
CO2加富及亏缺灌溉对菜豆中期与后期生长的影响
2021-06-02 20:55翟锡姣郑少文王帅全
中国瓜菜 2021年4期
翟锡姣 郑少文 王帅全
摘 要:为了探明菜豆中期与后期生长对CO2加富与亏缺灌溉的生理响应,试验设两个CO2水平:正常大气浓度和倍增CO2浓度,两个灌溉水平:自然灌溉12次和亏缺灌溉6次,研究CO2加富及亏缺灌溉对菜豆生长的影响。结果表明,CO2加富显著提高菜豆生长中期株高与干质量,净光合速率(Pn)与胞间CO2浓度(Ci)分别显著降低15.48%、37.67%,气孔导度(Gs)显著提高95.83%,菜豆生长后期的叶绿素a/b(Chla/b)显著提高12.29%,水分利用效率(WUE)显著提高46.51%,Ci显著降低12.87%。CO2加富显著提高亏缺灌溉下生长中期菜豆的株高与干质量,但显著降低根冠比,显著提高叶绿素含量以及Gs;CO2加富下生长后期的核酮糖-1,5-二磷酸羧化酶(Rubisco)及果糖-1,6-二磷酸酶(FBPase)活性分别显著提高78.05%与88.69%,CO2加富使亏缺灌溉下碳酸酐酶(CA)与FBPase活性分别显著提高83.73%与64.84%。综上所述,CO2加富提高菜豆生长中期对亏缺灌溉的适应性,对生长后期影响作用减小。
关键词:菜豆;CO2加富;亏缺灌溉;中期与后期;生长发育
中图分类号: S643.1 文献标识码: A 文章编号:1673-2871(2021)04-105-07
Abstract: In order to explore the physiological response of growth of kidney bean at middle and late stages to the CO2 enrichment and deficit irrigation, two CO2 levels were set for the test: normal atmospheric concentration and doubled CO2 concentration, two irrigation levels were carried out: natural irrigation for 12 times(CK)and deficit irrigation for 6 times, the effects of CO2 enrichment and deficit irrigation on morphological indicators, photosynthetic indicators and key enzyme activities of carbon assimilation in kidney beans were studied. The results showed that: CO2 enrichment could improve the plant height and dry quality, the net photosynthetic rate(Pn)and intercellular CO2 concentration(Ci)were reduced by 15.48% and 37.67% respectively, stomatal conductance(Gs)was increased by 95.83% in the middle growth stage, chlorophyll a/b was increased by 12.29% and water use efficiency(WUE)was increased by 46.51%, Ci was decreased by 12.87% in the late growth period; CO2 enrichment increased the plant height, dry quality, chlorophyll content and Gs, but decreased the root-cap ratio in the medium growth stage under deficit irrigation. The activities of Rubisco and FBPase were increased by 78.05% and 88.69% respectively under CO2 enrichment. The activities of CA and FBPase were increased by 83.73% and 64.84% respectively under CO2 enrichment and deficit irrigation. CO2 enrichment improved the adaptability of kidney bean to deficit irrigation in the middle growth stage but its effect on the late growth period was reduced.
