
序贯调节巨噬细胞极化的骨修复材料构建及其体外成管化作用研究*
2021-05-29 11:38贺思豪艾秋池周江玲侯天勇邢军超
重庆医学 2021年9期
程 鹏,贺思豪,艾秋池,周江玲,侯天勇,邢军超
(陆军军医大学第一附属医院骨科/全军矫形外科中心/陆军军医大学组织工程研究与开发中心,重庆 400038)
大段骨缺损是临床骨组织损伤救治的一大难题,重要原因之一在于缺乏有效可靠的具有组织重建功能的骨修复材料。既往骨修复材料的研发重点关注其成骨能力,往往忽略血管化这一成骨活动的基础条件[1-2]。有研究表明,移植骨的成骨活动总是与血管化过程密切相关,血管化的程度甚至直接决定了骨移植的成败[3-4]。早期良好的血管化不仅为移植材料输送氧气及营养物质、移除代谢废物,还为宿主成骨相关细胞如间充质干细胞(MSCs)等的迁移定植提供必需的条件[5-6]。在骨缺损缺血区的血管及骨重建过程中,局部炎症因子的协同作用调节的巨噬细胞极化扮演重要角色。巨噬细胞分为经典激活途径巨噬细胞(M1型巨噬细胞)和替代激活途径巨噬细胞(M2型巨噬细胞),在组织修复早期,M1型巨噬细胞发挥促炎功能,在后期M2型巨噬细胞发挥抗炎、促进损伤组织修复和血管生长的作用[7]。为此,本研究利用藻酸盐缓释系统和生物素-亲和素系统耦联脱钙骨基质(DBM)构建骨修复材料,实现干扰素(IFN)-γ和白细胞介素(IL)-4的序贯释放,调节巨噬细胞极化从而促进血管生成,为其在骨缺损修复治疗中的可行性提供实验依据。
1 材料与方法
1.1 材料与试剂
人骨髓来源内皮祖细胞(EPC)购自美国ATCC公司(PCS-800-012),人外周血来源单核巨噬细胞购自美国ATCC公司(CRL-9855);内皮细胞培养基(ECM)、0.25%胰蛋白酶、特优胎牛血清(FBS)、巨噬细胞集落刺激因子(M-CSF)、IMDM培养基、DMEM/F12培养基购自美国HyClone公司;海藻酸钠、Dil-ac-LDL、FITC-UEA-1、Biotin-sulfo-LC-LC-NHS、酶联免疫吸附试验(ELISA)试剂盒、反转录试剂盒购自美国Sigma公司;重组人IL-4、IFN-γ蛋白购自美国PeproTec公司;抑制剂购自美国Selleck公司。
1.2 方法
1.2.1细胞培养
(1)EPC的培养:细胞复苏后加入新鲜ECM,将细胞接种于培养皿中,于5% CO2,37 ℃恒温孵箱孵育24 h后见细胞贴壁生长,换液后以磷酸盐缓冲液(PBS)清洗2次以去除未贴壁细胞,然后加入新鲜培养基,隔天换液至细胞融合达到90%时传代,取P3代对数生长期的细胞使用。(2)巨噬细胞的培养:培养基为含10% FBS的IMDM培养基,细胞解冻、离心、弃上清液后加入新鲜培养基并接种于超低吸附培养瓶(美国Corning)中,于5% CO2,37 ℃恒温孵箱孵育24 h后换液,隔天换液至细胞密度达到80%时传代,传代后培养基中加入10 ng/mL的巨噬细胞克隆集落刺激因子(M-CSF),隔天换液至第5天,取细胞使用。
1.2.2材料构建
