
公路源重金属对青藏高原高寒草甸土壤线虫群落的影响
2021-06-03 04:27:58薛会英
生态学报 2021年9期
侯 磊,薛会英
1 西藏农牧学院资源与环境学院,林芝 860000 2 西藏高原森林生态教育部重点实验室,林芝 860000 3 东北林业大学生态研究中心,哈尔滨 150040
土壤线虫是土壤生物中的一类数量庞大、种类丰富,并且广泛分布于地球上所有土壤中的土壤动物[1-2],占据碎屑食物网的多个营养级[3],在土壤有机质分解及土壤养分循环方面有着重要作用[4],是土壤指示生物的典型代表[5]。高寒草甸是青藏高原最为主要的生态系统类型之一,极为脆弱,对人为干扰十分敏感[6]。西藏公路迅速发展[7-8],穿越各类脆弱生态系统,其所产生的重金属粉尘等[9-10]对距公路一定范围内的土壤产生影响[11],成为影响公路两侧高寒草甸的潜在环境因子,主要公路源重金属污染物包括铜(Cu)、锌(Zn)、铅(Pb)、镉(Cd)等[12]。
目前在重金属对土壤线虫的影响方面做了一些研究工作,如王赢利等[13]的研究认为重金属污染会降低稻田线虫数量,同时改变土壤线虫群落中的r对策和K对策线虫的比例;李钰飞等[14]通过设置重金属胁迫梯度研究林地和农田土壤线虫群落特征,发现不同生态类型土壤线虫多样性特征随重金属铜、锌浓度的变化规律不一致,较复杂的生态类型中线虫群落对重金属的抵抗力较强。此类研究一般采取的是人为控制试验来探讨土壤线虫群落变化特征,且多使用显微镜对线虫进行形态学分离鉴定,耗时费力[15]。高通量测序技术的发展为环境微生物群落研究提供了便利工具16],被认为有较好的结果一致性[17],也在不同地区土壤线虫多样性研究方面[18-20]得到了一些应用,但在青藏高原土壤线虫研究方面应用还较少[21-22]。
本文利用Illumina Miseq测序技术,在97%相似水平下对所得序列进行生物信息分析,研究青藏高原高寒草甸公路旁的土壤线虫群落特征,为掌握高寒草甸土壤线虫群落对重金属的响应,进一步了解人为干扰下的高寒草甸土壤线虫群落特征提供参考。
1 材料与方法
1.1 试验地概况
邦杰塘草原属西藏林芝市工布江达县,远离城市,人口极少。位于尼洋河上游,米拉山东侧,海拔平均约4450 m,为高寒草甸。年降水量550 mm,年均气温-3.8℃,年均风速3 m/s。主要植物有高山嵩草(Kobresiapygmaea)、紫花针茅(Stipapurpurea)、高山点地梅(Androsacegmelinii)、鹅绒委陵菜(Potentillaansrina)、马先蒿(Pedicularisreaupinanta)、独一味(Lamiophlomisrotate)、肉果草(Lanceatibetica)等[23]。邦杰塘草原海拔落差小,人为干扰少,主要受自由放牧影响,拉林高等级公路贯穿其中,采样点选择在公路旁的地势平坦区域,海拔4339 m,坐标92°26′19.9″E,29°54′28.27″N。