Key words: Kidney beans; CO2 enrichment; Deficit irrigation; Middle stage and late stage; Growing development
CO2是植物光合作用的必要原料,在晴天、強光下日光温室中的植物光合能力很强,CO2浓度下降很快,CO2常常处于亏缺状态。而CO2浓度的升高对作物的影响也并非是独立的,水分对植物生长发育也发挥着重要作用。目前水资源匮缺、水分利用率较低制约作物生产和农业发展,同时也直接或间接地影响着生态环境。有研究表明CO2加富能够提高作物株高、茎粗、叶片指数及叶绿素含量,在很大程度上能够有效提高光合作用[1]。在水分亏缺时,CO2加富可以提高植株净光合速率,降低蒸腾速率和气孔导度[2]。因而揭示作物生长发育、光合效率等对CO2加富及水分变化的响应对预测未来大气CO2浓度升高下作物生产力与需水规律的变化具有重要意义。
菜豆(Phaseolus vulgaris L.)籽粒含有丰富的蛋白质、碳水化合物以及纤维素,在许多国家和地区作为主要营养来源之一作物被广泛种植。有研究表明蔬菜在不同时期需水量不同,对蔬菜的影响也不同[3]。菜豆前期对水分需求较少,长期过量施用CO2在植株中产生光合适应现象,造成CO2肥料浪费[4]。目前,已有大量研究主要集中在全生育期或苗期CO2与水分对作物造成的影响,关于CO2加富及亏缺灌溉对温室菜豆生长中期与后期影响的研究报告较少。笔者采用盆栽法,测定菜豆中期及后期两个时期各项形态及光合指标,旨在为中国北方地区设施栽培提供理论依据。
1 材料与方法
1.1 材料
供试菜豆品种为超早一尺秀,抗性强,分枝力强,蔓生,由太原市乐丰园种业有限公司提供。
1.2 试验地概况
试验于山西农业大学园艺学院园艺站东西走向的日光温室中进行。试验所用容器为聚乙烯塑料盆,上口直径34 cm,下口直径21 cm,高28 cm,盆土由田园砂壤土与草炭按体积比3∶1混合配置,每盆装15 kg。混合基质养分(w)状况:碱解氮质量分数45.15 mg·kg-1,速效磷质量分数15.49 mg·kg-1,速效钾质量分数353.3 mg·kg-1。在2019年4月14日选取籽粒饱满、大小一致的种子进行温汤浸种,2019年4月15日进行播种,播种时深度约1.5 cm,表层覆混合的蛭石与田园土0.5 cm,每盆3穴,每穴播种3粒,每个处理5盆,3次重复,共60盆。在单叶展开至第一片复叶展开前将每穴定苗2株,常规田间管理。
1.3 试验设计
