
基于生长过程的海南桉树纸浆林土壤理化性质和植物多样性分析
2021-06-01 03:45张沛健徐建民卢万鸿林志锋尚秀华
中南林业科技大学学报 2021年5期
张沛健,徐建民,卢万鸿,林志锋,尚秀华
(1.中国林业科学研究院 热带林业研究所,广东 广州 510520;2.南京林业大学,江苏 南京 210037;3.国家林业和草原局 桉树研究开发中心,广东 湛江 524022;4.海南金华林业有限公司,海南 儋州 578101)
林木生长与光照、能量、土壤水分和养分等环境因子关联密切,而植被和土壤是一个密不可分的整体,土壤作为生态系统中物质和能量交换的重要介质,为林下物种生长和发育提供必需的水分和矿质营养,是林木赖以生存的重要基础[1-3]。土壤条件的优劣可直接影响森林生态系统植被的更新和演替[4-5],同时生态系统的群落构成和多样性又影响着土壤的形成、发育以及土壤养分的有效性[6]。因此,研究人工林林下物种配比、组成结构以及土壤养分,对深入了解林木生长状况和森林群落演替过程以及经营管理具有重要参考价值[7]。
目前,桉树人工林面积已超过546万hm2,年产4 500多万m3木材,为我国木材生产作出了重要贡献[8-9],也是我国最重要的纸浆原材料之一。海南作为我国桉树的主要种植区,现有栽培面积12.9万hm2,大部分以生产纸浆材为经营目标。由于桉树种植以纯林为主,所以桉树林生态稳定性相对较弱,导致生态问题日益加剧,不利于桉树人工林的可持续发展。但桉树人工林林下植被物种分布、结构特征以及多样性对桉树生产力和生态系统功能具有显著影响[10],林下植被又是桉树林生态系统的重要组成部分,在维护林地地力和促进人工林养分循环中起着重要作用[11],因而桉树林下植物多样性及土壤特征的研究日益受到重视。相关学者对桉树人工林群落植物多样性和土壤理化性质进行了较多研究[12-16],也有学者对海南桉树林下植物多样性做了研究,是以桉树一个林龄的主要植被和土壤性质进行简单的分析[17]。但是对桉树人工林生长发育全过程中植物多样性和土壤特征关系的动态变化趋势及其规律的研究未见报道,群落之间的内在联系尚不明确。本研究以海南尾细桉(无性系TH9128-1)纸浆林为研究对象,根据林业行业标准(2017)中桉树的龄组划分标准[18],对尾细桉幼龄林(1~2 a)、中龄林(3~4 a)、近熟林(5 a)、成熟林(6~7 a)、过熟林I(10 a)、过熟林II(15 a)6个不同龄组的林木生长状况、土壤理化性质、林下植物多样性进行研究,探讨其动态变化趋势和规律,深入分析三者之间的关系,从而探索桉树纸浆林生长的主要影响因子,为以后的桉树种植和经营提供理论依据,促进桉树产业走上可持续发展道路。
1 研究区概况