1.2 土样采集和土壤重金属测定
于2019年8月,在离路基0、5、15、30、50、75 m处布设与公路平行的采样线,每条采样线每隔15 m左右布设1个采样单元,共设3个。各样线分别编号为P1、P2、P3、P4、P5、P6,表示在距路基不同距离上的样本。使用土钻取土样,装入塑封袋后储存在冷藏箱中尽快带回实验室。自然风干土样,去除碎石、根系和各种残渣。一部分碾碎,过1 mm筛。将粒径1 mm的土壤充分混合,多点取样5 g左右,用研钵研细,过孔径为-0.15 mm筛,采用“硝酸-氢氟酸-高氯酸”消解体系进行消解,采用火焰原子吸收分光光度法(F-AAS)测定全量 Cu、Zn、Pb、Cd。另一部分土样用以测定土壤线虫群落特征。
1.3 土壤线虫群落的高通量测序
1.3.1DNA抽提和PCR扩增
根据E.Z.N.A.®soil试剂盒(Omega Bio-tek,Norcross,GA,U.S.)说明书进行总DNA抽提,DNA浓度和纯度利用NanoDrop2000进行检测,利用1%琼脂糖凝胶电泳检测DNA抽取质量。采用NF15′-GGTGGTGCATGGCCGTTCTTAGTT- 3′和18Sr2bR 5′-TACAAAGGGCAGGGACGTAAT- 3′引物进行PCR 扩增[16,24]。
1.3.2Illumina Miseq测序
利用上海美吉生物医药科技有限公司的Miseq PE300平台进行测序。
1.4 数据分析
利用上海美吉生物医药科技有限公司云平台(www.i-sanger.com)分析土壤线虫群落及其与土壤重金属之间的关系。使用云平台mothur软件,基于OTU水平进行多样性指数分析,包括Shannon指数、Ace指数和Chao指数。为保证分析在同一水平上进行,分析前进行抽平操作。采用one-way ANOVA进行线虫数据的统计学检验。测序数据比对数据库是NCBI(NT)。RDA分析利用R语言vegan包,用于探讨土壤线虫群落与重金属的相关性。采用环境因子排序线性回归分析重金属因子对土壤线虫群落多样性变异的解释比例。利用SPSS 20.0软件分析土壤化学性质数据差异性。
2 结果与分析
2.1 土壤化学性质
