
巨菌草根际土壤微生物群落代谢功能多样性分析
2021-05-30 13:26:56翁志强许钰滢吕正阳蔡杨星王荣波叶文雨
广东农业科学 2021年4期
翁志强,许钰滢,吕正阳,刘 娟,王 哲,蔡杨星,王荣波,叶文雨
(1.福建农林大学国家菌草工程技术研究中心/福建农林大学生命科学学院,福建 福州 350002;2.福建省农业科学院植物保护研究所,福建 福州 350002)
【研究意义】土壤微生物是土壤生态系统的分解者和土壤有机质循环的主要动力,它参与土壤养分转化和循环过程,是土壤生态系统的重要组成部分,在植物养分供应和生长过程中发挥重要的作用[1-3]。土壤微生物分解有机质,增加土壤养分,为植物的生长提供养分。同时,土壤微生物对土壤理化性质也有一定的影响。所以土壤微生物的数量种类与土壤肥力的高低具有一定的联系[4-5]。巨菌草(Pennisetum giganteumJUJUNCAO)隶属禾本科狼尾草属(Pennisetum),是福建农林大学林占熺教授引进并选育的一类草本植物,巨菌草是致富草、治沙草、脱贫草、友谊草,已经帮助一些非洲国家突破了农业发展瓶颈。巨菌草具有广泛用途,如食药用菌栽培、饲养牛羊、防风固沙、生物质能源制备、造纸等,对其根际土壤展开土壤微生物群落的功能多样性的研究,有助于对其进行更为合理的种植与利用[6-12]。【前人研究进展】土壤微生物群落在植物的生长过程中发挥着重要的作用,土壤微生物促进植物的生长,植物为土壤微生物提供营养。Biolog-ECO 微平板具有31 种碳源,96 孔板3 次重复,根据微生物对不同碳源的利用程度来研究土壤微生物的功能多样性。Biolog 技术是研究土壤微生物群落的功能多样性的简单、快速方法之一,越来越多的科研工作者使用该技术。目前,Biolog 技术已经用于植物、果园、油田、矿区等林业、花卉等根际土壤微生物的研究。前人对巨菌草进行了栽培等方面的研究,对巨菌草根际土壤微生物的研究尚未见文献报道[13-15]。【本研究切入点】将巨菌草根际土作为研究对象,开展其微生物群落功能多样性的研究,探讨适宜巨菌草自然生长的立地条件,为巨菌草及其他植物的生长提供理论依据,以期为更好地经营管理植物提供新的思路和基础理论基础。【拟解决的关键问题】本研究通过对平潭试验基地巨菌草根际土壤与非根际土壤不同碳源利用,揭示根际土壤微生物群落功能的多样性,促进巨菌草产业化种植,保护生态,同时为巨菌草生态治理应用提供参考。
1 材料与方法
1.1 试验材料
供试土壤样品采自福州平潭实验基地巨菌草根际土与非根际土壤。土壤为砂质土壤,保水性能差,保肥性能弱。
主要仪器设备:酶联免疫检测仪SpectraMaxi3(美谷分子仪器(上海)有限公司)、八通道移液器(Eppendorf AG)、Biolog-Eco 生态板(美国Biolog 公司)、生化培养箱(宁波海螺赛福实验仪器厂)、UNIQUE-R20 纯水机(锐思捷科学仪器有限公司)、超净工作台ZHJH-C1214B(厦门德维科技有限公司)等。
1.2 试验方法
1.2.1 土壤样品的采集 在每块样地内以S 型采样法选取5 个采样点,用土钻采集根际土和非根际土壤样品,混合后装入已灭菌的土壤袋中,带回实验室后1 d 内进行分析[16]。