海南岛(18°10′~20°10′N,108°37′~111°03′E)位于中国最南端,属于热带季风气候,年日照时数约为2 500 h,年平均气温在23~26℃之间,年平均降水量为1 000~2 500 mm,以8—9月份降水量最为充沛,雨热同期,干湿季明显,地带性土壤为砖红壤,热带风暴和台风灾害常有发生。研究试验样地位于海南低山地区的桉树主产区,选择地貌、海拔、土壤类型等立地条件相近的同品系尾细桉人工林作为研究区域,因调查的试验林为同一企业经营,所以各林地的人为经营措施(如整地造林、施肥、除草等抚育管理)基本一致。
2 研究方法
2.1 样地设置和样品采集
采用空间替代时间的方法,2018—2019年在研究区的尾细桉纸浆林内按不同林龄(1~7 a、10 a和15 a)分别设置20 m×20 m的标准样地,幼龄林设有9个标准样地,中龄林设有33个标准样地,近熟林设有7个标准样地,成熟林设有7个标准样地,过熟林I设有5个标准样地,过熟林II设有9个标准样地,共调查标准样地70个,记录每个标准样地的地理坐标、土壤类型、海拔、土层厚度等立地因子,对标准样地内的桉树进行每木检尺,记录树高(H)和胸径(D),并计算单株材积(V)。每个样地选取5株优势木,统计并记录优势木树高和胸径。同时,在每个标准样地内沿对角线选取3个5 m×5 m的样方调查灌木层,在灌木层样方四角和中心位置设置5个1 m×1 m 的样方调查草本层,分别统计并记录各样方中植被种名、株数/丛数、盖度和高度。在标准样地的每个灌木层样方中随机挖取深60 cm的土壤剖面,用环刀取原状土,带回实验室进行土壤物理性质分析;同一土壤剖面从上到下取土壤样品,混合均匀后装入密封袋,带回实验室,经过处理后测定其化学性质。各龄组样地的基本情况见表1。
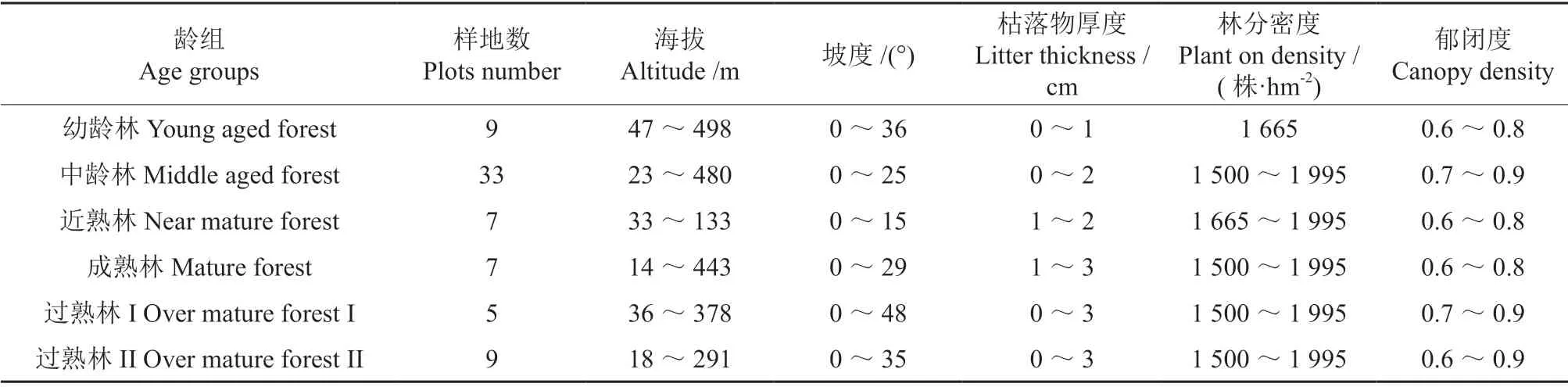
表1 不同龄组标准样地基本情况Table 1 Basic conditions of plots of different age groups
2.2 土壤理化性质的测定方法
采用环刀法测定土壤含水量和容重[19]。采用四分法取1 kg左右的混合土样,经风干后研磨过筛再测定土壤化学性质,各指标的测定方法为:pH值采用氯化钾浸提-电位法测定;有机质含量采用重铬酸钾氧化-容量法测定;全N含量采用凯氏法测定;全P含量采用碱熔-钼锑抗比色法测定;全K含量采用氢氧化钠碱熔-火焰光度法测定;碱解N采用碱解扩散法测定;有效P含量采用双酸浸提-钼锑抗比色法测定;速效K含量采用乙酸铵浸提-火焰光度法测定[20]。
2.3 数据处理
根据野外调查数据,分别计算尾细桉人工林林下灌木层和草本层的丰富度指数(R)[21,23]、辛普森(Simpson)多样性指数(D)、香农-威纳(Shannnon-Wiener)多样性指数(H)、均匀度指数(J)[24-25],计算公式如下。