各样本土壤化学性质如表1。可见Cu、Pb、Zn、Cd四种重金属最小值均出现在最远的P6,且P6的Cu、Pb、Cd与P1—P5差异显著(P<0.05),Zn与P1、P2差异显著(P<0.05),但与P3、P4、P5差异不显著。磷、pH、氮、有效磷在P6也呈现出与其他样本的差异。
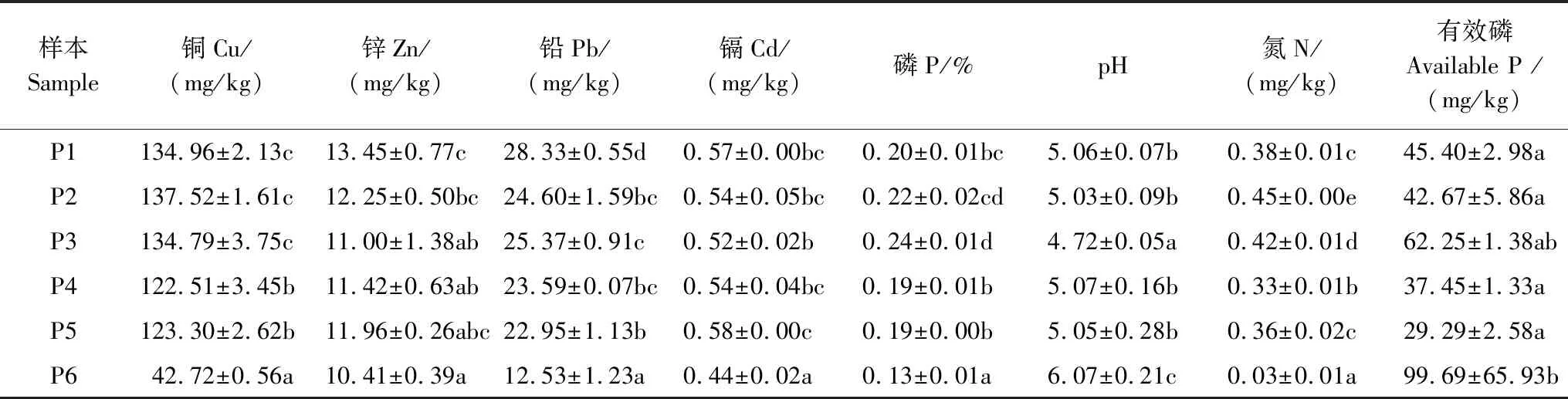
表1 土壤化学性质
2.2 OTU组成
利用物种丰富度指数sobs制作稀释曲线。如图1所示,各样本的sobs稀释曲线趋于平缓,数据量增加后对OTU无影响,说明所得序列基本可以反映实际环境的土壤线虫群落结构。

图1 土壤线虫测序的稀释曲线Fig.1 Rarefaction curves of soil nematodes communitiesP1—P6分别指离公路0、5、15、30、50、75 m处土样
不同样本土壤线虫群落共同拥有的OTU有10个,如图2,P1独有11个OTU,P2独有2个OTU,P3独有4个OTU,P4独有3个OTU,P5独有2个OTU,P6独有9个OTU。
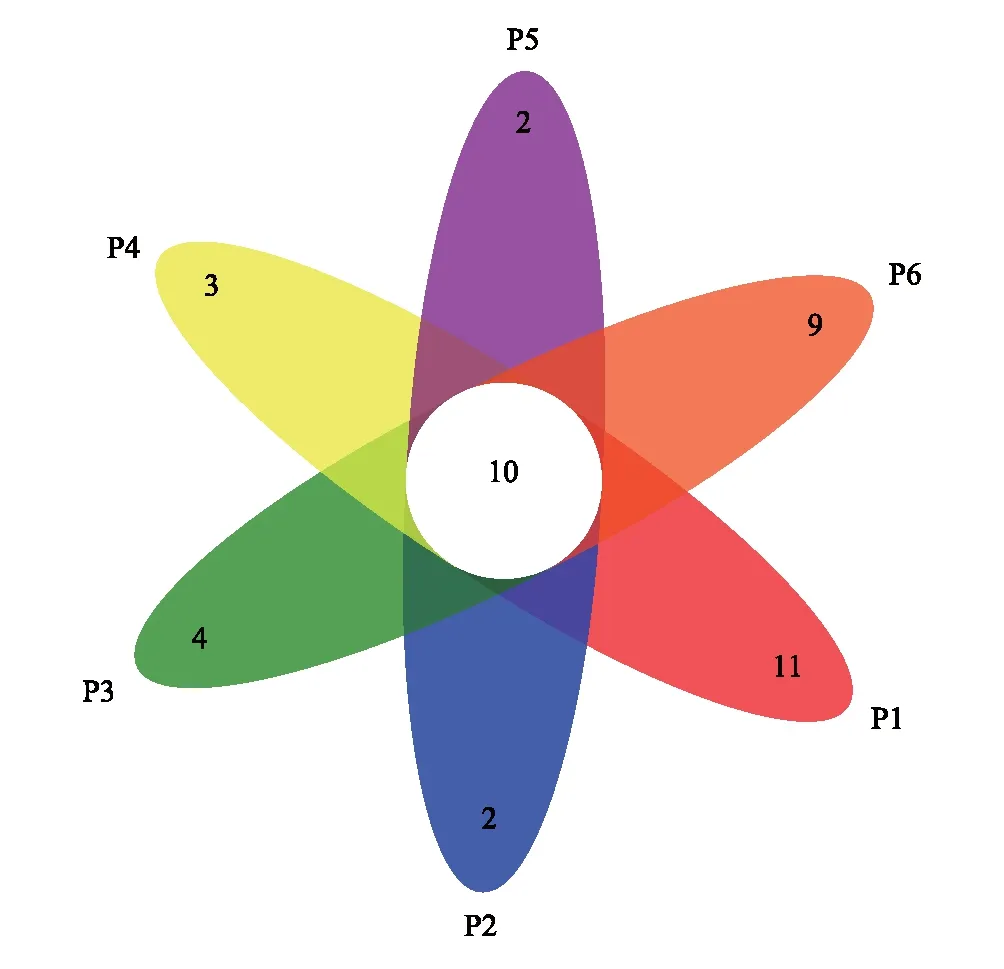
图2 土壤线虫群落OTU维恩图Fig.2 OTU Venn diagram of soil nematodes communitiesOTU:操作分类单元 Operational taxonomic unit
2.3 α多样性
各样本的α多样性指标如表2。不同样本土壤线虫群落多样性各指数略有不同,但最小值均出现在P6。其中P6的Shannon指数与P1、P2、P3、P4差异均显著(P<0.05),与P5差异不显著;P6的Ace指数与P1、P2、P3、P4、P5差异均显著(P<0.05);P6的Chao指数与P1、P2、P3、P4、P5差异均显著(P<0.05)。以上表明距离公路最远的P6土壤线虫群落具有与P1、P2、P3、P4、P5不同的线虫多样性特点。
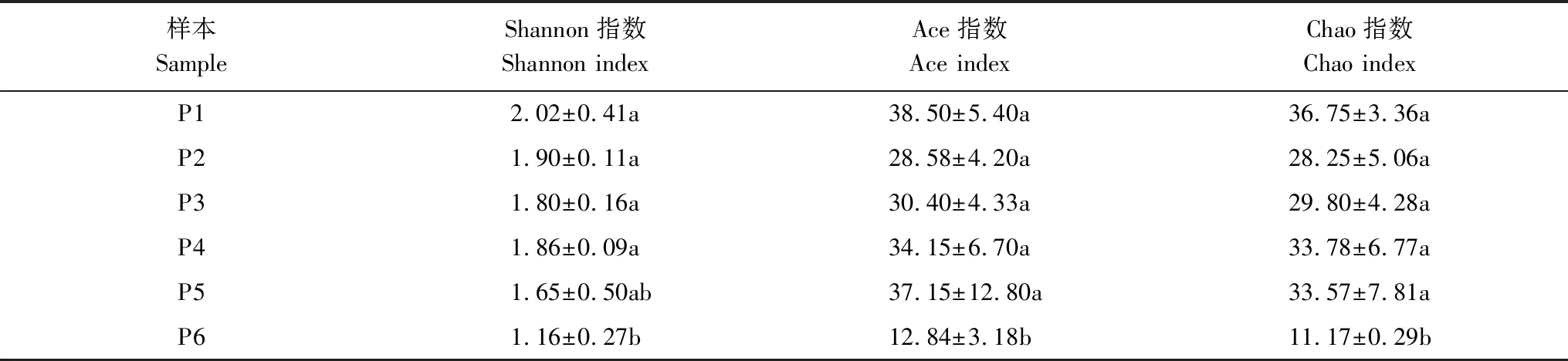
表2 土壤线虫群落α多样性
2.4 土壤线虫群落组成
2.4.1纲水平土壤线虫群落组成
纲水平土壤线虫群落分布如图3。在P1、P2、P3、P4、P5中,一类未分类线虫纲(unclassified_p_Nematoda)是土壤线虫优势群落,分别占各样本土壤线虫群落的75.31%、68.90%、70.30%、68.58%、72.54%;其次为刺嘴纲(Enoplea),分别占各样本土壤线虫群落的13.30%、18.59%、15.64%、22.41%、22.94%;再次为色矛纲(Chromadorea),分别占各样本土壤线虫群落的11.39%、12.51%、14.06%、9.01%、4.52%。