将材料分为4组:空白对照组(Ctrl组)、单纯IFN-γ组(IFN组)、单纯IL-4组(IL-4组)、IL-4加IFN-γ组(联合组)。(1)DBM的制备:按照本课题前述方法制备支架,依托陆军军医大学第一附属医院骨库,以云南小香猪胫骨、股骨近远端松质骨块,制备支架材料。在密闭玻璃容器中加入 0.6 mol/L盐酸,支架投入其中浸泡脱钙。常温浸泡 24 h,每8小时更换盐酸1次,当皮质骨呈半透明能弯曲的弹性状态,松质骨块呈海绵状压缩后自动恢复原状,将骨块取出,以无菌蒸馏水反复冲洗、浸泡,直至pH值为7,吸出浸泡液将DBM支架材料置于离心管中,300×g 离心 5 min,对DBM进行辐照处理以灭菌,真空冷冻干燥机干燥后分装备用。(2)DBM的生物素化:取 DBM支架于PBS浸泡后修剪大小约3 mm×3 mm×3 mm,将其浸泡于10 mmol/L Biotin-sulfo-LC-LC-NHS中1 h,PBS冲洗3次以清除未附着的生物素,70%乙醇溶液浸泡消毒10 min,以PBS洗净后浸泡在PBS中,4 ℃浸泡 24 h。(3)蛋白生物素化:调整重组人IL-4蛋白溶液浓度为1 mg/mL,向其中加入10 mmol/L Sulfo-NHS-Biotin粉末,室温放置1 h完成蛋白生物素化,PBS透析移除未附着的生物素,无菌过滤后存于4 ℃备用。(4)支架-蛋白耦联:将生物素化的DBM支架浸泡于172 μg/mL的亲和素溶液中1 h,PBS冲洗3次,再将IL-4组与联合组浸泡于生物素化的IL-4溶液1 h,IFN组和Ctrl组浸泡于PBS溶液中1 h,PBS冲洗3次。由于亲和素可提供4个高亲和力的生物素结合位点供其特异性结合,使得IL-4与支架之间的非共价连接十分牢固。(5)复合材料构建:制备含重组人IFN-γ蛋白的海藻酸钠溶液(IFN-γ 1 mg/mL,海藻酸钠80 mg/mL),以其浸泡IL-4耦联的支架材料30 min,Ctrl组与IL-4组浸泡在不含IFN-γ的海藻酸钠溶液中30 min,浸泡于10.1 mg/mL氯化钙溶液中10 min,重复以上步骤3次,见材料表面有浅白色絮状物附着,去离子水洗净后-80 ℃冻存干燥备用,电镜下观察材料的微观结构。
1.2.3缓释动力检测
取联合组复合材料放置在10 mL的DMEM/F12培养基中,放置在37 ℃恒温箱中,每天更换液体并收集培养基,以10 000 r/min离心10 min,取上清液通过ELISA试剂盒测定IL-4与IFN-γ水平,计算累计释放量。
1.2.4复合材料对巨噬细胞极化的影响
取M-CSF刺激5 d的巨噬细胞,以1×106/mL的浓度采用双面静置接种法接种于各组材料,加入培养基,培育3 d后更换培养基。分别在培养1、4、7 d收集培养基,通过ELISA检测其中蛋白水平。相同时间点取各组材料,液氮研磨后提取mRNA,按反转录试剂盒说明书进行逆转录后行实时定量PCR(RT-PCR)。肿瘤坏死因子α(TNF-α)、趋化因子受体7(CCR7)、趋化因子配体18(CCL18)、血小板衍生生长因子(PDGF-BB)、血管内皮生长因子(VEGF)等引物序列见表1。扩增条件:95 ℃ 预变性30 s;95 ℃ 变性5 s,57 ℃ 退火20 s,72 ℃ 延伸15 s,40 个循环。

表1 RT-PCR引物序列(5′-3′)
1.2.5Transwell实验
取EPC以ECM重悬制备单细胞悬液,调整细胞浓度为1×104个/mL。在24孔板中组装Transwell小室,于下室按照实验分组,分别加入700 μL各组培育3 d后的培养基。按照实验分组于Transwell上室中分别加入200 μL含单细胞悬液、含Axinitib(VEGFR2抑制剂)或含SB225002(IL-8抑制剂)预处理EPC的单细胞悬液。将Transwell小室置于5%CO2、37 ℃的孵箱内,迁移6 h后终止。以无菌棉签擦拭碳酸酯膜上层以去除未迁移的细胞。用PBS冲洗碳酸酯膜3次。以4%的多聚甲醛固定15 min,PBS冲洗3次。0.5%Triton-X100破膜10 min,PBS冲洗3次后以DAPI工作液(1 μg/mL)避光染色5~8 min,PBS冲洗3次。荧光显微镜下每个碳酸酯膜随机选取10个高倍视野(×200)分别计数,计算并比较每组细胞迁移数量。