丰富度指数:

Simpson多样性指数:

Shannon多样性指数:

Pielou均匀度指数:

式中:S表示样方内出现的物种数目;Pi为相对重要值,即第i个物种的个数占据所有物种总数的比例。
采用Excel 2010软件对数据进行统计和制图,运用SPSS 24.0软件进行方差分析和相关性分析。
3 结果与分析
3.1 不同龄组尾细桉人工林生长状况
由表2可知,不同龄组尾细桉林分的胸径、树高和材积随着生长发育呈增长趋势,与幼龄林相比,中龄林、近熟林、成熟林、过熟林的胸径增幅分别为74.44%、103.02%、118.51%、153.92%、189.33%,单株材积的增幅分别为307.11%、478.66%、519.49%、961.36%、1 384.84%。在尾细桉林不同生长发育阶段,林分生长量各指标呈显著增加的趋势,其中中龄林、近熟林生长量增幅较大,成熟林和近熟林之间增幅较小,由于过熟林龄组跨度较大,所以过熟林间增幅也较大。优势木的树高和胸径随着生长发育也呈增加趋势,与幼龄林相比,中龄林、近熟林、成熟林、过熟林的优势木树高增幅分别为62.32%、85.94%、102.14%、131.16%、178.40%,优势木胸径的增幅分别为73.88%、100.14%、143.26%、152.92%、224.22%。
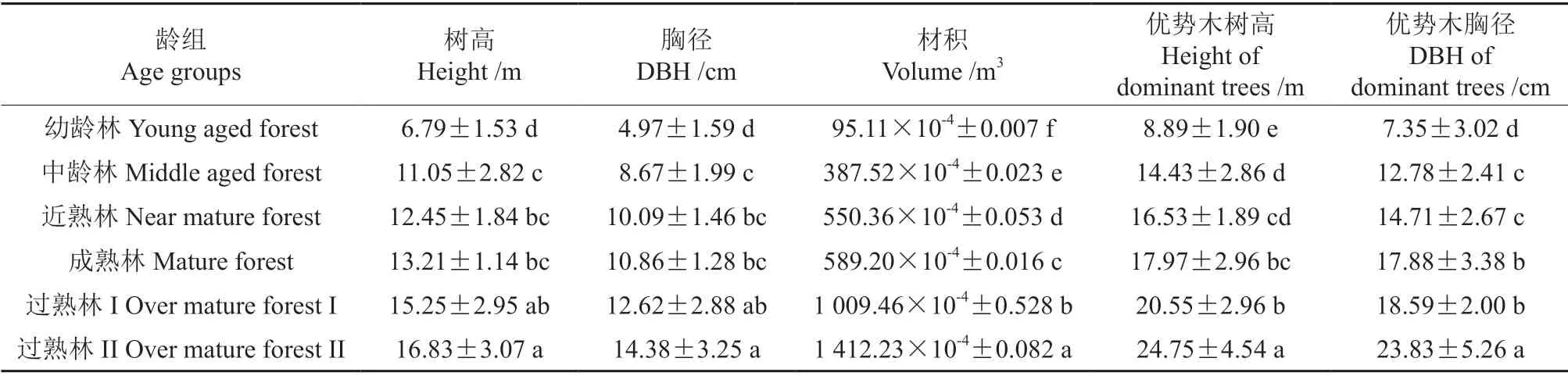
表2 不同龄组尾细桉生长量及优势木生长状况†Table 2 Growth and its dominant trees of E.urophylla×E.tereticornis in different age groups
3.2 不同龄组尾细桉人工林土壤理化性质
3.2.1 尾细桉人工林土壤物理性质
不同龄组尾细桉人工林土壤物理性质如表3所示。由表3可知,尾细桉不同龄组对土壤物理性质影响差异性显著。随着林木的生长发育,土壤容重呈先增加后降低的趋势;土壤含水量、总孔隙度、毛管孔隙度和空气孔隙度随着林龄的增加整体上变化一致,均呈先降低后增加的趋势,且都是在过熟林I达到最低,过熟林II有所增加。

表3 不同龄组尾细桉人工林土壤物理性质Table 3 Physical properties of soil in E.urophylla×E.tereticornis at different age groups
3.2.2 尾细桉人工林土壤化学性质
树木生长需要从土壤中不断吸取养分,并且需要适宜的土壤酸碱度。从表4可知,尾细桉不同龄组土壤化学性质各指标差异性均显著(P<0.05)。随着林分的生长发育,土壤有机质含量和pH值变化趋势一致,均呈下降的趋势,而在过熟林II阶段时又有所增大。全氮、全磷、有效磷含量随着林分的生长发育呈明显的波动。全钾、碱解氮和速效钾含量的变化趋势一致,均为先降低后增加。过熟林II的全氮、全钾、全磷含量均最高。