但在距公路最远的P6,土壤线虫群落分布状况与P1、P2、P3、P4、P5均有较大不同,色矛纲线虫(Chromadorea)成为优势线虫群落,占68.20%;其次为一类未分类线虫纲(unclassified_p_Nematoda),占31.40%;刺嘴纲(Enoplea)最少,仅占0.40%。综上,P6的优势线虫群落与P1、P2、P3、P4、P5有很大不同。这可能与土壤状况有关。
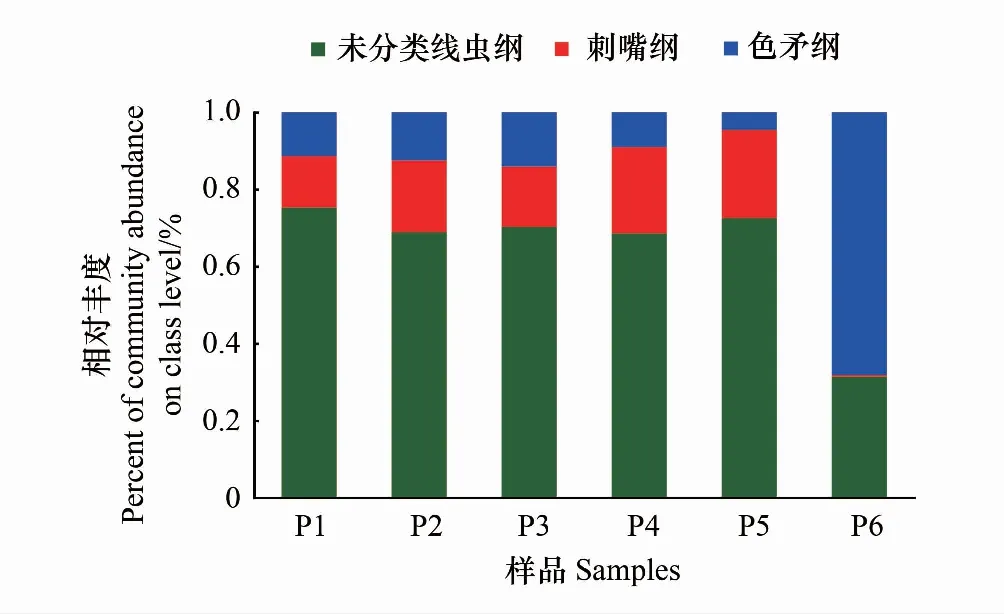
图3 纲水平土壤线虫的分布Fig.3 Frequence of class level of soil nematode communities
2.4.2目水平土壤线虫群落组成
目水平土壤线虫群落分布如图4。在P1、P2、P3、P4、P5中,一类未分类线虫目(unclassified_p_Nematoda)是优势土壤线虫群落,分别占各样本土壤线虫群落的75.31%、68.90%、70.30%、68.58%、72.54%;其次为矛线目(Dorylaimida)和垫刃目(Tylenchida),但在P3略有不同,P3的小杆目(Rhabdtida)相对较多。与P1—P5比较,在距公路最远的P6,小杆目(Rhabdtida)为优势土壤线虫群落,占59.68%;其次为一类未分类线虫目(unclassified_p_Nematoda),占31.40%;垫刃目(Tylenchida)最少,仅占8.52%。从各样本可知,目水平P6的优势土壤线虫群落与P1、P2、P3、P4、P5有很大不同。这可能与土壤状况有关。

图4 目水平土壤线虫群落的分布Fig.4 Frequence of order level of soil nematode communties
2.5 土壤线虫群落与重金属的关系
线虫门群落与土壤重金属的关系如图5。第一和第二主轴共解释了门水平土壤线虫群落相对丰度39.22%的方差变化,其中第一主轴解释了24.84%的方差变化,第二主轴解释了14.38%的方差变化。Cu、Pb、Zn、Cd四种重金属均与线虫门(Nematoda)正相关。
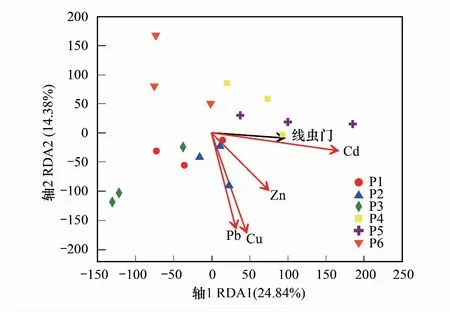
图5 线虫门与重金属的冗余分析 Fig.5 Redundancy analysis of Nematoda and heavy metal