1.2.6管腔形成实验
将Matrigel胶置于4 ℃过夜使其缓慢融化,加入至96孔板中(50 μL/孔),37 ℃的孵箱内静置30 min,以各组孵育7 d的培养基制备浓度为2×105/mL的EPC单细胞悬液,每孔加入50 μL细胞悬液,37 ℃孵箱内孵育8 h后镜检。
1.3 统计学处理
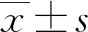
2 结 果
2.1 材料性状
DBM外观如图1A所示,IFN-γ藻酸盐混合溶液与氯化钙溶液反复浸泡后可在支架孔隙表面形成浅白色胶状物(图1B)。扫描电镜结果显示Ctrl组支架孔隙率均一,微孔表面较为光滑(图1C),IL-4、IFN组可见蛋白基质均匀皱缩分布在支架表面(图1D、E),联合组由于IFN-γ藻酸盐混合溶液与氯化钙溶液的反应形成凝胶样褶皱,见图1F。

A、B:支架大体观;C:Ctrl组;D:IL-4组;E:IFN组;F:联合组。
2.2 复合材料的缓释动力
复合材料与IFN-γ和IL-4的结合稳定。复合材料通过藻酸盐缓释系统实现与IFN-γ外层负载达到早期释放的效果,IFN-γ在前4 d稳定快速释放,4 d内释放接近完全;复合材料通过生物素-亲和素系统实现IL-4的内层耦联达到缓释效果,IL-4在前3 d无明显释放,在第4~7天快速稳定释放,此后每天少量释放,至第13天释放完全,见图2。

图2 IFN-γ及IL-4缓释动力(n=5)
2.3 复合材料对巨噬细胞极化状态的影响
RT-PCR结果显示,IFN-γ早期的释放使联合组与IFN组巨噬细胞向M1型极化,在培育第1天,IFN-γ可诱导巨噬细胞释放M1型标志物TNF-α,表现出M1型极化的部分趋势;第4天,IFN-γ的释放使得巨噬细胞表达M1型标志物CCR7、TNF-α及IL-1β;第7天,M1型标志物表达水平相比Ctrl组无明显差异。在第1、4天,耦联炎症因子的3组中细胞均高表达IL-8,在第7天,IL-8的表达在各组间无明显差异。IL-4在第4~7天的稳定释放使IL-4组巨噬细胞向M2型极化,在培育第1天,IL-4组细胞即高表达M2型标志物CD206,证实IL-4可诱导M2型细胞极化;在第4天,IL-4组细胞CD206表达明显升高,而联合组与Ctrl组无明显差异;在第7天,IL-4组细胞CD206与Ctrl组无明显差异,而联合组CD206表达明显升高;在第4、7天,联合组由于IL-4的释放导致M2型标志物CCL18及PDGF-BB表达水平明显升高。对于VEGF表达而言,联合组细胞在第1天表达VEGF,明显高于IL-4组,与IFN组水平相当,联合组细胞VEGF表达水平在第4天达到高峰,在第7天仍高表达,且明显高于其他3组,见图3。ELISA结果显示,在培育第1天和第4天,IFN-γ的释放导致巨噬细胞高表达M1型标志物TNF-α、IL-1β及趋化因子IL-8;IL-4的释放导致巨噬细胞高表达M2型标志物CCL18、PDGF-BB;在第4天和第7天,由于IL-4的释放,复合材料明显促进巨噬细胞的M2型极化标志物的分泌。对于VEGF的表达变化趋势,ELISA结果与PCR结果较为一致,见图4。

a:P<0.05,与Ctrl组比较;b:P<0.05,与IL-4组比较;c:P<0.05,与IFN组比较;d:P<0.05,与联合组比较。