试验为完全随机设计,前期菜豆生长处于同一水平,苗龄45 d后开始不同水分处理,试验灌溉设2个水平:设计中期至后期灌溉12次(对照W0),每3 d灌溉1次,中期至后期灌溉6次(W1),每6 d灌溉1次,每次灌溉量为2 L。设2个CO2浓度水平:正常大气浓度(400±20) ?mol·mol-1和倍增CO2浓度(800±20) ?mol·mol-1,(表示为C),共4个处理W0(正常大气环境下灌溉12次)、CW0(CO2加富环境下灌溉12次)、W1(正常大气环境下灌溉6次)、CW1(CO2加富环境下灌溉6次),在菜豆播种45 d后开始施放;用塑料薄膜隔離出相对独立的空间,其中富碳环境气源为CO2钢瓶,选择在晴天6:30—8:30将气体通过塑料管均匀地施放到密闭空间中,并通过自动控制系统控制CO2浓度始终维持在设定水平,阴雨天不施。
1.4 测定指标
1.4.1 形态指标 株高采用卷尺测定主干根部到生长点的高度;茎粗采用游标卡尺测定地面以上1 cm处;叶面积取自植株由上至下第3片完全展开的功能叶片,使用智能叶面积仪(Yaxin-1241)进行测定;将植株根部洗净,分地上部分与地下部分,于110 ℃杀青30 min,80 ℃烘干48 h,至恒质量时进行称量,即为地上部分干质量与地下部分干质量,根冠比=地下部分干质量/地上部分干质量。
1.4.2 光合相关参数 于晴天9:00—11:00在自然光照条件下用Li-6400XT光合仪进行光合指标的测定,测定植株由上至下第3片完全展开的功能叶的中间叶片,测定时使用开放气路,光合仪的光量子通量密度设置为1000 μmol·m-2·s-1,叶室温度为25 ℃,测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr),水分利用效率(WUE)=Pn/Tr。
叶绿素含量的测定采用80%丙酮浸提法,置于黑暗下24 h,用分光光度计在645 nm与663 nm下比色测定。按公式计算叶绿素a、叶绿素b含量和叶绿素a/b:Ca=12.72×A663-2.59×A645; Cb=22.88×A645-4.67×A663。
光合碳同化关键酶测定:按照ELISA试剂盒说明书(北京索莱宝科技有限公司提供)测定菜豆生长后期叶片中的碳酸酐酶(CA)、核酮糖-1,5-二磷酸羧化酶(Rubisco)、果糖-1,6-二磷酸酶(FBPase)活性。
1.5 数据分析
用Microsoft Excel 2013软件对数据进行处理和作图,用SPSS20.0和SAS9.0软件进行统计分析。
2 结果与分析
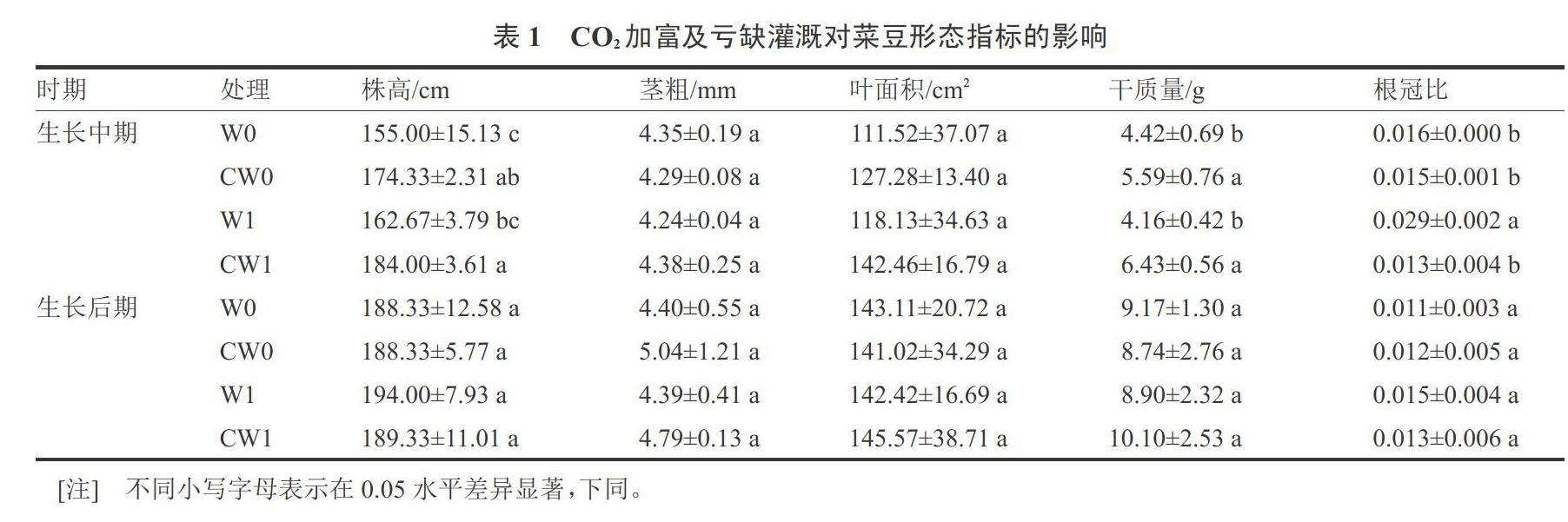
2.1 CO2加富及亏缺灌溉对菜豆形态指标的影响
