
靶向线粒体治疗脊髓损伤的疗效:基于动物实验的系统评价
2021-05-28 08:10:12王丽群庞日朝刘建成张安仁
中国康复理论与实践 2021年5期
王丽群,庞日朝,刘建成,张安仁
1.成都中医药大学养生康复学院,四川成都市 610075;2.西部战区总医院康复医学科,四川成都市 610083;3.同济大学附属上海市第四人民医院康复医学科,上海市200434
线粒体是一种双膜细胞器,通过氧化磷酸化产生细胞的大部分三磷酸腺苷(adenosine triphosphate,ATP)。线粒体的结构包括线粒体外膜、线粒体膜间隙、线粒体内膜和线粒体基质。线粒体外膜是一个磷脂双层,包含电压依赖性阴离子通道,该通道在打开时允许小分子通过,包括离子、二磷酸腺苷和ATP[1]。线粒体内膜的结构更加复杂,包含多个膜内通道,这些通道的开放受到严格控制,其调节电子传输链(elec‑tron transport chain,ETC)以保持ATP 合成所必需的电化学梯度[2]。ETC 由四个膜结合的复合体和两个载体分子(辅酶Q 和细胞色素C)组成。复合体包括:复合体I、复合体Ⅱ、复合体Ⅲ和复合体Ⅳ。线粒体可以充当能量储存器,调节细胞内Ca2+的水平,并在依赖于线粒体的钙缓冲和ATP 产生的神经元中起着重要的作用[3]。
脊髓损伤是脊柱损伤最严重的并发症,往往导致毁灭性和持续性的功能丧失。脊髓损伤具有复杂的病理过程,一般分为原发性损伤和继发性损伤两个阶段。继发性损伤由原发性损伤逐渐发展而来,对脊柱组织产生进一步的化学和机械损伤。脊髓损伤早期继发性反应是去极化和电压依赖性离子通道的打开,导致谷氨酸受体的激活,大大增加谷氨酸的浓度,并产生持续的兴奋性毒性和细胞死亡,坏死细胞中高水平的谷氨酸盐通过增加细胞内Na+和Ca2+的浓度并降低细胞内K+的浓度来改变离子通量[4]。Ca2+积累过多,将触发线粒体膜通透性转换孔(mitochondrial permea‑bility transition pore,mPTP),mPTP 的开放会干扰质子梯度,使ATP 失活,增加水和其他分子进入线粒体基质,导致细胞肿胀并最终死亡[5]。正常情况下,电子从ETC 泄露并与线粒体基质的氧气结合形成活性氧,内源性抗氧化系统可防止活性氧,引起毒性。然而,脊髓损伤可导致ETC受损和能量不足,形成过多活性氧,超过抗氧化系统的可控范围,最终导致氧化损伤和病理水平。活性氧促使强氧化剂过氧亚硝酸盐生成增加,诱导线粒体内一氧化氮合酶的活化,同时还可以触发细胞膜脂质过氧化、蛋白质羰基化和酪氨酸硝化,进而损伤线粒体并造成线粒体功能障碍[6]。另外,线粒体功能障碍还会导致细胞色素C 释放到细胞质中并造成细胞凋亡[7]。中枢神经系统所需的大部分能量由线粒体提供,而神经元的生存则需要足够的能量。因此,脊髓损伤后的线粒体功能障碍可能导致神经元死亡,加剧损伤[8]。
