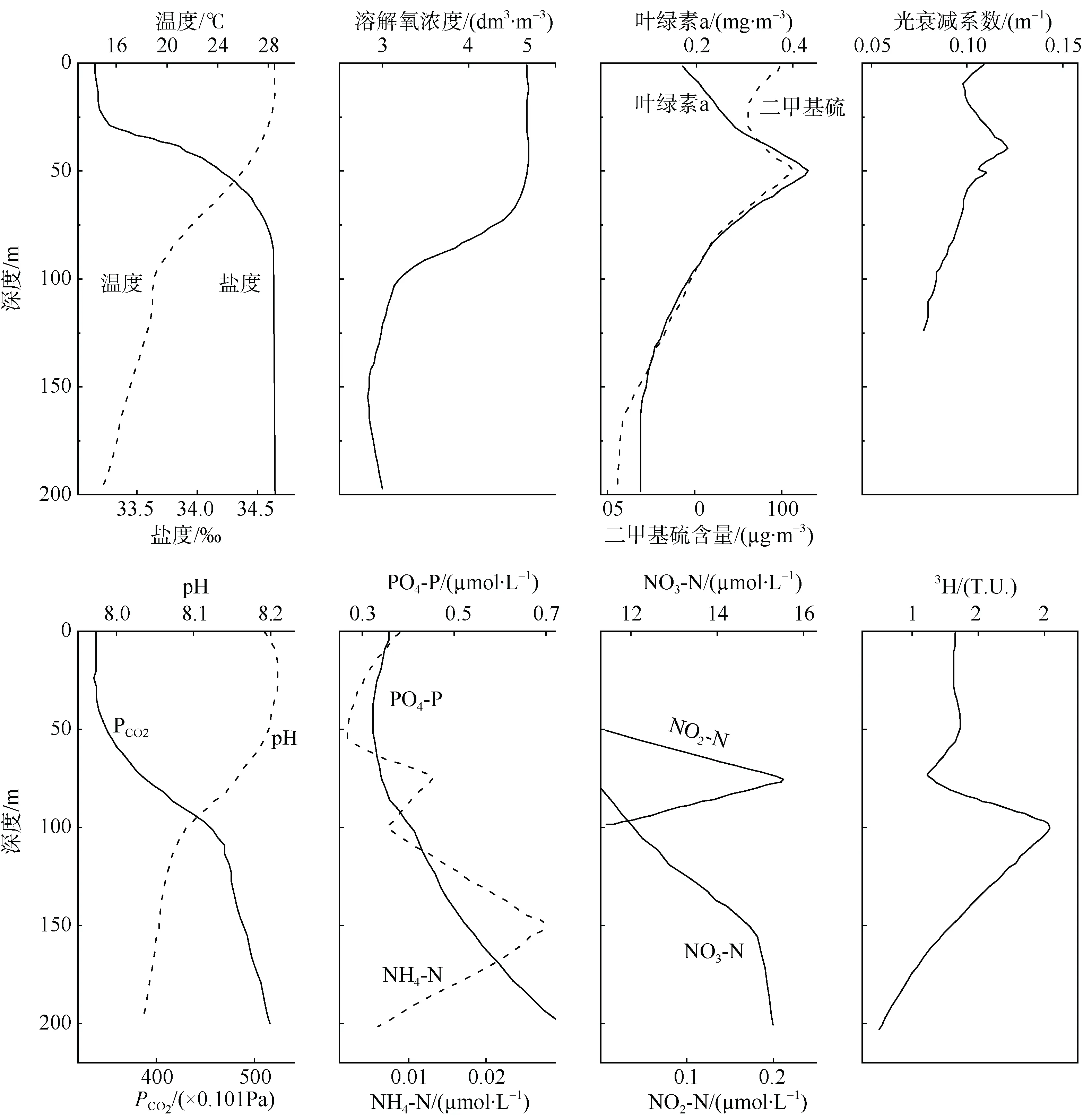
图1 1991 年12 月南沙群岛海域上层水理化参数的极值现象(林洪瑛 等, 2001)Fig. 1 Vertical profiles of environmental parameter extremums in the upper-layer waters of the Nansha Islands during December 1991. After Lin et al (2001)
1985—1999 年间对南沙群岛海域的12 个航次的调查发现, 从海洋表面开始垂直向下20~100m 深度处分别存在亚硝酸盐、溶解氧、氧饱和度、pH、叶绿素a 和光衰减系数的极大值薄层现象, 以及海水二氧化碳分压的极小值。这些极值现象可终年存在, 但由极值的特征值及其与表层含量的差值所反映的极值强度在不同的季节稍有不同。另外,冬季调查的结果显示, 放射性核素氚3H 的含量以及234Th/238U 比值在75~100m 深度存在极大值, 而二甲基硫(dimethyl sulfide, DMS)的含量在20~75m深度存在极大值(图1)(林洪瑛 等, 1989, 2001)。南沙群岛海域次表层海水生物密集和化学参数极值的存在, 表明该海域上层水的生境独特(林洪瑛 等,1989, 2001, 2003)。
上述现象的缘由, 可能是南沙群岛海域表层海水光照强烈且海水温度高, 强光照对浮游植物的生长产生抑制, 再叠加上温跃层常年存在, 阻碍了深层海水对上层海水营养盐的补充。表层海水的营养盐含量很低, 导致上层海水的初级生产水平低下。由于次表层存在内波等中小尺度的动力学作用过程,海水垂直混合较好, 浮游生物可在此处获取相对充足的营养盐, 且该处光照和温度适宜, 因此该水层的浮游植物生长旺盛, 其生物量聚集和叶绿素含量也呈现极大值。悬浮在次表层的大量生物颗粒和其他微粒使光的散射加强, 产生海水消光值的最大值;主要产生于浮游生物细胞的二甲基硫的含量也在此处出现极大值, 这都反映了在此范围的深度层存在浮游生物的密集层(林洪瑛 等, 2001)。浮游植物的光合作用吸收海水中的二氧化碳, 使跃层深度层出现海水二氧化碳分压的极小值和pH 的极大值。在均匀层的上部水体中, 光合作用产生的氧加上海气交换溶解的氧使得上层海水氧饱和度提高; 而在次表层, 海水温跃层和密度跃层阻碍了上层溶解氧的补充, 浮游生物呼吸和大量有机物质的降解消耗了光合作用产生的氧, 使该层海水溶解氧含量降低,因而溶解氧的极大值往往出现在跃层上界深度处。研究显示, 在南沙群岛海域, 上准均匀层中光合作用产生的颗粒有机物质绝大部分输送至次表层,50m 深度的输出效率可达 95%, 在约80m 深度的真光层底部仍可达65% 。因此, 南沙群岛海域海水次表层的“生物泵”作用比其他海域更强烈, 使“贫瘠”的南沙群岛热带海域次表层形成一片“绿洲”。基于这些现象的发现, 林洪瑛等(2001)指出南海次表层海水跃层极值现象的本质是存在“跃层生态系”, 继而提出了“跃层生态系”的概念。