1.2.2 巨菌草根际土及其对照土壤微生物群落多样性分析 采用Biolog-ECO 技术对土壤微生物群落功能及代谢多样性进行分析,利用酶标仪在590 nm 波长下进行测定和数据读取,分析其多样性,3 次重复。生态板31 种碳源分为6 类:羧酸类7 种、多聚合物类4 种、糖类10 种、酚类2 种、氨基酸类4 种、胺类2 种。具体方法:称取10 g 新鲜根际土壤,加入盛有100 mL、0.85%NaCl 溶液的三角瓶中,摇床280 r/min,振荡30 min,取出后冰浴静置2 min,用0.85% NaCl溶液梯度稀释至1 000 倍。用移液器将上述稀释液加入Biolog-Eco 微平板中,每孔加150 μL,25 ℃恒温培养,每隔24 h 使用酶标仪读取数据,共记录7 d[17]。
将每次读板得到的吸光值减去对照(培养0 h)孔的吸光值,以去除本底干扰,负值全部归0。分别计算31 个孔中碳源的平均颜色变化率(Average well color development,AWCD)和培养144 h 的数据进行土壤微生物多样性分析。

式中,C 为生态板中微孔的吸光度值(590 nm),R 为生态板中空白对照孔的光密度值,n 为底物数量(Eco 板,n=31)。
Shannon-Wiener 多样性指数、Simpson 优势度指数、Pielou 均匀度指数等采用DPS(7.05)软件进行数据处理与分析。以培养144 h 的数据进行碳源代谢和主成分分析。
2 结果与分析
2.1 土壤微生物群落平均颜色变化率(AWCD)
AWCD 值反映了巨菌草根际土壤样品中微生物群落对碳源利用的整体情况和代谢活性。分别培养巨菌草根际土壤及其对照的微生物在0、24、48、72、96、120、144 h 共7 个时段的碳源利用情况,巨菌草根际土壤及其对照的微生物对碳源利用在144 h 后趋于平缓。从图1 可以看出,无论是巨菌草根际土壤还是其对照土壤,微生物群落在0~48 h 时段对碳源的利用最低,在144 h对碳源的利用趋于平缓,因此选择144 h 的吸光值作为计算微生物群落多样性的指标值。巨菌草根际土壤AWCD 值明显高于对照土样品,说明巨菌草根际土壤微生物对碳源的利用程度大于对照土壤。

图1 巨菌草根际土壤与非根际土壤微生物碳源利用Fig.1 Carbon source utilization of microorganisms in rhizophere and non-rhizosphere soils of Pennisetum giganteum
2.2 巨菌草根际土与非根际土壤微生物群落功能多样性分析
Shannon 多样性指数是均匀度和丰富度的综合指标,Simpson 多样性指数值越大,表示多样性程度越高,Shannon 多样性指数值越大,表示群落多样性越大。由表1 可知,巨菌草根际土壤的Simpson 优势度指数、Shannon-Wiener 多样性指数、Brillouin 多样性指数、Pielou 均匀度指数比非根际土壤略大,表明巨菌草根际土壤微生物的群落多样性程度高。

表1 巨菌草根际土壤及其对照土壤微生物群落多样性指数Table 1 Diversity indexes of microbial communities in rhizophere and control soils of Pennisetum giganteum
2.3 根际土壤微生物群落利用碳源情况