由表1可知,在菜豆生长中期,单独CO2处理下,CW0处理与W0处理相比,株高与干质量分别提高12.47%、26.47%,两处理间株高、干质量均具有显著性差异;两处理间根冠比降低但差异不显著。单独水分处理下,W1与W0处理相比,株高提高4.94%,干质量下降5.88%,但两处理间差异不显著;两处理间根冠比显著提高81.25%。复合处理下,CW1与W1处理相比,株高与干质量分别提高13.11%与54.57%;根冠比降低55.17%,两处理间差异显著。
在菜豆生长后期,各处理株高、茎粗、叶面积、干质量及根冠比之间没有显著差异。
在菜豆生长中期和后期,各处理之间茎粗与叶面积没有显著差异,表明CO2加富与亏缺灌溉对菜豆茎粗与叶面积没有显著影响。
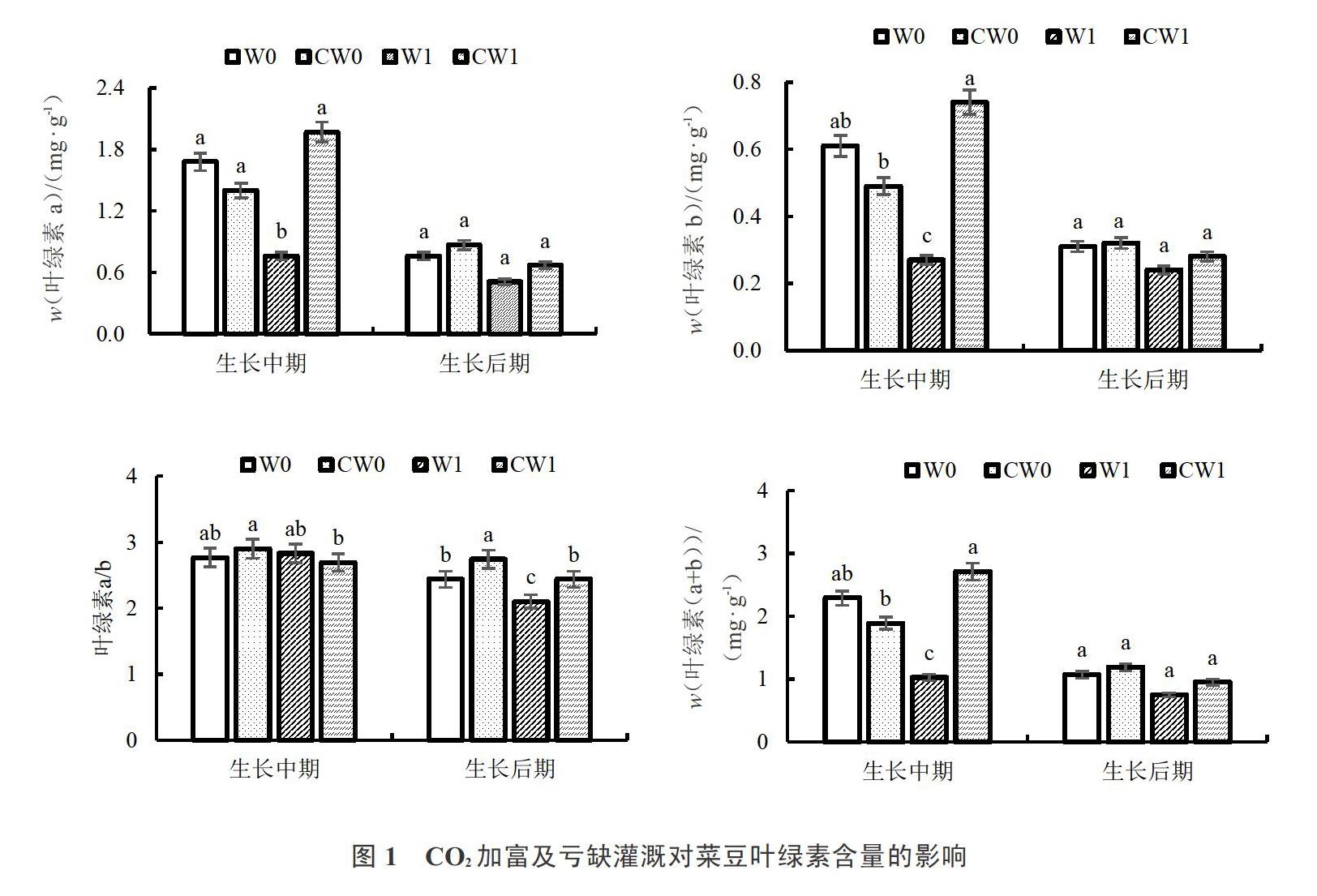
2.2 CO2加富及亏缺灌溉对菜豆叶绿素的影响
由图1可以看出,单独CO2处理下,CW0与W0处理相比,在菜豆生长中期,Chla、Chlb、Chl(a+b)含量分别降低16.67%、19.67%、17.47%,Chla/b提高,但两处理间各指标差异均不显著;在菜豆生长后期,Chla、Chlb、Chl(a+b)含量分别提高14.47%、3.22%、11.21%,两处理间3项指标差异均不显著,而Chla/b显著提高12.29%。
单独水分处理下,W1与W0处理相比,在菜豆生长中期,Chla、Chlb、Chl(a+b)含量分别显著降低54.76%、55.74%、55.02%,而Chla/b提高但差异不显著;在菜豆生长后期,Chla、Chlb、Chl(a+b)含量均降低但差异不显著,Chla/b显著降低13.39%。
复合处理下,CW1与W1处理相比,在菜豆生长中期Chla、Chlb、Chl(a+b)含量分别显著提高159.21%、174.07%、163.11%,Chla/b降低但差异不显著;在菜豆生长后期Chla、Chlb、Chl(a+b)含量提高但没有显著性差异,Chla/b显著提高16.19%。
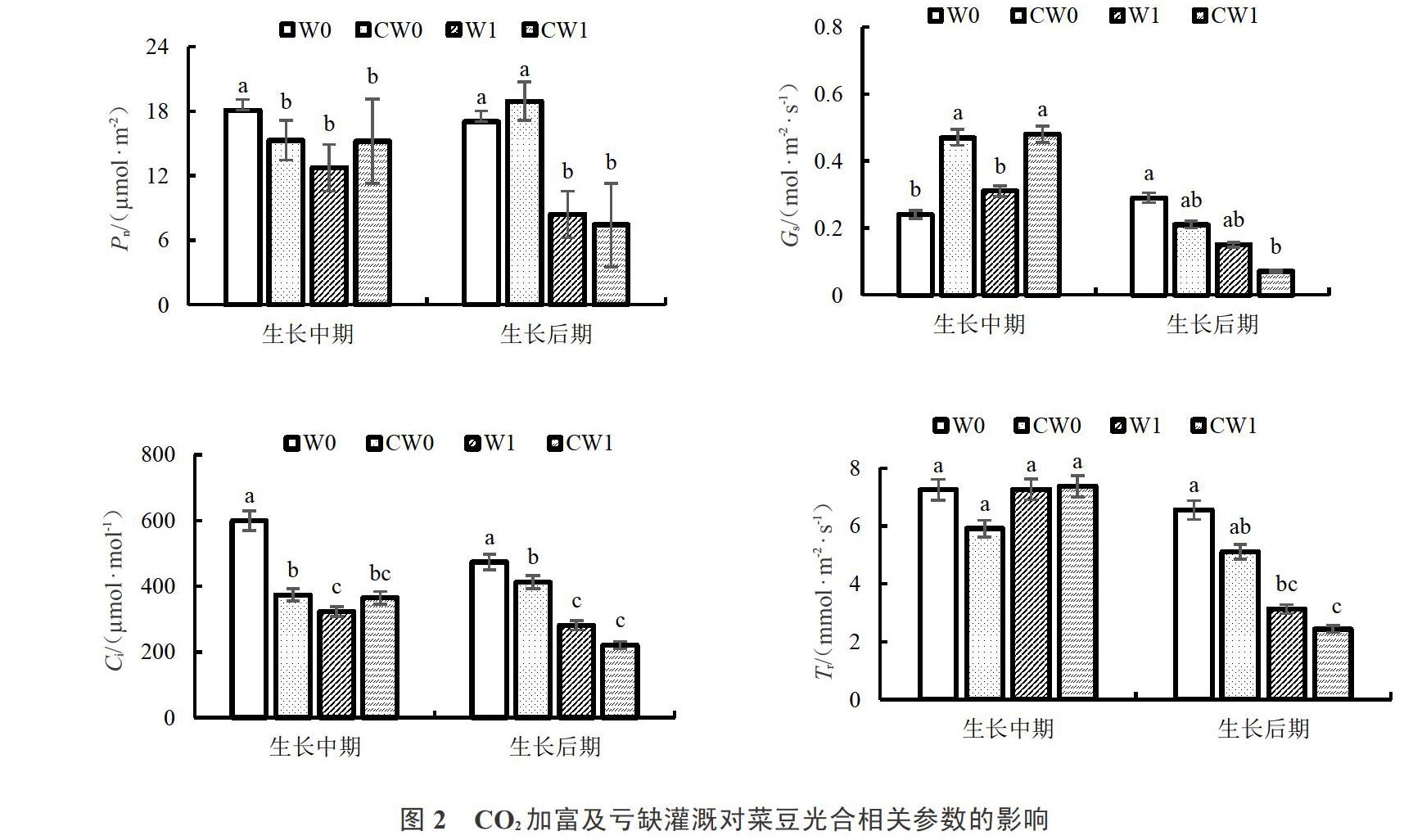
2.3 CO2加富及亏缺灌溉对菜豆光合指标的影响
由图2可以看出,单独CO2处理下,CW0与W0处理相比,在菜豆生长中期Pn与Ci分别显著降低15.48%与37.67%,Gs显著提高95.83%;Tr降低18.62%,WUE提高1.57%,差异均不显著;在生长后期Pn提高,Gs与Tr降低,但都不具有显著差异性,Ci在生长后期显著降低12.87%,WUE显著提高46.51%。