南海次表层海水存在生态环境参数极值现象是近40 年来南海化学海洋学研究的重要发现, 对研究河口-陆架边缘海-深海盆复杂构造海域的海洋学过程具有重要的启示作用。
2 南海碳循环过程十分复杂, 动力作用下的生物泵过程以及碳源汇区域和季节变化巨大, 在不同的区域不同的时间南海碳源汇的性质和强度迥异, 全年尺度上总体表现为大气二氧化碳的弱源
南海碳循环研究是我国海洋碳循环研究的重要组成部分, 近40 年来国内外多个研究团队从碳通量、碳源汇及控制因素、碳循环的关键过程等角度开展了大量研究(Song, 2010), 与渤黄东海碳循环研究一起构成了我国近海碳循环研究的主框架(宋金明 等, 2018; Song et al, 2018)。韩舞鹰团队对南海碳循环的早期研究主要基于观测数据的南海碳循环箱式模型, 其研究结果表明, 南海碳的收支其输入通量从大到小分别为中层水、底层水、河流和雨水, 输出南海的碳通量则为上层水、沉积埋藏和海气界面交换。其中, 99%的碳输入主要通过中层水和底层水进入南海, 然后随上升流进入上层水体, 而河水和雨水输入的1%的碳则直接进入上层; 这些输入到南海上层水体的碳, 约有91%从南海流出至邻近大洋区域,少部分以颗粒态的形式垂直输送至下层。而垂直向下输送的颗粒态碳分别有93%和0.6%在南海中层和底部被分解, 进入再循环并返回上层, 最终约有 6%沉降至海底, 被沉积物埋藏(韩舞鹰,1998)。南沙海域真光层内海水颗粒有机碳生成速率为298mg·m-2·d-1, 其中约三分之一在真光层内分解参与再循环, 剩下近三分之二离开真光层向下输送;总溶解无机碳穿过跃层垂直向上输送通量为3600mg·m-2·d-1, 而真光层颗粒无机碳向下输送通量为27mg·m-2·d-1; 总的来看, 大气二氧化碳经海气界面进入海洋的净通量为13mg·m-2·d-1(韩舞鹰 等,1994)。
在南海碳源汇研究方面, 前期研究总体认为南海是大气二氧化碳的碳汇, 但后来的研究大多认为南海是大气二氧化碳的弱源, 南海北部的珠江口及其邻近海域由于初级生产力较高, 可吸收大气CO2,体现为大气的较强碳汇。而随着离岸距离不断增加,南海外陆架海域在“高温-寡营养”的影响下转而成为大气CO2的源(焦念志 等, 2018)。从总体而言, 夏季南海北部海域是大气 CO2的一个显著的源, 如在台湾海峡南部海域海水向大气释放CO2的通量以碳计为 5.28~37.11mg·m-2·d-1, 其释放通量在不同的时间和空间上也有差异。2002 年7 月10 日的调查结果显示, 台湾海峡南部海水向大气释放CO2通量以碳计为5.28~16.58mg·m-2·d-1。如将南海分为受珠江冲淡水影响的北部陆架、南海北部陆坡及海盆、南海中南部海盆和吕宋海峡西侧海域四个区域,可以发现, 北部陆架海域PCO2较低且季节变化不明显(320~390μatm), 南海北部陆坡和海盆区则表现出强烈的季节性变化特征, 即呈现气温高时表层海水PCO2高, 气温低时表层海水PCO2低, 南海中南部海盆区域海水全年PCO2相对较高(360~425μatm), 而在吕宋海峡西侧海域, 冬季在涡旋/上升流偶发事件的影响下海水向大气净释放CO2, 四个区域面积加权后其CO2平均通量以碳计为13.21± 20.42mg·m-2·a-1,每年释放(1.80±2.76)×1013gC 的CO2, 如外推至除北部湾和泰国湾外的整个南海海域, 释放通量则为(3.36±5.13)×1014gC·a-1(刘茜 等, 2018)。