2.4 复合材料对血管生成的影响
管腔形成实验结果显示,IL-4及联合组条件培养基可显著促进管腔形成;阻断VEGF受体(Axitinib)均可明显抑制联合组条件培养基的促血管生成作用,阻断IL-8(SB225002)对联合组条件培养基的促血管生成作用影响甚微,见图5。
2.5 复合材料对EPC迁移的影响
Transwell细胞迁移结果显示,IFN组及联合组条件培养基可明显促进EPC迁移;尽管联合组VEGF表达量较高,阻断VEGF受体(Axitinib)后未见EPC的迁移发生明显改变;而阻断IL-8(SB225002)可明显抑制联合组条件培养基对EPC的募集作用,见图6。

a:P<0.05,与Ctrl组比较;b:P<0.05,与IL-4组比较;c:P<0.05,与IFN组比较;d:P<0.05,与联合组比较。
图4 巨噬细胞蛋白表达变化(n=5)

A:Ctrl组;B:IFN组;C:IL-4组;D:联合组;E:联合组+Axitinib组;F:联合组+SB225002。

A:Ctrl组;B:IFN组;C:IL-4组;D:联合组;E:联合组+Axitinib组;F:联合组+SB225002;a:P<0.05,与Ctrl组比较。
3 讨 论
巨噬细胞是机体的重要免疫细胞,具有吞噬病原、抗原递呈和分泌多种细胞因子等功能。除了在免疫反应过程中发挥重要作用,巨噬细胞在血管生成及骨生成过程中也扮演着不可或缺的角色[8-9]。研究表明,当宿主巨噬细胞缺失时,移植物内的血管生成无法进行,而充分的血管发生对于骨的发育及修复必不可少[10-11]。在不同的组织微环境、细胞因子作用下,根据细胞表面标识性分子及功能的不同,巨噬细胞可极化成M1和M2型。在IFN-γ为代表的众多因子的作用下,巨噬细胞极化为M1型,而在IL-4等因子作用下,其向M2型极化。在组织损伤修复过程中,早期损伤局部产生的炎症因子导致巨噬细胞以M1型为主,尔后M2型为巨噬细胞的主要存在形式[12-13]。基于巨噬细胞在免疫防御、血管生成、骨生成方面的重要作用及其不同极化状态下的不同生理功能,通过移植材料调节宿主巨噬细胞极化状态,从而改善其移植环境内血管化水平的促组织修复方式极有可能更适合于临床大段骨缺损的救治。因此,骨移植材料如能顺序、可控地激活M1及M2型巨噬细胞,将充分调动宿主自身的抗感染及组织损伤后修复能力,这不仅符合当今医学界推崇的原位组织工程理念,还将为临床大段骨缺损的治疗提供新的思路。
为此,本研究利用藻酸盐缓释系统和链霉素-亲和素系统耦联DBM构建了复合材料。藻酸盐是在海藻中提取的天然多糖碳水化合物,为一种天然纤维素,无毒副作用,来源丰富、价格低廉,且具有良好的生物相容性和可降解性[14]。藻酸盐在钙离子溶液中可即刻形成热稳定凝胶,其作为控释系统已被广泛应用[15]。亲和素与生物素有着极强的亲和性,此外还能与一些含生物素的大分子蛋白结合,因此,生物素化的DBM支架和IL-4可由亲和素提供过高亲和力结合位点实现较为牢固的非共价连接。结果显示,复合材料可实现IFN-γ与IL-4的序贯释放。既往有学者采用气体发泡-粒子沥滤的方法制备了多聚-L-谷氨酸(PLG)支架,实现了促血管生成因子如VEGF及促成熟因子PDGF-BB的序贯释放,成功地改善了支架的血管生成能力,并揭示了细胞因子有序作用的重要性[16]。血管生成过程复杂,对众多生长因子和信号分子有着严格的时间和剂量要求,使得针对单一或少数细胞因子的控释产生的作用存疑[17]。而巨噬细胞主要通过分泌因子的方式调控血管生成,其对成骨相关细胞如EPC、MSC、成骨细胞等的生物学行为均有着重要的调控作用。因此,通过材料序贯调控巨噬细胞的分泌功能可能对血管生成及骨修复形成级联放大的调控作用。本研究结果证实,复合材料可在序贯释放细胞因子的基础上实现巨噬细胞极化状态的序贯调节,即在4 d内,巨噬细胞以M1型为主,而后以M2型为主;体外实验结果与组织损伤修复过程中巨噬细胞的极化状态变化时间较为吻合,亦体现了材料的仿生学特征。