表2 为巨菌草根际土与非根际土壤样品144 h 各类碳源利用情况。巨菌草根际土壤微生物群落不能利用的碳源有L-苏氨酸、2-羟基苯甲酸、α-丁酮酸、D-苹果酸、α-D-乳糖、1-磷酸葡萄糖等碳源,其非根际土壤不能利用的碳源有腐胺、1-磷酸葡萄糖等碳源。

表2 巨菌草根际土壤及其对照土壤微生物对BIOLOG-Eco 板上碳源的利用情况Table 2 Vtilization of Carbon sources on BIOLOG-Eco plate of microorganisms in rhizophere and control soils of Pennisetum giganteum
2.4 土壤微生物群落利用碳源主成分分析
运用SPSS19.0 软件采取降维的方法,基于培养144 h 的AWCD 值进行样品土壤微生物利用碳源的主成分分析。
PCA1 解释变量方差的54.727%,PCA2 解释变量方差的45.273%,说明PCA1 对群落的贡献略大。第一主成分与第二主成分贡献率之和达到100%,说明提取的PCA1 和PCA2 主成分可以反映土壤微生物对6 类碳源的利用情况(表3)。
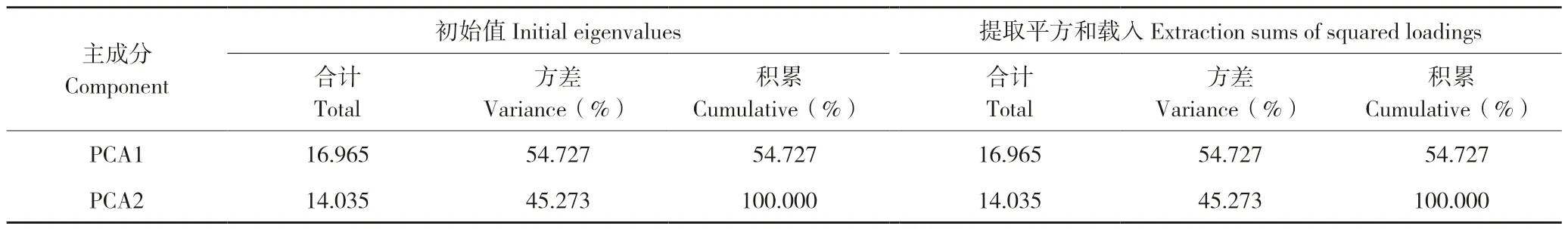
表3 巨菌草根际土壤及其对照土壤微生物群落碳源利用主成分分析Table 3 PCA on carbon source utilization of microbial communities in rhizophere and control soils of Pennisetum giganteum
31 种碳源对PC1 和PC2 贡献的特征向量系数反映了单一碳源对PC1 和PC2 的贡献(图2),由图2 可知,对PC1 贡献大的碳源(系数>0.50)有22 种,其中糖类8 种(β-甲基-D-葡萄糖苷、D,L-α-磷酸甘油、D-半乳糖酸γ-内酯、1-磷酸葡萄糖、α-D-乳糖、D-甘露醇、D-木糖、N-乙酰-D 葡萄糖氨),羧酸类占5 种(D-苹果酸、D-葡糖胺酸、衣康酸、D-半乳糖醛酸、γ-羟丁酸),氨基酸类3 种(L-苯丙氨酸、L-丝氨酸、L-天门冬酰胺),多聚物3 种(吐温40、吐温80、α-环式糊精),胺类2 种(腐胺、苯乙胺),酚酸类1 种(4-羟基苯甲酸)。可见影响PC1 的主要为糖类和羧酸类;对PC2 贡献大的碳源有7 种,其中酚酸类2 种(2-羟基苯甲酸、4-羟基苯甲酸),糖类1 种(D-纤维二糖),氨基酸类3 种(L-苯丙氨酸、L-苏氨酸、L-精氨酸)。可见影响PC2 的主要为氨基酸类。4-羟基苯甲酸和L-苯丙氨酸对PC1 和PC2 贡献系数都>0.50。
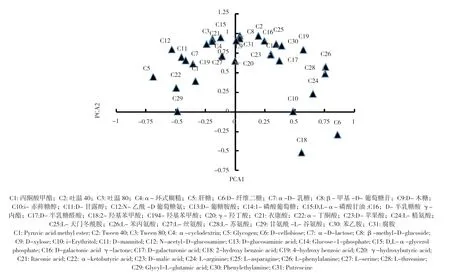
图2 31 种碳源对PC1 和PC2 贡献的特征向量系数Fig.2 Eigenvector coefficients of 31 carbon sources with loadings for PCA1 and PCA2
3 讨论