这些数据表明, 南海碳源汇性质和强度具有明显的区域性和时间变化性, 陆架边缘海特征十分明显。许多研究者还对碳循环过程进行了剖析, 譬如发现南海热带气旋发生前后, 底层富含CO2的水体上涌, 对南海从碳汇转而变成碳源有重要影响; 碳通量的大小与季风强弱有关, 外源输入的碳仅占初级生产碳的1.4%~1.6%; 硝酸盐浓度较高的水域, 碳生产的输出较低, 输出碳的生产与初级生产呈正相关关系。
对多年调查结果进行总结分析, 可以发现南海海盆是大气CO2的弱源区, 海洋向大气释放的碳通量以碳计为25.22±3.60mg·m-2·d-1, 南海北部陆架海域是碳汇区, 海水吸收大气的碳以碳计为-26.42±42.04mg·m-2·d-1, 南海总体上每年向大气释放的碳约为(1.33±1.88)×1012g(图2)。南海海盆和北部陆架分别为大洋主控型的边缘海和河流输入主控型陆架海, 分别接受大洋和河流输入的外源无机碳和营养盐, 经由一系列动力过程进入真光层后被生物消耗, 浮游生物对无机碳和营养盐利用差异以及它们来源/除去途径的不同最终决定了CO2的源汇格局。在南海海盆, 海水无机碳相对过剩, 部分以CO2形式向大气释放, 即为源; 而在南海北部陆架,无机碳相对缺乏, 海水需从大气补充CO2, 即为汇(戴民汉 等, 2020)。
综上所述, 由于南海区域性差异太大, 在不同时间和不同区域, 碳源汇的性质和强度会有巨大的差异(Song, 2010), 现有的结论也是基于有限调查数据的扩展评估。深层次了解南海的碳源汇特征, 揭示南海碳循环的控制过程还有大量的工作要做。
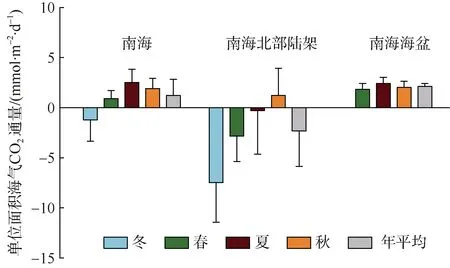
图2 南海海气界面碳通量(戴民汉 等, 2020)Fig. 2 CO2 fluxes across the water-air interface in the South China Sea. After Dai et al (2020)
3 南海北部的珠江口邻近海域和深海盆的生态环境特征与化学物质循环和陆架边缘海、珊瑚礁等密切相关, 但又与陆架边缘海、珊瑚礁等显著不同, 珠江口底层海水明显低氧, 珠江口海域是一个以低氧为特征生态环境脆弱区
从1980 年代至今, 调查表明, 珠江口海水存在持久性缺氧区, 即一年四季持续低氧, 但低氧强度的季节性变化很明显。2000—2001 年的调查结果表明, 水质恶化导致珠江口下游出现明显的季节性低氧现象, 其中河口下游至香港外侧区域低氧最为严重, 缺氧主要出现在6 月至9 月, 而长期监测数据也显示珠江口下游水体低氧正在逐年加剧(图3), 其底层水溶解氧(dissolved oxygen, DO)每年每升约下降0.06mg, 在虎门到广州后航道存在严重的表层低氧区, DO 最低达0.48mg·L-1以下, 其生物群落明显受到低氧影响。