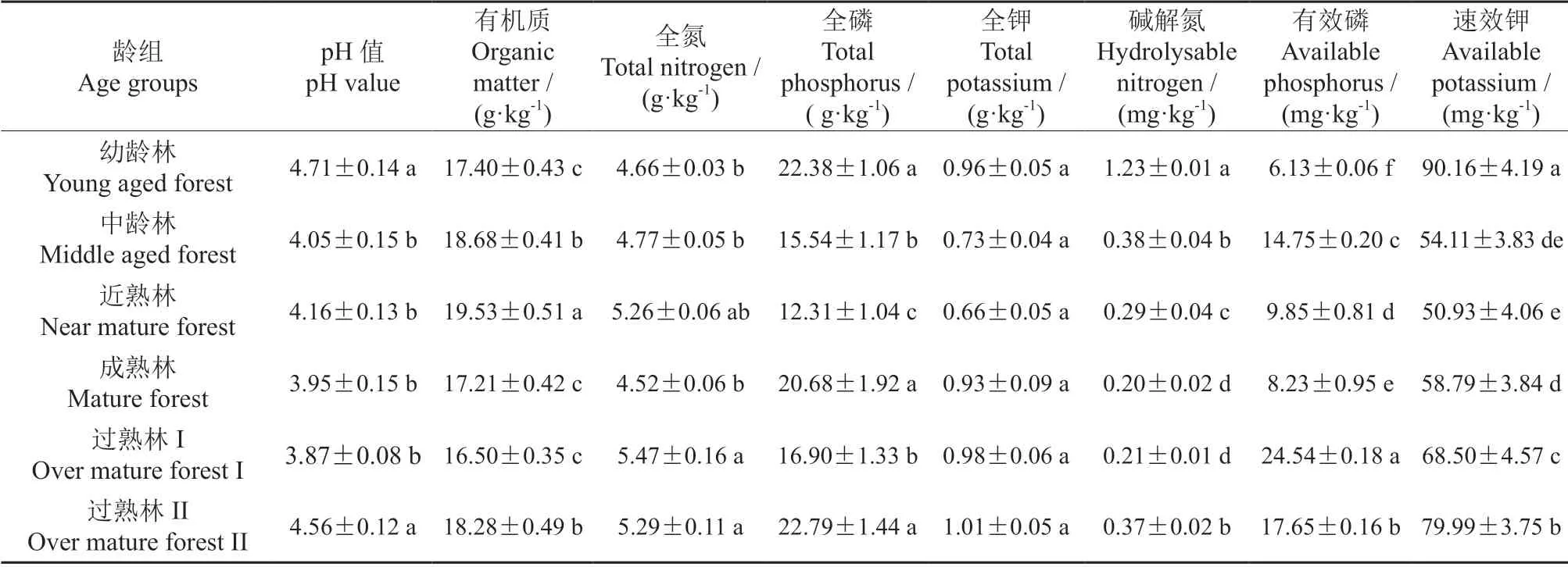
表4 不同龄组尾细桉人工林土壤化学性质Table 4 Chemical properties of soil in E.urophylla×E.tereticornis at different age groups
3.3 不同龄组尾细桉人工林林下植物组成及多样性
3.3.1 尾细桉人工林林下物种组成
在调查的所有样方中,6个龄组的尾细桉人工林林下植物种类为82科196属234种,其中灌木植物共118种,草本植物共116种。其中禾本科Poaceae植物最多,占19属23种;其次是菊科Compositae植物,共计15属18种;再次是豆科Leguminosae植物,为13属15种。分别对其林下灌木层和草本层的相对重要值进行统计,限于篇幅,仅列出各龄组植物优势种及其相对重要值(表5)。结果(表5)表明:在草本层中,幼龄林优势种为阔叶丰花草、芒和飞机草,中龄林优势种为含羞草、耳草和飞机草,近熟林优势种为牛筋草、飞机草和含羞草,成熟林优势种为芒、牛筋草、狗尾草和阔叶丰花草,过熟林I优势种为阔叶丰花草、含羞草、芒和飞机草,过熟林II优势种为牛筋草和海金沙,表明飞机草在尾细桉所有龄组中均为优势种,牛筋草和海金沙随着林分生长发育的进行逐渐成为林地的优势种。在灌木层中,地桃花和白背叶为幼龄林的优势种,地桃花和野牡丹为中龄林的优势种,银柴和车桑子为近熟林的优势种,白背叶为成熟林的优势种,银柴为过熟林I优势种,簕欓花椒为过熟林II优势种。

表5 尾细桉人工林林下植物物种组成Table 5 Species composition of the understory vegetation under of E.urophylla×E.tereticornis plantations
3.3.2 尾细桉人工林林下植物多样性
将不同龄组尾细桉林下灌木和草本的丰富度指数、辛普森(Sinpson)多样性指数、香农-威纳(Shannon-Wiener)多样性指数和均匀度(Peilou)指数分别按不同龄组分组并进行单因素方差分析,结果(表6)表明,除灌木层辛普森(Sinpson)多样性指数外,其余多样性指标差异性均达到了显著水平(P<0.05),表明林分龄组对尾细桉林下灌草多样性具有显著影响。

表6 不同龄组尾细桉林下灌草多样性Table 6 Species diversities of the understory shrub and herbage layers relative to different age groups of E.urophylla×E.tereticornis plantations