脊髓损伤具有复杂的病理生理过程,目前有众多改善脊髓损伤的治疗方法,包括手术治疗、药物治疗、细胞移植和纳米技术等[9‑12]。然而,目前临床上仍未找到确切、有效的方法[13],无论是传统的手术干预和药物治疗,还是新型的细胞移植和纳米技术,均存在其不可避免的缺陷。在神经元中,线粒体在钙稳态、控制膜兴奋性以及神经传递和可塑性中起着重要作用。由于神经元特别依赖于线粒体的钙缓冲和ATP产生,因此对线粒体缺陷高度敏感。越来越多的证据表明,线粒体功能障碍参与多种疾病的发病机制,尤其是晚发性神经退行性疾病,如帕金森病、阿尔茨海默病和亨廷顿氏病等[14]。脊髓损伤后线粒体维持体内稳态的能力降低,导致ATP 依赖性细胞功能丧失、钙超载、兴奋性中毒和氧化应激的丧失,进而加剧损伤[15]。以上研究表明,线粒体功能障碍与脊髓损伤导致的有害继发性病理生理密切相关。因此,损伤后线粒体功能的恢复可能是治疗脊髓损伤的潜在有效方法。可以通过不同方法直接靶向脊髓损伤后线粒体功能障碍,包括通过使用替代性能源、抗氧化、抗凋亡、抑制mPTP 和增强线粒体生物发生等多种机制靶向线粒体药理学方法(表1)。靶向线粒体治疗脊髓损伤可能改善的功能和相关评定方法或指标见表2。本研究收集国内外已发表的靶向线粒体治疗脊髓损伤动物实验,对靶向线粒体治疗脊髓损伤的作用进行系统综述,为今后临床治疗脊髓损伤提供实验依据和新思路。

表1 靶向线粒体治疗脊髓损伤的作用机制及其相关药物

表2 靶向线粒体治疗脊髓损伤涉及的功能结局和相关评定方法或指标
1 资料与方法
1.1 文献检索
检索PubMed、Web of Science、中国知网和万方数据知识服务平台,检索年限自建库至2021 年2 月。中文检索词:脊髓损伤、线粒体、功能障碍、治疗等;英文检索词:spinal cord injury、mitochondria、dysfunction、treatment等。
1.2 纳入标准
疾病及动物种属:脊髓损伤动物模型,不限制动物种属和造模方法。
干预措施和对照措施:靶向线粒体治疗的药物干预和安慰剂或其他对照。
主要结局指标:脊髓损伤动物的运动功能(BBB评分)、脊髓组织病理学(受损组织和残余组织面积)、抗氧化功能、抗凋亡功能、线粒体生物发生。
1.3 排除标准
①非干预类动物实验;②非对照研究;③数据不完整;④非中、英文;⑤重复发表。
1.4 文献筛选与资料提取