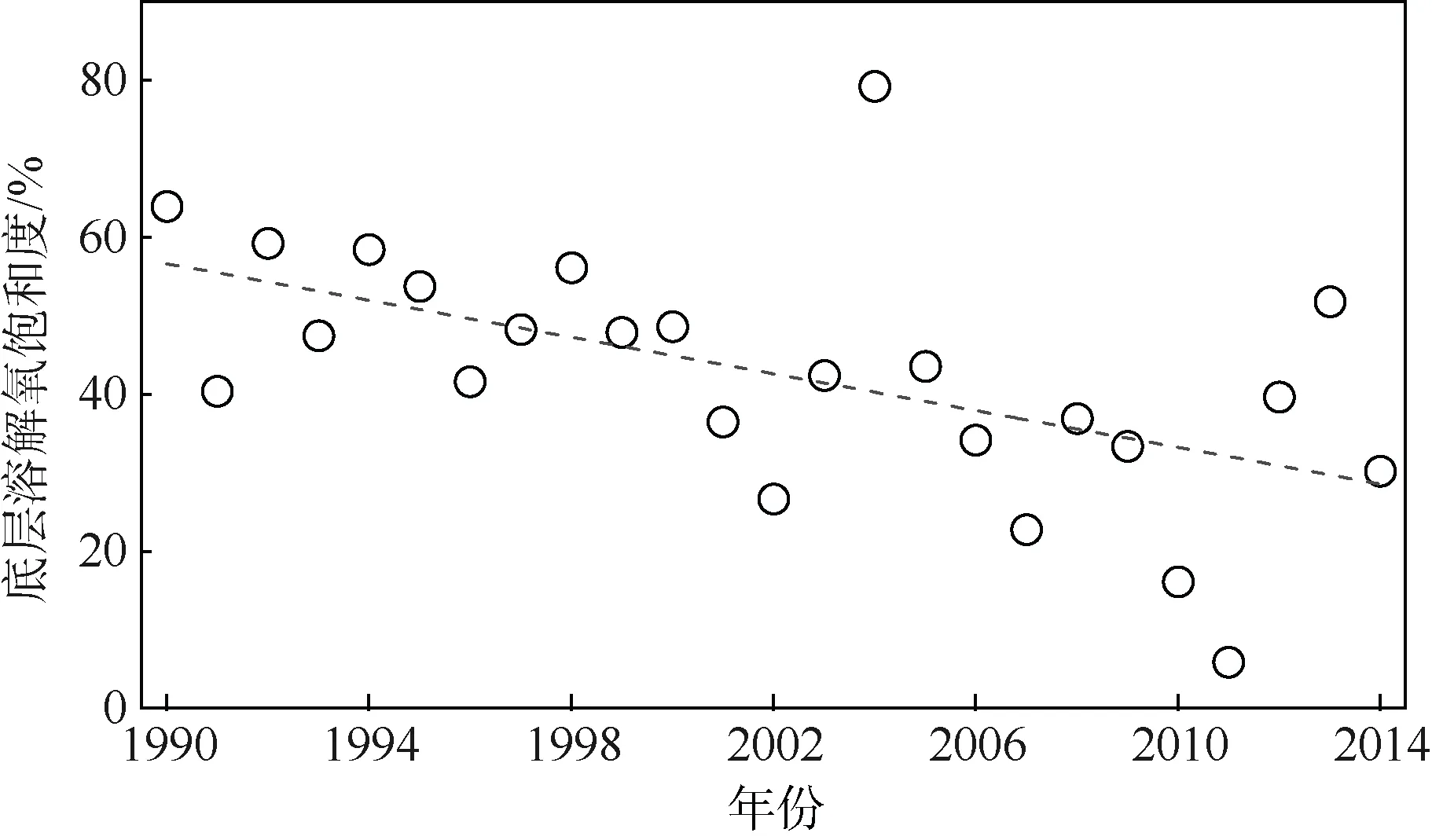
图3 珠江口(22°9'12.7″N, 3°57 '43.6″E, 深度21m)O2饱和度的变化(Qian et al, 2018)Fig. 3 Temporal variation of bottom-water dissolved oxygen saturation (22°9'12.7″N, 3°57 '43.6″E, 21 m depth).After Qian et al (2018)
2011 年春季, 珠江口上游出现显著低氧区, 范围从东莞江入海口往北至东江干流入海口, 多数站位海水中DO 含量低于 3.5mg·L-1, 且表、底层水体中溶解氧含量相差不大, 表层低氧面积略大于底层。这些结果表明此时该区域水体垂向混合充分, 层化不明显, 对低氧形成的影响不大, 因此径流排放的有机污染物降解耗氧可能是低氧形成的主要原因, 而沿着径流方向, 有机污染物受到海水的稀释作用而逐渐降低, 耗氧能力随之降低, DO含量则在海水带来的新鲜氧气输入和化学需氧量(chemical oxygen demand, COD)降低的双重作用下而逐渐升高(李秀芹 等, 2014)。也有研究认为, 在珠江口伶仃洋特别是万山群岛附近海域, 由温盐差造成的水体层化是夏季底层DO 偏低的主要原因(叶丰 等, 2012)。以珠江口为代表的南海北部海域水体生态环境与陆架生态系统有相似之处, 生态环境参数明显受控于陆地物质输入、上升流混合、近岸海洋工程与海域利用(包括海水养殖等)等因素影响(龙爱民 等, 2006), 同时这些因素又与珠江口特异的地形地貌相耦合, 影响了珠江口海域低氧的形成及规模。
研究表明, 大量有机物和铵的输入是造成珠江口邻近海域强烈局部缺氧的重要诱因, 但就综合而言, 近海低氧形成的根本原因正如作者一直认为的是由于海洋区域地形地貌的“凹槽”、“凹沟”或“被水坡”的“静水作用”所致, 这种特殊的地形地貌使水体交换受到阻碍, 水体分层, 有机物沉降分解以及陆源污染物输入等, 加重了在这种区域水体缺氧的程度或范围(宋金明, 1997, 2004; 宋金明等, 2020)。正如珠江口海域的滩多、槽(水道)多、岛屿多, 内伶仃洋水下地形呈现三滩(西滩、中滩和东滩)两槽(西槽和东槽), 许多小地貌类型如槽沟、沙波、洼地和浅滩等在多处出现, 该区域位于矾石水道以北最大的洼地, 面积约为4.34km2, 有的洼地很小, 面积仅有0.05km2, 珠江水下地形变化大。伶仃洋地形地貌 “三滩二槽”的演变可分为两大阶段的5 个演变时期, 包括自然冲淤阶段的2 个时期:(1)形成期, 至1936 年, 中槽淤浅成为现在的中滩,初步形成“三滩二槽”的格局; (2)稳定期, 西滩不断淤积, 深槽不断加深, 至1984 年, 显著的“三滩二槽”形成并稳定存在, 总体呈现“西淤东冲”地貌变化趋势。人为活动干扰阶段的3 个时期: (3)中滩淤积加剧期, 1984—1997 年期间, 由于上游采沙导致分流比变化, 中滩淤积加剧; (4)中滩蚀淤过渡期,1997—2004 年期间, 泥沙来源大幅减少, 中滩由淤积状态变为有冲有淤的不稳定状态; (5)中滩冲刷加剧期, 2004 年至今, 西部口门及水道则为冲刷地貌,伶仃航道和东槽因海砂开采和疏浚, 显著下蚀(李团结, 2017)。