对不同龄组的各植物多样性指数进行分析,结果(图1)表明,随着林分生长的进行,灌木层和草本层物种丰富度指数变化趋势表现不一致,灌木层整体上呈增加的趋势,草本层整体上呈减少的趋势。在成熟林期,灌木层丰富度指数增幅较大,其值达到最高,而草本层丰富度指数降幅明显,出现低于灌木层的现象,而在林分生长过程的其它阶段,草本植物均多于灌木植物。灌木和草本的辛普森指数均在近熟林时期最高,草本辛普森指数随着生长的进行呈先升高后降低的趋势。灌木和草本的香农-威纳指数随着生长的进行基本上呈先升高后降低的趋势,其值均在近熟林时期达到最大。灌木和草本的均匀度指数随着生长的进行基本呈先升高后降低再升高的趋势。分析可知,各多样性指数随着生长的进行基本上都呈先升高后降低再升高的趋势,且各指数均在近熟林时期达到最高。

图1 不同龄组尾细桉人工林植物多样性指数Fig.1 Vegetation diversity of E.urophylla×E.tereticornis plantations in different age groups
3.4 不同龄组尾细桉林木生长、土壤理化性质、林下植物多样性的相关性
3.4.1 尾细桉林木生长指标与土壤理化性质的相关性
以尾细桉6个龄组中胸径、树高、材积以及优势木的树高和胸径等生长指标为因变量,以土壤理化性质含水量、容重、孔隙度、pH值、有机质、养分等指标为自变量,分析二者的相关性,结果见表7。研究发现,尾细桉胸径与土壤全磷含量呈显著正相关(P<0.05),虽然与土壤其它理化指标相关性未达到显著水平,但与全钾含量、速效钾含量、有机质含量相关性较高。树高与土壤有机质含量呈显著负相关(P<0.05),与全磷含量、全钾含量、速效钾含量相关性较高。材积与土壤理化性质各指标在统计学上均未达到显著水平(P>0.05),但与有机质含量、碱解氮含量相关性较高。优势木树高与土壤全钾含量呈显著性正相关(P<0.05),与速效钾含量相关性也较高。除了全氮和全钾含量,优势木胸径与有效磷含量呈显著正相关(P<0.05),与其它指标呈极显著负相关(P<0.01)。综合分析,尾细桉生长指标与土壤有机质含量及土壤养分含量的相关性高于土壤物理性质和土壤pH值。

表7 尾细桉生长性状与土壤理化性质的相关系数†Table 7 Correlation coefficient between growth trait and soil physiochemical properties in E.urophylla×E.tereticornis plantations
3.4.2 尾细桉林木生长指标与林下植物多样性的相关性