单独水分处理下,W1与W0处理相比,在菜豆生长中期Pn、Ci与WUE分别显著降低29.46%、46.13%与31.37%,Gs与Tr均提高但差异不显著;在菜豆生长后期,Pn、Ci与Tr分别显著降低50.56%、40.7%、52.21%,Gs降低,WUE提高,但均不具有显著性差异。表明亏缺灌溉使两个生长时期的菜豆光合速率降低,Ci也降低;生长中期WUE降低,表明此时期Pn降幅程度较Tr大。
复合处理下,CW1与W1处理相比,在菜豆生长中期Pn、Tr、Ci、WUE均提高,但无显著性差异,Gs显著提高54.84%;在后期Pn、Gs、Ci、Tr均降低,WUE提高,但均无显著性差异。表明CO2加富对亏缺灌溉下两个时期的Pn影响不显著,CO2加富提高亏缺灌溉下生长中期的Gs,促进气孔开放,加快对CO2吸收速度,提高光合能力,对后期Gs无显著影响,两个时期Ci、Tr与WUE对CO2加富与水分协同变化的响应不敏感。
由表2可知,在菜豆生长中期,净光合速率与叶绿素a、叶绿素b及叶绿素(a+b)均呈显著正相关,与胞间CO2浓度及水分利用效率均呈极显著正相关;蒸腾速率不仅与叶绿素a/b呈显著负相关,还与水分利用效率呈显著负相关。
由表3可知,在菜豆生长后期,净光合速率只与叶绿素a含量呈显著正相关,与气孔导度、胞间CO2浓度及蒸腾速率呈极显著正相关;气孔导度与胞间CO2浓度呈显著正相关,蒸腾速率与胞间CO2浓度、氣孔导度呈极显著正相关。
2.4 CO2加富及亏缺灌溉对菜豆碳同化关键酶活性的影响
由图3可以看出,在菜豆生长后期,单独CO2处理下,CW0与W0处理相比,菜豆CA活性提高22.25%,差异不显著;Rubisco活性显著提高78.05%,FBPase活性显著提高88.69%,表明CO2加富对菜豆CA活性没有显著影响,但可显著提高Rubisco、FBPase活性。
单独水分处理下,W1与W0处理相比,菜豆CA活性降低16.25%,Rubisco活性提高26.83%,FBPase活性提高11.3%,但差异均不显著。表明亏缺灌溉对3种酶活性没有显著影响。
复合处理下,CW1与W1处理相比,菜豆CA显著提高83.73%,FBPase活性显著提高64.84%,Rubisco活性提高20.19%,但差异不显著。表明CO2加富显著提高亏缺灌溉下CA与FBPase活性升高,对Rubisco活性无显著影响。
3 讨论与结论
3.1 CO2加富及亏缺灌溉对菜豆形态指标的影响
水分亏缺抑制植物生长,致使株高及干物质积累降低,通过减少叶面积以及气孔关闭来减少水分的蒸发,并且对地下部分生长的抑制作用低于地上部分,将植株体内资源优化配置, 在有限范围内维持正常活动[5-6]。本研究结果表明,生长中期亏缺灌溉显著提高根冠比,促进地下部的生长发育。有研究表明高浓度CO2促进了地上部分和地下部分生物量的累积,但是地上部分生物量增加占主导地位[7]。本试验结果显示,在菜豆生长中期CO2加富下缓解亏缺灌溉对植株地上部生长的抑制作用。