由此可以推知, 珠江口海域的低氧尽管一直存在, 但其分布的范围、强度等在不同季节、年代是不同的(Qian et al, 2018)。
深入的过程研究对揭示南海北部生态环境变化机制至关重要(Song, 2010)。有机质降解和硝化作用是造成溶解氧含量下降的重要过程, 而稳定碳、氮同位素可用于评估其对溶解氧消耗的相对贡献。相关研究结果显示, 陆地和海洋来源的有机碳均是水体有氧呼吸的主要碳源, 分别可占呼吸耗氧的38%~60%和40%~62%; 铵盐的硝化作用是导致溶解氧降低的另一重要过程, 根据铵盐浓度、氮同位素及实际耗氧过程的化学计量比, 可以估算出硝化作用在夏季珠江口上游和中游分别贡献34%~64%和17%~32%的溶解氧消耗, 而冬季硝化作用对上游溶解氧消耗的贡献可达 60%(叶丰 等,2019)。
南海海盆在800~1000m 出现溶解氧极低值层(oxygen minimum zone, OMZ), 在1000m 以深DO 又随深度的增大而略微升高。南沙深海盆具有明显的深海大洋特征, 又与高生产力的珊瑚礁生态系密切关联, 上层水体生态环境与南海北部陆架区域相似,呈现次表层的“跃层生态系”特征, 下层水具有明显的大洋属性, 且下层水溶解氧含量较高(宋金明,2004)。
4 南沙珊瑚礁生态系统物质循环快速, 生物过程控制着化学物质的垂直转移, 提出了维持珊瑚礁生态系统高生产力的新机制-“拟流网理论”
在中国陆架边缘海中, 南海具有渤海、黄海和东海所没有的珊瑚礁生态系统, 这是南海海洋科学研究中难得的探索领域。近40 年来, 研究者在南沙珊瑚礁系统生物海洋学、地质构造与沉积等领域发表了大量学术论文和著作, 在我国海洋科学研究进程中占有重要一席。南沙珊瑚礁生态系统地处相对寡营养的热带海域, 但其生产力却很高, 如同热带海洋荒漠中的点片状绿洲。我国海洋科学工作者聚焦这一领域进行了系统研究, 发现和提出了一些新的论点。
早期的研究认为, 珊瑚礁是一个典型的自养生态系统, 毋需同外围进行营养物质交换, 但后来的研究发现, 珊瑚礁生态系统可向外围贫瘠海域输送少量营养盐。因此, 从物质守恒和维持珊瑚礁生产力的角度看, 珊瑚礁内必定有外源的营养盐输入(宋金明, 1999a)。随着对珊瑚礁生态系统研究的不断深入, 科学家们从不同的学科角度提出了维持珊瑚礁营养盐供给与高生产力的4 种主要假说和模型。具体假说和模型如下。
1) 珊瑚礁内部营养盐高效循环模型, 认为珊瑚礁内的珊瑚和石灰藻类等生产者对营养环境高度适应, 凭借内部环境中(礁体和邻近沉积环境)保守而高效的营养物质循环, 珊瑚礁生态系统可维持较高的总生产力, 但其净生产力却很低;