对尾细桉生长指标和林下植物多样性进行相关性分析,结果(表8)发现,尾细桉人工林林下草本丰富度指标对其胸径、树高、优势木树高、优势木胸径具有极显著的负相关关系(P<0.01),与材积指标呈显著负相关(P<0.05),尾细桉生长指标与其它林下植物多样性指标相关性不显著。
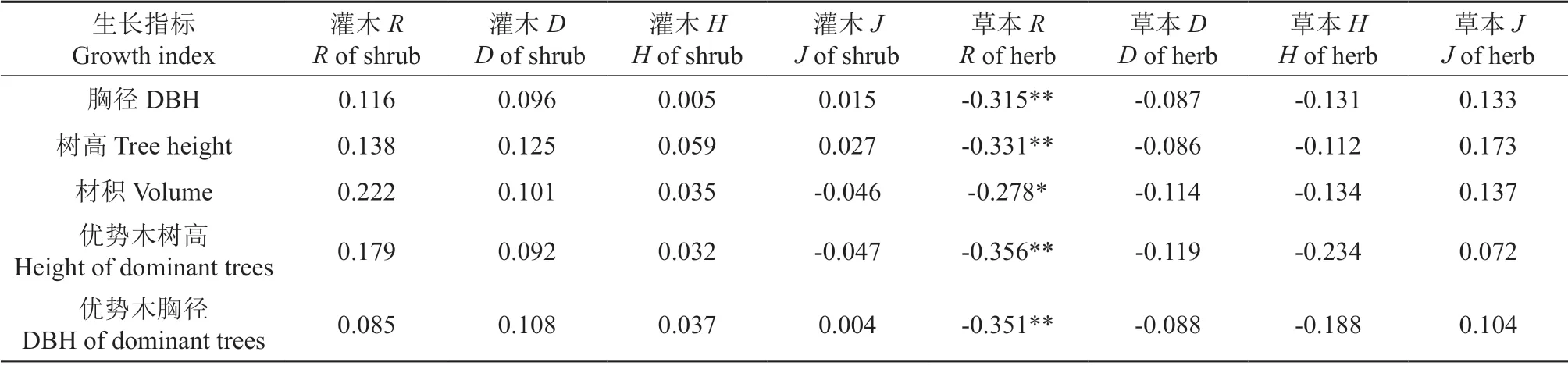
表8 尾细桉生长性状与林下植物多样性的相关系数Table 8 Correlation coefficient between growth trait and diversity indices in E.urophylla×E.tereticornis plantations
3.4.3 尾细桉林下植物多样性与土壤理化性质的相关性
土壤理化性质和桉树林下植物多样性并非相互独立,二者是相互关联和影响的,因此,对二者进行相关性分析。研究结果(表9)表明,灌木层物种丰富度与土壤容重、土壤全钾含量和速效钾含量具有显著相关性(P<0.05),灌木层辛普森多样性指数与土壤理化性质相关性不显著,灌木层香农-威纳多样性指数与土壤速效钾含量、均匀度指数和土壤容重呈显著正相关(P<0.05);草本层丰富度指数与土壤有机质含量呈显著正相关(P<0.05),草本辛普森多样性指数和香农-威纳多样性指数与土壤容重均呈极显著正相关(P<0.01),与土壤总孔隙度均呈显著负相关(P<0.05),草本层香农-威纳多样性指数与空气孔隙度呈显著负相关(P<0.05);草本均匀度指数与土壤各理化性质指标相关性不显著。进一步表明,林下植物多样性对土壤结构及其物理性质的影响大于土壤化学性质的影响。

表9 尾细桉林下植物多样性与土壤理化性质的相关系数Table 9 Correlation coefficient between vegetation diversity indices and soil physiochemical properties in E.urophylla×E.tereticornis plantations
4 讨 论
林木生长、植物多样性与森林土壤养分之间的关系一直是森林经营者重点关注的问题之一。通过分析植物多样性的差异以及土壤理化性质对林木生长的影响,获得影响林木生长的主要环境因子,对于实现森林的可持续经营与管理具有重要意义[26-27]。本研究发现,随着林分生长发育的进行,尾细桉林木生长量及优势木生长量呈显著增加的趋势,由于桉树属于速生丰产树种,轮伐期为5~7 a,在前5年生长速度较快,进入过熟林时期生长量缓慢增加,为了解该地区桉树林木生长状况及实现可持续经营与管理奠定了理论基础。