土壤重金属与纲水平土壤线虫群落的关系如图6。第一主轴和第二主轴共解释了纲水平土壤线虫群落相对丰度72.65%的方差变化,其中第一主轴解释了69.16%,第二主轴解释了17.18%。Cu、Pb、Zn、Cd四种重金属互相均呈正相关。第一主轴上的Cu和Pb是最主要的重金属影响因子,相关系数分别为-0.9933和-0.9979;第二主轴上Cd是最主要的重金属影响因子,相关系数为-0.5553。四种重金属均与刺嘴纲(Enoplea)和一类未分类线虫纲(unclassified_p_Nematoda)群落正相关,与色矛纲(Chromadorea)群落负相关。
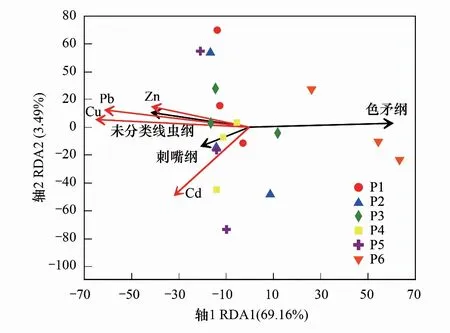
图6 纲水平土壤线虫群落与重金属的冗余分析 Fig.6 Redundancy analysis of class level of soil nematode communities and heavy metal
土壤线虫Chao指数与土壤重金属Cu、Pb、Zn、Cd的线性回归结果如图7,R2依次为0.648365、0.63319、0.55778、0.17770,说明Cu对不同位点线虫Chao指数差异的解释度最大,Cd的解释度最小,Cu、Pb、Zn对土壤线虫多样性的影响相对较强。

图7 土壤线虫多样性与重金属的回归分析Fig.7 Regression analysis of soil nematode diversity and heavy metals
3 讨论与结论
土壤线虫是对环境因子响应敏锐的一类土壤生物[25]。土壤线虫群落的分布特征受不同生态类型的影响[14]。试验区各位点土壤重金属与西藏土壤环境背景值[26]相比,Cu和Cd均超过背景值,Zn和Pb均未超过。对于试验区土壤环境背景值的调查尚未报道,但从距公路不同距离土壤重金属含量来看,较远的位点土壤重金属相对较低,理论上更靠近土壤环境背景值。一般在重金属浓度较高的土壤中,土壤线虫数量呈下降趋势[27-28]。由于本研究没有采用传统鉴定法,因此未能确定土壤线虫数量。在青藏高原高寒草甸,先前的研究认为Zn与线虫门(Nematoda)有较强的负相关性[22],Zn、Cu与纲水平线虫群落相关性较大[21]。在门水平,本研究认为Zn、Cu等重金属与线虫门(Nematoda)有较强的正相关,与纲水平线虫群落有较大相关,这与先前的门水平研究结果正好相反。此外,在纲、目水平上的线虫丰度组成与此前的研究差异很大,侯磊等[21]的研究结果表明在一般高寒草甸刺嘴纲(Enoplea)、矛线目(Dorylaimida)线虫占相对优势地位。Clarke等[29]认为一定浓度的Cu和Zn刺激线虫卵孵化过程继而增加线虫丰度,因此初步估计在青藏高原高寒草甸内部存在区域之间的土壤线虫群落结构上的差异,这种差异可能与不同区域土壤重金属含量有关。当然这种差异还需要在高寒草甸进行系统的土壤线虫群落调查才可较为深入地分析。