2) 珊瑚礁边缘上升流与地热-内上升流模型,珊瑚礁及其沉积环境明显受海洋和大气物理过程控制, 海流受礁缘地形限制而产生的上升流可能是珊瑚礁营养盐的主要来源;
3) 珊瑚礁水动力与停留时间模型, 认为水流速度能够促进生物对营养盐的吸收, 实验发现海水流速的加大与珊瑚礁内生物吸收营养盐的量成正比,证实珊瑚礁内的生物可通过潟湖内、外海水的运动可以补充相当量的营养组分;
4) 珊瑚礁内外动力作用下的拟流网模型,认为热带海洋中的珊瑚礁如同流网, 其中相对固着的生物可以高效地富集、利用寡营养海流中的营养物质, 再加上珊瑚潟湖内部营养盐可快速、高效地循环, 共同维持了珊瑚礁生态系统的高生产力。
据拟流网模型, 营养盐的原位再生多发生于珊瑚上部间隙水中和沉积物底部, 珊瑚礁特有的高孔隙率(20%~35%)、高渗透性特征为细菌降解、有机质矿化和营养物质再生提供了一个巨大的流动的水库, 大的礁洞中的营养物质易被水流带走,而大量的小孔隙则为控制营养物质流失发挥了巨大的作用, 固着的珊瑚礁体像流网一样有效地从不断流动的海水中汲取营养物质, 而且吸收量随海水流速的增加而增加, 在寡营养水体中起到了营养物质“富集器”的作用(宋金明, 1999a, 2004;Song, 2010)。同时, 当相对固着的生物富集了寡营养海水中的营养物质后, 其旺盛的生命活动和潟湖内的特殊环境使得营养物质快速再生, 营养物质的循环速度可达外海的几十倍至数百倍, 这对维持珊瑚礁生态系统的高生产力至关重要(宋金明, 1999b, 2004)。在南沙渚碧礁的潟湖内, 两个站位产生的颗粒有机碳(particulate organic carbon,POC)分别有93.55%和95.83%在水体中即被矿化分解, 沉降至沉积物中的POC 很少, 其中生物捕食和细菌腐解作用可将99%左右的生物碎屑POC转变为无机碳, 重新进入循环; 潟湖内颗粒总氮(particulate total nitrogen, PTN)和颗粒有机氮(particulate organic nitrogen, PON)释放率分别超过 90% 和 86%, 颗 粒 总 磷(particulate total phosphorus, PTP)释放率为58.7%、85.2%, 颗粒有机磷(particulate organic phosphorus, POP)的释放率在90%以上(宋金明, 2004)。
对南沙珊瑚礁系统营养盐利用的研究表明, 珊瑚的虫黄藻可“奢侈消费营养盐”, 即当大量营养盐在短期内输入到珊瑚礁区时, 以珊瑚及藻类为主的珊瑚礁生物可以快速地过量吸收营养盐, 从而迅速降低海水营养盐水平。这不仅可避免珊瑚礁长期处于富营养化状态, 也在客观上固定了宝贵的营养源。这种营养盐调节机制很像沙漠中骆驼可一次性补充大量水, 而后可维持多天的无水补充状态。珊瑚礁的这种“奢侈消费营养盐”的机制对维持珊瑚礁的正常运转很重要(赵卫东 等, 2001)。这就是我国研究者提出的“拟流网理论”(宋金明, 1999a, b; 2004;Song, 2010)的核心——“潟湖内快速循环—奢侈消费营养盐”。换言之, 尽管珊瑚礁外的海水寡营养,但随海水流动, 寡量的营养盐进入固着的珊瑚礁就如同水流中的鱼虾进入流网, 被固着的虫黄藻等奢侈吸收下来; 营养盐继而在珊瑚礁内进行快速循环,维持珊瑚礁的高生产力。