土壤理化性质是控制植物生长发育的关键生态因子[28],其中土壤容重和孔隙度影响土壤养分的有效性和植物生长[29-30]。本研究发现不同龄组对土壤容重和孔隙度均产生了显著影响。土壤容重综合反映土壤结构及物理性质,腐殖质多,疏松多孔,容重越小,土壤越紧实,容重越大[31]。随着林分生长发育的进行,尾细桉人工林土壤容重呈先升高后下降的趋势,在近熟林阶段其值达到最高,这与林下灌木和草本多样性指标变化趋势基本一致,在近熟林阶段林下植物多样性最大,可能是土壤的疏松度和腐殖质含量间接影响了林下植物多样性。土壤的化学特征是表征土壤肥力的重要指标,直接影响着植物的生长发育[32-34]。本研究中尾细桉不同龄组土壤化学性质各指标差异性均显著,在尾细桉幼龄林阶段土壤pH值最高,随着生长发育的递进土壤呈现酸化趋势,原因可能是植物根系对离子的吸附作用而释放H+,加上有机质的积累和土壤微生物的呼吸作用等因素,导致土壤酸度增加[35]。在15年生过熟林阶段,土壤养分相对较高,形成的原因可能是15年生过熟林植物群落趋于稳定,灌木、草本层植物和桉树形成了复合生态系统,灌木、草本的枯枝落叶和植物残体在总的凋落物组分中所占比例较高,造成了该阶段土壤养分的积累。
林龄反映了森林群落的完整性和演替的进程,不同龄组其林下植物的多样性会有所差异。本研究发现不同龄组的尾细桉人工林林下植物多样性差异显著,林下物种丰富,出现的植物种类共计82科196属234种,灌木植物共118种,草本植物共116种。相比广西、广东部分地区桉树林下植物的种类[36-39],海南桉树人工林林下植物更为丰富,这主要是由于海南属热带季风气候,雨量充沛,长夏无冬,有利于各种植物的生长发育。本研究表明,各龄组尾细桉林下灌木层和草本层物种丰富度和多样性差异显著,林下植物多样性指数随着林分发育的进行基本上呈先升高后降低再升高的趋势,各指数基本上在近熟林时期达到最高。其中草本层的物种丰富度和多样性随着林分发育的进行波动较大,原因是草本层植物生长周期短,更易受到环境因子的影响[27]。各指数在前期阶段变化较大,主要是因为林分成熟前光照条件较好,郁闭度较小,各物种竞争剧烈,在尾细桉过渡到成熟林和过熟林阶段,林分郁闭度较高,林下植物竞争趋于稳定,其物种丰富度和多样性变化相比幼龄阶段较小。这和一些经典的理论模型结果一致,均认为林下植物的丰富度在幼林龄阶段趋于最大,随着林龄的增加物种丰富度随之减少[40]。
植被群落的地上生物量和物种多样性与土壤理化性质(尤其是水肥条件)是相互关联、相互制约的[41]。本研究中,除尾细桉优势木胸径指标与土壤理化性质相关性较强外,其余生长指标与其相关性较弱。林分树高与土壤有机质相关性显著,胸径与全磷含量相关性显著,优势木树高与全钾含量相关性显著。虽然有些土壤理化性质指标与尾细桉生长量指标在统计学上没有达到显著水平,但一些指标对尾细桉生长量指标影响较大,如全磷含量、全钾含量、速效磷含量与生长指标相关性均较强,所以土壤中磷和钾的含量能影响尾细桉生长状况,在经营管理中要注意磷肥和钾肥的施用。