3.2 CO2加富及亏缺灌溉对菜豆叶绿素含量的影响
叶绿素是参与光合作用光能吸收、传递和转化的重要色素,有研究表明亏缺灌溉有利于提高作物的叶绿素含量,增强作物抵抗水分胁迫的能力[8],也有研究表明水分亏缺往往造成植株叶片叶绿素合成不足,进而降低光合作用[9]。本研究结果表明,在菜豆生长中期,亏缺灌溉使叶绿素含量下降,这可能是水分亏缺使菜豆体内大量积累活性氧促使叶绿体色素降解[10]。在菜豆生长后期,亏缺灌溉降低Chla/b,Chla/b能够反映补光色素复合体LHCII在植株所有含叶绿体结构中所占的比例,其比值越大说明补光能力越强,Chla/b比值降低,表明在此时期菜豆叶片补光能力减弱,减弱光合作用中光能的吸收和转化。有研究表明,大气CO2浓度升高后,辣椒中叶绿素a、叶绿素b、叶绿素(a+b)含量均有增加趋势[11]。试验结果显示,菜豆生长中期CO2加富对叶绿素含量没有显著影响,可能是在自然灌溉条件下叶绿素含量已达到较高水平,CO2加富使叶绿素含量提高空间不大,CO2加富促进此时期亏缺灌溉条件下菜豆叶片叶绿素的合成,这与田露等[5]研究结果一致。
3.3 CO2加富及亏缺灌溉对菜豆光合相关参数的影响
水分是植物进行光合作用必不可少的反应物,水分亏缺下植株自身含水量降低,使Pn、Tr及Gs依次降低,导致光合速率下降[12],本研究结果表明,在菜豆生长中期,亏缺灌溉降低Pn、WUE及Ci,这与以上研究结果一致。水分亏缺条件下植株仍能正常生长的关键是具备高水平的水分利用效率,有研究表明植株负面影响光合速率,有助于提高WUE[13],此时期亏缺灌溉下WUE降低,可能是净光合速率降低,而蒸腾速率没有显著变化,水分利用效率降低主要是光合导致,此时期水分利用效率与净光合速率呈显著正相关。本研究结果显示,在菜豆生长后期,亏缺灌溉降低菜豆Pn、Tr与Ci,引起净光合速率下降有气孔限制和非气孔限制两种因素,两个时期亏缺灌溉对Gs都没有显著影响,表明气孔能够保持一定水平的开度,而Pn显著下降,表明菜豆植株Pn显著下降主要是非气孔因素导致。