在血管生成的过程中,EPC的迁移是关键环节。在组织损伤信号的引导下,EPC由骨髓动员至外周循环,再迁移至损伤部位促进局部新血管的形成,此外EPC已被证实在多种动物模型中具有促进成骨细胞形成从而促进骨修复的作用[5]。本研究结果发现,复合材料可明显促进EPC向巨噬细胞的迁移,提示复合材料具有良好的促血管生成作用。EPCs的迁移受到多条信号转导通路的调节,比如PI3K/Akt,SDF-1/CXCR4/eNOS等,本课题组前期研究证实,在骨缺损模型中,EPC向移植物迁移的主要信号途径为CXCR2-Src-PKL/Vav2-Rac1通路[5]。IL-8是CXCR2的一种高亲和力配体,最初被描述为一种炎性趋化因子,可以趋化白细胞。有研究表明,IL-8可与G-CSF协同作用,通过结合CXCR1和CXCR2来调节EPC的动员进而促进骨的修复[3,18]。SB225002为CXCR2的高选择性抑制剂,在本研究中,发现复合材料对EPC向巨噬细胞迁移的促进作用可被SB225002阻断,提示CXCR2及其配体在其中的关键作用,也从另一方面论证了通过调控巨噬细胞旁分泌功能促进血管生成的可行性。除了通过分泌作用调控成骨细胞形成等过程外,EPC促进血管生成的主要方式在于分化为内皮细胞,直接促进血管再生。EPC通过向内皮细胞分化及促进成骨细胞形成进而促进血管化和骨修复。本研究中,观察到联合组条件培养基可明显促进血管生成,而阻断VEGFR2可明显抑制此作用,说明了VEGF在巨噬细胞调节血管生成过程中的重要作用;而VEGF在联合组持续高表达,亦反证了复合材料良好的促血管生成能力。此外,在众多的细胞因子中,VEGF和PDGF-BB的促血管生成作用最具代表性,VEGF的主要作用为募集内皮细胞启动血管生成,而PDGF-BB主要参与血管生成后期管腔形成及周细胞募集过程,二者在血管生成过程中相继大量表达从而使血管生成过程有序进行[19-20]。本研究制备的复合材料通过调控巨噬细胞由M1向M2的极化,使得VEGF和PDGF-BB的表达高峰可以相继出现。
综上所述,血管化是骨移植成功的关键,而炎性反应在其中起重要作用。如果骨缺损局部的炎症微环境可以通过材料的制备技术和工艺加以合理控制,那么未来的材料将有希望彻底摆脱细胞的束缚、借助宿主的自我修复功能完成重建。为此,本研究构建了可序贯调节巨噬细胞极化的复合材料,通过体外实验初步证实了其促血管生成能力,可为骨缺损的治疗提供新的思路,在骨缺损的应用及研究中具有一定的潜在应用价值。
猜你喜欢
现代财经-天津财经大学学报(2022年5期)2022-06-01
航天电子对抗(2022年2期)2022-05-24
建材发展导向(2022年2期)2022-03-08
兽医导刊(2020年23期)2020-12-30
纤维复合材料(2018年3期)2018-04-25
农村青少年科学探究(2017年6期)2017-09-11
现代检验医学杂志(2016年4期)2016-11-15
合成化学(2015年4期)2016-01-17
电源技术(2015年1期)2015-08-22
应用化工(2014年10期)2014-08-16