桉树植物群落的生长状况与土壤养分尤其是磷、钾含量关系密切,土壤养分含量的差异性会在桉树林木以及林下植被的生长状况及多样性上体现出来,植被生长状况及多样性影响土壤养分积累[42]。尾细桉人工林林下草本丰富度指标与其生长指标如胸径、树高、优势木树高、优势木胸径具有极显著的负相关关系,与材积指标也呈显著负相关,由此可知林下草本植物数量的增加影响了桉树林木的生长,所以在经营管理中要加强除草等抚育工作。本研究表明,灌木层和草本层物种多样性指数与土壤理化性质指标之间也存在一定的相关性,其中灌木物种丰富度指数与土壤容重、全钾含量、速效钾含量具有显著相关性,灌木均匀度指数与土壤容重具有显著相关性,而且尾细桉林木生长与土壤钾含量也具有一定的相关性,说明土壤钾肥含量能显著影响桉树林地植物和林木的生长。相比灌木植物,草本植物更易受土壤理化性质的影响,尤其更易受土壤物理性质的影响,这说明草本植物对环境反应敏感,随机性强,易受到不同状况的干扰。土壤养分与草本层植物相关性不显著,与杨再鸿等[17]的研究结果一致,而全钾和速效钾含量对灌木植物的生长和分布密切相关,这与前者研究略有不同,原因可能是由取样和桉树龄级不同造成的。
海南具有典型的环境气候特点,由于本研究在分析林木生长、林下植物与林地土壤的关系时,未考虑气候因子和土壤微生物因素的影响,研究尺度小,这是本研究的局限性。以后在综合各立地因子对桉树林生态群落多时空大尺度上的动态变化影响有待进一步研究,对提高林地生产力和生态群落变化的综合评价也是今后研究的方向。
5 结 论
本研究通过对海南大面积种植的不同龄组尾细桉纸浆林的林木生长、土壤理化性质和林下植物多样性的相关分析,明确了该地区桉树林生长发育全过程中植物多样性和土壤特征关系的动态变化趋势和规律。研究发现土壤养分磷和钾的含量以及容重对尾细桉人工林林木生长和植物多样性具有较大影响,林木与林下草本植物存在明显的竞争关系,林木和植被在维护林地土壤质量及促进养分循环中起着重要作用,通过相互作用提高了生态系统的稳定性。因此,在桉树人工林的经营管理中,为促进桉树生长和维护植物多样性,要注意钾肥和磷肥的施用,同时为了更好地维持森林群落的物种多样性和稳定性,可适当延长轮伐期,以促使桉树人工林走上可持续发展的道路。
猜你喜欢
乡村科技(2021年1期)2021-12-03
农村实用技术(2021年6期)2021-12-01
疯狂英语·新阅版(2021年7期)2021-08-27
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
东坡赤壁诗词(2020年2期)2020-06-04
教育家(2020年14期)2020-04-26
农民致富之友(2020年4期)2020-04-03
幼儿画刊(2018年1期)2018-01-04
农业与技术(2017年9期)2017-05-28