张振花等[14]研究表明,CO2加富通常会提高Pn和Ci,并通过降低植株叶片Gs,消耗相对较少的水分,降低Tr,间接提高WUE。本研究表明,在菜豆生长中期,CO2加富降低Pn与Ci,显著提高Gs,在菜豆生长后期CO2加富提高WUE,显著降低Ci,Gs与Tr变化趋势一致,都呈下降趋势。生长中期研究结果与张振花等[14]的相反,可能与环境因子、作物品种或作物自身变化有关[15],还需进一步研究。Sreeharsha等[16]研究表明CO2加富促进豌豆在苗期和生殖生长期Gs升高,因而CO2加富降低叶片Gs不是普遍的规律。有研究表明高CO2浓度下发生水分亏缺时,CO2施肥效应会受到一定程度的抑制[17]。本试验研究结果显示,亏缺灌溉下CO2加富除菜豆生长中期Gs显著提高外,对光合其他指标均没有产生显著影响,表明CO2施肥效应未受到抑制。
3.4 CO2加富及亏缺灌溉对菜豆碳同化关键酶的影响
核酮糖-1,5-二磷酸羧化酶(Rubisco)是植物进行光合碳同化的关键酶,有研究表明水分亏缺会引起Rubisco活性降低,影响叶片的气体交换和光合作用的正常进行[18-19]。在一定范围内,随着CO2浓度的增加Rubisco的活性上升[20],但是也有一些研究表明,高CO2条件下生长的叶片Rubisco活性有所降低[21]。试验结果表明,CO2加富提高Rubisco活性,亏缺灌溉处理对Rubisco的活性没有显著影响,CO2加富对亏缺灌溉条件下的Rubisco活性也没有显著影响。碳酸酐酶(CA)是转运无机碳的关键酶,能够有效催化HCO3-和CO2的可逆水合反应,抑制CA活性会降低Rubisco的羧化活性,从而降低对CO2的固定,最终降低光合速率[22],适量升高的CO2浓度能够提高CA活性[23]。试验结果表明,亏缺灌溉下CO2加富提高CA活性。有研究结果显示,随着水分胁迫程度加深,果糖-1,6-二磷酸酶(FBPase)活性明显降低,而FBPase对一定程度的水分亏缺具有“忍耐力”,在水分胁迫下能保持高的酶活性来促进光合产物的运输,减轻反馈抑制,有助于植物适应不良环境[24]。试验结果表明,CO2加富可以提高FBPase活性,亏缺灌溉对FBPase活性没有显著影响,表明FBPase能够适应亏缺灌溉处理,CO2加富提高亏缺灌溉下FBPase活性。亏缺灌溉对CA、Rubisco、FBPase活性没有显著影响,表明3种碳同化关键酶对亏缺灌溉都有一定的适应性。
综上所述,CO2加富与亏缺灌溉对菜豆生长中期生长发育影响较大,对生长后期影响减弱。而由于笔者是在盆栽试验条件下得出的结果,还有待通过大田生产实践来进一步检验。
参考文献
[1] 高宇,崔世茂,宋阳,等.CO2加富对温室辣椒幼苗生长及光合特性的影响[J].作物杂志,2017(5):80-84.
[2] 李清明,刘彬彬,艾希珍.CO2浓度倍增对干旱胁迫下黄瓜幼苗膜脂过氧化及抗氧化系统的影响[J].生态学报,2010,30(22):6063-6071.