巨菌草具有独特生态功能,根际微生物群落对其生长发育及环境适应性密切相关。Biolog-Eco技术是研究土壤微生物多样性的技术手段之一,采用该技术研究菌草根际土壤微生物群落功能多样性,有助于探寻巨菌草最适生长条件。环境条件、植被、土壤肥力、物质循环和能量转化都与土壤微生物的活性有极大的相关性,土壤微生物是土壤物质循环的主动力[20-24]。在利用Biolog-Eco生态板技术研究土壤微生物群落代谢活性时,如果 AWCD(平均颜色变化率)值越高,说明土壤微生物群落代谢活性就高,反之,说明其微生物群落代谢活性低[25]。本研究中,巨菌草根际土壤各时段的AWCD 值均高于非根际土壤,这表明巨菌草根际土壤微生物的代谢活性高于非根际土壤,同时也说明对碳源的利用程度也高于非根际土壤。一方面由于巨菌草植株高大,根系发达等特点,为土壤微生物创造了良好的生存环境,有利于土壤微生物群落多样性和功能多样性的形成。但是巨菌草根际土壤微生物活性的提高是否会影响巨菌草及其他植物的生长发育、产量及品质也未做相关研究。哪一类微生物对巨菌草的影响是显著的也是值得进一步的研究。
土壤微生物对不同碳源利用能力的差异可以在一定程度上指示土壤微生物的群落结构组成[26]。本实验碳源利用分析结果表明,巨菌草根际土壤微生物群落能利用六大类(糖类、多聚物类、氨基酸类、羧酸类、胺类和酚酸类)碳源,其生理活性高于非根际土壤,提高了对碳源的利用。这可能是由于巨菌草植株残体为土壤微生物群落提供了丰富的营养,进一步造成了土壤微生物生境不同,进而造成微生物群落利用碳源的不同,这种解释有待于进一步用培养基培养法分离微生物进行验证。
平潭菌草试验基地地处沙漠区,种植菌草后沙漠变绿洲,这在一定程度上改善了当地的生态环境条件。土壤微生物多样性指数比较分析表明,巨菌草根际土壤微生物的Pielou 指数、Shannon指数、Brillouin 指数高于非根际土壤,但差异不显著。这可能是由于植物根系分泌物不同所致。巨菌草根际土微生物群落结构和优势微生物的种类,还有待通过高通量测序技术、PLFA 等技术进行进一步研究。
4 结论
本研究通过Biolog-ECO 技术分析巨菌草根际土壤微生物群落代谢功能多样性,结果表明,巨菌草根际土壤微生物群落具有较高的AWCD 值,巨菌草根际土壤微生物群落的碳源代谢能力大于非根际土壤。巨菌草根际土壤微生物群落的代谢功能强于非根际土壤。L-苏氨酸、2-羟基苯甲酸、α-丁酮酸、D-苹果酸、α-D-乳糖、1-磷酸葡萄糖等碳源巨菌草根际土壤微生物群落不能利用。应用培养基培养法研究土壤微生群落时,应优先考虑不能利用的其他碳源。巨菌草根际土壤微生物的活性及多样性均比非根际土壤高,该研究结果为巨菌草的合理种植、生态治理与土壤改良及提高产量的研究提供理论依据。
猜你喜欢
小学生学习指导(高年级)(2023年10期)2023-12-10 16:50:49
发明与创新·中学生(2023年5期)2023-04-20 01:59:52
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
现代园艺(2017年23期)2018-01-18 06:57:57
中国蔬菜(2016年8期)2017-01-15 14:23:38
足球周刊(2016年15期)2016-11-02 15:46:29
财政监督(2016年21期)2016-03-27 02:48:36
金色年华(2016年8期)2016-02-28 01:39:40
发明与创新(2015年21期)2015-02-27 10:39:09