综合的研究表明, 珊瑚礁生态系统的营养过程并不唯一, 其控制因素与珊瑚礁的发育环境有关。珊瑚礁形成的地质构造是其前提, 物理因素(如温度、盐度、波浪、光照、潮位和大气过程等)是关键的制约性因素。珊瑚礁在周围寡营养的海水环境中发育所需要的营养盐, 主要靠珊瑚礁自身发展的多种营养调节机制来维持, 这些调节机制与珊瑚礁生长的物理环境协同作用, 维持了其强大的生产力和丰富的生物多样性。但在诸多因素中,哪些因素起主导作用, 不同区域的珊瑚礁生态系统可能都不相同。
5 对南海沉积物化学的系统研究发现, 沉积物-水体化学物质循环存在密切的耦合作用; 南海珊瑚礁或沉积岩心化学物质分布变化可反演其历史变化, 南海冰期表层海水古生产力高, 约为间冰期的1.6倍, 晚中新世南海南部发生了一次“生物勃发事件”, 其生产力主要受季风和陆源营养物质输入量影响, 东北季风与西南季风在不同区域其影响程度不同
古海洋研究可了解过去海洋的变化, 更为重要的是可为预测未来变化奠定基础(Song et al, 2020)。自2000 年以来, 南海的古环境变化研究有诸多的进展。
沉积物研究是探究南海古环境演变的基础, 南海表层沉积物有9 种类型, 呈条带状分布, 与等深线分布基本一致, 沉积物的类型与沉积物化学组成有十分密切的关系, 在南海北部陆架区, 其表层沉积物由现代沉积、再造沉积、残留沉积和少量的残余沉积组成, 南部陆架区地处热带, 生物繁茂, 珊瑚礁发育良好, 生物沉积较为普遍, 半深海-深海区生物碎屑含量高。研究发现, 1989—2004 年间南海北部陆架海域水体溶解无机氮含量明显上升, 增幅为每年每升0.38~0.81μmol, 显著高于同期无机磷的增量, 水体氮磷比从10 增加至16 左右(Ning et al,2009)。这些近岸水体的高氮随动力输运至开阔海域,对南海深部产生影响, 如在南沙深海盆海域, 沉积物的埋葬效率只有0.63%, 因此表层沉积物几乎全部的氮均可参与再循环, 沉积物氮与叶绿素和初级生产力分布呈一定的正相关, 由陆源输入及N2的固定提供的氮均很低, 但由沉积物提供的氮源对新生产力的贡献为46%, 很大程度上补偿了浮游植物对水体中营养盐的消耗, 对维持该海域的初级生产起到重要作用(郑国侠 等, 2006)。
在历史不同时期, 南海的不同区域均表明其古生产力有巨大变化, 即冰期较高, 间冰期较低。在对南 沙 南 部 海 域 重 力 柱 状 样 No.17962 (7°11′N,112°5′E)的研究中, 该处近30 万年来的末次冰期和全新世的输出生产力比较分析结果表明, 冰期古生产力明显更高, 约为全新世的 1.6 倍(房殿勇 等,1998) 。 南 海 南 北 陆 坡 两 个 站 位(06°09′30″N,112°12′48″E; 20°07′N, 117°23′E)的柱状样中底栖有孔虫研究表明, 南海近4 万年来, 末次冰期南海冬季风增强, 在南部陆坡产生上升流, 加上低海平面时陆源营养物输入增加, 使得该海区的表层古生产力急剧增大; 末次盛冰期结束后, 南海冬季风明显减弱, 夏季风增强, 并于全新世早期约10000a BP达到最强, 这不仅可能在北部陆坡产生上升流, 而且其大量的夏季风降雨导致河流输入的营养物增多,使南海北部的表层古生产力显著提高(翦知湣 等,1999)。