在青藏高原高寒草甸土壤重金属Cu、Pb、Zn、Cd含量较低的样本中,土壤线虫群落结构与重金属含量Cu、Pb、Zn、Cd较高样本差异明显。重金属含量较低样本以色矛纲(Chromadorea)和小杆目(Rhabdtida)土壤线虫为主要群落类型;重金属含量较高样本以一类未分类线虫纲(unclassified_p_Nematoda)和一类未分类线虫目(unclassified_p_Nematoda)为主要群落类型。说明重金属含量升高改变了高寒草甸土壤线虫纲水平和目水平的群落结构,使近自然状况下的色矛纲(Chromadorea)、小杆目(Rhabdtida)优势土壤线虫群落转变为一类未分类线虫纲(unclassified_p_Nematoda)、一类未分类线虫目(unclassified_p_Nematoda)优势土壤线虫群落。重金属Cu、Pb、Zn、Cd含量对土壤线虫群落α多样性产生影响。在重金属含量较低样本中,Shannon指数、Ace指数、Chao指数比重金属含量较高样本均要小,且整体上Shannon指数、Ace指数、Chao指数与重金属含量较高样本差异显著(P<0.05)。重金属Cu、Pb、Zn、Cd与线虫门(Nematoda)和刺嘴纲(Enoplea)正相关,但与色矛纲(Chromadorea)负相关。比较而言,在所讨论的四种重金属中,Cu对土壤线虫群落多样性变异具有最高的解释比例,Pb、Zn、Cd对变异的解释依次减小。本研究未能确定未分类线虫的纲、目具体名称,应当丰富对比生物文库基础数据,以利于今后土壤线虫群落的研究。此外,本文未结合线虫形态学鉴定,因此物种组成分析是基于相对丰度而言的,无法进行定量分析[15,30]。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
今日农业(2022年14期)2022-09-15 01:44:26
青海草业(2022年2期)2022-07-23 09:34:58
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
活力(2019年21期)2019-04-01 12:17:10
中成药(2018年2期)2018-05-09 07:20:04
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:02
天然产物研究与开发(2016年6期)2016-06-05 10:29:30