[3] CHEN S,ZHOU Z J,ANDERSEN M N,et al.Tomato yield and water use efficiency–coupling effects between growth stage specific soil water deficits[J].Acta Agriculturae Scandinavica,Section B- Soil & Plant Science,2015,65(5):460-469.
[4] 杨志刚.长期CO2加富对温室辣椒结果期生长的影响及生理基础研究[D].呼和浩特:内蒙古农业大学,2016.
[5] 田露.水稻幼苗对高浓度CO2和水分胁迫的生理响应研究[D].沈阳:沈阳师范大学,2016.
[6] 刘长利,王文全,崔俊茹,等.干旱胁迫对甘草光合特性与生物量分配的影响[J].中国沙漠,2006,26(1):142-145.
[7] 李旭芬,石玉,李斌,等.CO2加富对盐胁迫下番茄幼苗生长和渗透调节特性的影响[J].西北农业学报,2019,28(8):1309-1316.
[8] 常莉飞,邹志荣.调亏灌溉对温室黄瓜生长发育产量及品质的影响[J].安徽农业科学,2007,35(23):7142-7144.
[9] 解斌,李俊豪,赵军,等.干旱胁迫对2种梨砧木生长及叶绿素荧光特性的影响[J].西北农业学报,2019,28(5):753-761.
[10] SOUZA R P,MACHADO E C,SILVA J A B,et al.Photosynthetic gas exchange,chlorophyll fluorescence and some associated metabolic changes in cowpea(Vigna unguiculata)during water stress and recovery[J].Envi-ronmental and Experimental Botany,2004,51(1):45-56.
[11] 袁蕊,聂磊云,郝兴宇,等.大气CO2浓度升高对辣椒光合作用及相关生理特性的影响[J].生态学杂志,2017,36(12):3510-3516.
[12] 吴玲利,雷小林,龚春,等.干旱胁迫对白木通光合生理特性的影响[J].中南林业科技大学学报,2015,35(11):68-73.
[13] 姚有华,白羿雄,吴昆仑.亏缺灌溉对藜麦光合特性、营养品质和产量的影响[J].西北农业学报,2019,28(5):713-722.
[14] 张振花,孙胜,刘洋,等.增施CO2对温室番茄结果期叶片光合特性的影响[J].生态学杂志,2018,37(5):1398-1402.
[15] 張振贤,程智慧.高级蔬菜生理学[M].北京:中国农业大学出版社,2008:28-34.
[16] SREEHARSHA R V,REDDY A R.Dynamics of vegetative and reproductive growth patterns in pigeonpea(Cajanus cajan L.)grown under elevated CO2[J].Procedia Environmental Sciences,2015,29:147-148.
[17] 高素华,郭建平,周广胜.高CO2浓度下羊草对土壤干旱胁迫的响应[J].中国生态农业学报,2002,10(4):31-33.
[18] 杨青华,郑博元,李蕾蕾,等.外源NO供体对水分亏缺下玉米叶片碳同化关键酶及抗氧化系统的影响[J].作物学报,2018,44(9):1393-1399.
[19] 李翔,桑勤勤,束胜,等.外源油菜素内酯对弱光下番茄幼苗光合碳同化关键酶及其基因的影响[J].园艺学报,2016,43(10):2012-2020.
[20] 姜伟,孙萍,罗文熹,等.CO2浓度增加对黄瓜叶片气孔和Rubisco活性的影响[J].莱阳农学院学报,1998,15(2):3-5.
[21] 唐如航,郭连旺,陈根云,等.大气CO2浓度倍增对水稻光合速率和Rubisco的影响[J].植物生理学报,1998,34(3):3-5.
[22] 黄瑾,夏建荣,邹定辉.微藻碳酸酐酶的特性及其环境调控[J].植物生理学通讯,2010,46(7):631-636.
[23] 刘清钰,莫华,杨海波.饱和CO2对亚心形扁藻生长及碳酸酐酶活性的影响[J].安徽农业科学,2017,45(31):61-64.
[24] 刘艳,王丽雪,王有年.水分胁迫对苹果叶片叶绿体1.6-二磷酸果糖磷酸酯酶活性的影响[J].内蒙古农业大学学报(自然科学版),2002,23(4):42-45.
猜你喜欢
山东农业科学(2016年12期)2017-01-21
现代农业科技(2016年20期)2016-12-20
食品与健康(2015年7期)2015-07-14
故事大王(2014年3期)2014-05-21
探索财富(2013年10期)2013-11-18
作文大王·中高年级(2008年5期)2008-07-31