南海南部 ODP1143 站12Ma 以来的表层生产力变化研究表明, 在 12.3—5.7Ma 期间, 表层生产力较高, 之后开始降低; 在 5.7—0.7Ma 期间, 表层生产力降到最低; 自0.7Ma 以来, 表层生产力又明显升高。晚中新世生物生产力的分析证实, 印度洋-太平洋的“生物勃发事件”在南海同样发生过。南海南部的晚中新世“生物勃发事件”可能是由于较强的上升流导致了较高的生物生产力, 使得该站位的蛋白石堆积速率、碳酸盐堆积速率和总堆积速率都表现出较高的水平(李建如 等, 2002)。
6 南海化学海洋学研究展望
近40 年的南海化学海洋学研究对南海及其邻近环境的化学组成、分布变化、控制因素和过程解析有了比较系统的了解。但由于南海既有陆架边缘海的性质, 也有深海大洋特征, 如陆架边缘特别是南海北部受到大河流的输入影响, 南部南沙海域又有大量珊瑚礁分布, 因此南海生态环境异常复杂,许多海洋学过程尚不清楚, 如南海动力过程和边缘陆地影响究竟对其生态环境的控制分别有多重要?南海与邻近太平洋的相互作用的过程究竟有何变化规律?南海资源环境利用对其生态环境的影响过程如何?等等, 了解这些复杂的过程需要多种有效的化学海洋学手段, 从这个角度来说, 南海的化学海洋研究还任重而道远。
近年来, 健康海洋作为海洋资源环境可持续利用, 支撑社会经济高质量发展的基础, 得到了前所未有的重视。对南海而言, 探明南海海岸带以及近海生态环境演变的过程与规律, 预测其发生的变化对健康海洋至关重要。需重点开展的研究包括: 南海特别是南海北部环境污染管控与水环境安全; 南海资源开发过程中生态环境变化趋势与预测; 南海健康海洋过程研究与评估体系构建; 南海生态系统、生物地球化学循环对全球变化的响应及可持续发展; 南海陆地-近海-大洋-大气耦合系统的运作与发展趋势以及南海海岸带/近海的蓝碳潜力及其对人类活动和气候变化的响应等。
猜你喜欢
海洋学研究(2023年2期)2023-07-27 13:44:32
沉积学报(2023年1期)2023-04-29 18:57:49
海洋通报(2022年2期)2022-06-30 06:07:04
清华大学学报(哲学社会科学版)(2022年2期)2022-03-26 09:25:34
家庭影院技术(2021年5期)2021-07-21 07:15:36
海洋通报(2020年5期)2021-01-14 09:26:58
百科探秘·海底世界(2020年8期)2020-07-29 08:57:46
时代英语·高一(2019年1期)2019-03-13 10:29:48
时代英语·高二(2017年4期)2017-08-11 18:04:36
中国科技博览(2016年25期)2016-12-20 18:28:41