由2 名研究者独立筛选文献、提取资料并交叉核对。如有分歧,则通过讨论或与第三方协商解决。文献筛选时首先阅读文题和摘要,在排除明显不相关的文献后,进一步阅读全文,确定最终纳入的研究。文献筛选流程见图1。

图1 文献筛选流程图
阅读全文后由两名研究者独立提取相关数据并交叉核对。提取以下内容。①纳入研究的基本信息:第一作者、发表年份、样本量、实验动物的种属及模型种类、干预方式等。②结局指标:脊髓损伤动物的运动功能(BBB评分)、脊髓组织病理学(受损组织和残余组织面积)、抗氧化能力、抗凋亡能力、线粒体的生物发生等。
1.5 文献质量评价
偏倚风险评估:采用基于Cochrane偏倚风险评估工具制定的SYRCLE 动物实验偏倚风险评估工具(SYRCLE's risk of bias tool for animal studies)[26],包括6种偏倚类型(选择偏倚、实施偏倚、测量偏倚、失访偏倚、报告偏倚和其他偏倚)和10 个条目,评估结果最终以“是”、“否”和“不确定”表示,其中“是”代表低风险偏倚,“否”代表高风险偏倚,“不确定”代表不确定风险偏倚。
证据质量评估:采用CERQual (Confidence in the Evidence from Reviews of Qualitative research)[27]进行评估。CERQual 工具从4 个方面进行评估:①方法学局限性;②相关性;③一致性;④数据充分性。综合以上4 部分的评价结果对系统评价单个结果进行信度分级(高、中、低、极低)。
1.6 统计学分析
由于纳入的动物实验研究存在临床异质性和方法学异质性,难以进行定量分析,故仅进行定性分析。
2 结果
初步检索得到相关文献757 篇,其中英文653 篇,中文104 篇。经逐层筛选,最终纳入11 篇动物实验,其中中文3篇,英文8篇。文献筛选结果见图1。
2.1 纳入文献的基本特征
纳入的11篇文献[28‑38]的基本特征见表3。
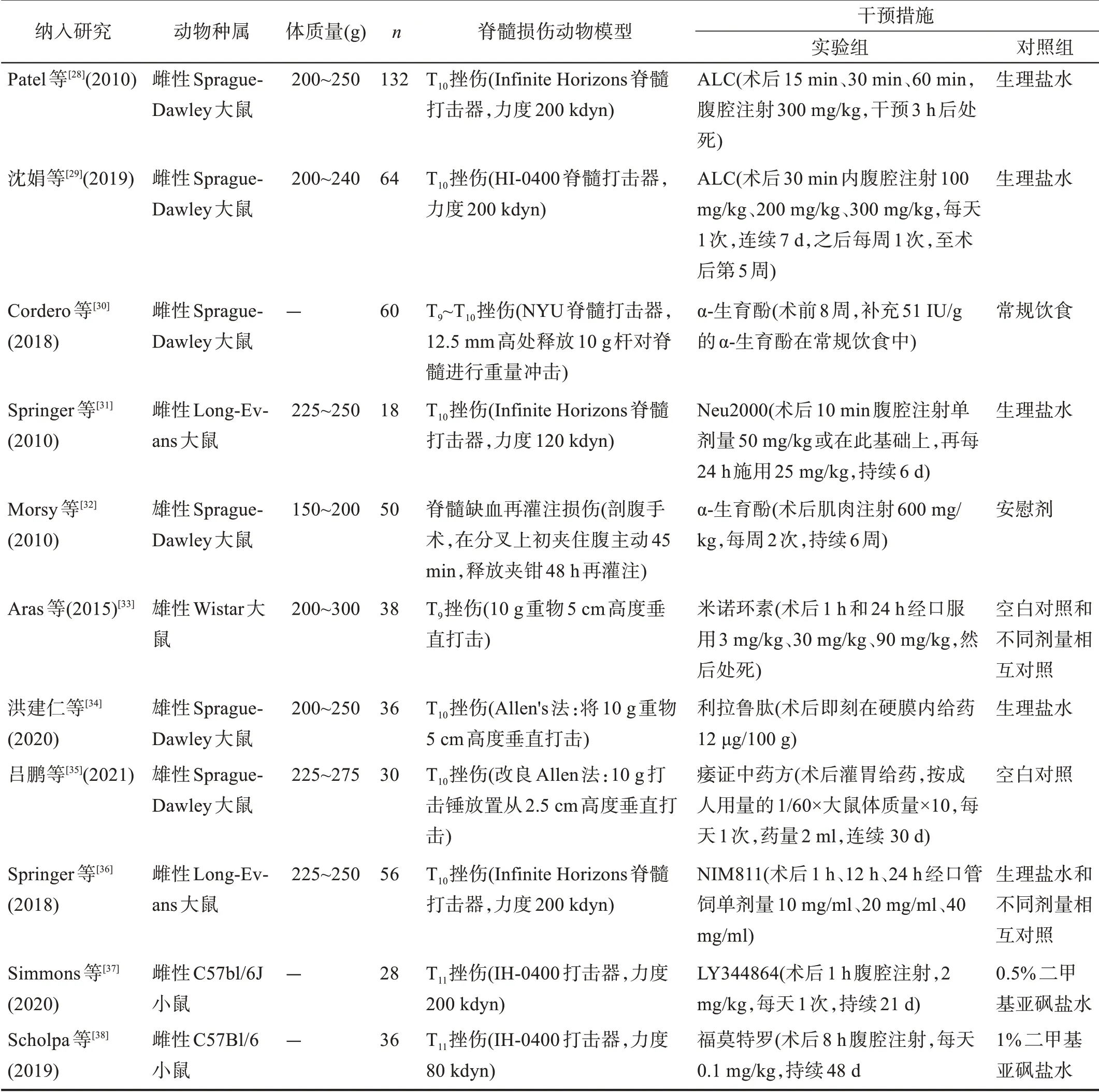
表3 纳入研究的基本特征
2.2 质量评价
2.2.1 偏倚风险评估结果
根据SYRCLE 动物实验偏倚风险评估工具对纳入研究进行评价。11项研究都对实验动物基线情况进行报道,平行可比。文献均提及“随机”或“随机对照试验”,但其具体的随机化方法和是否采取适当的方法来实现随机序列的不可预测性尚不清楚。7 项研究交代对饲养者和研究者实施盲法;7 项研究报告对结果评价者实施盲法;所有研究均对动物进行随机化安置;7项研究对结局指标进行随机评估;9项研究的数据报告完整。见表4。
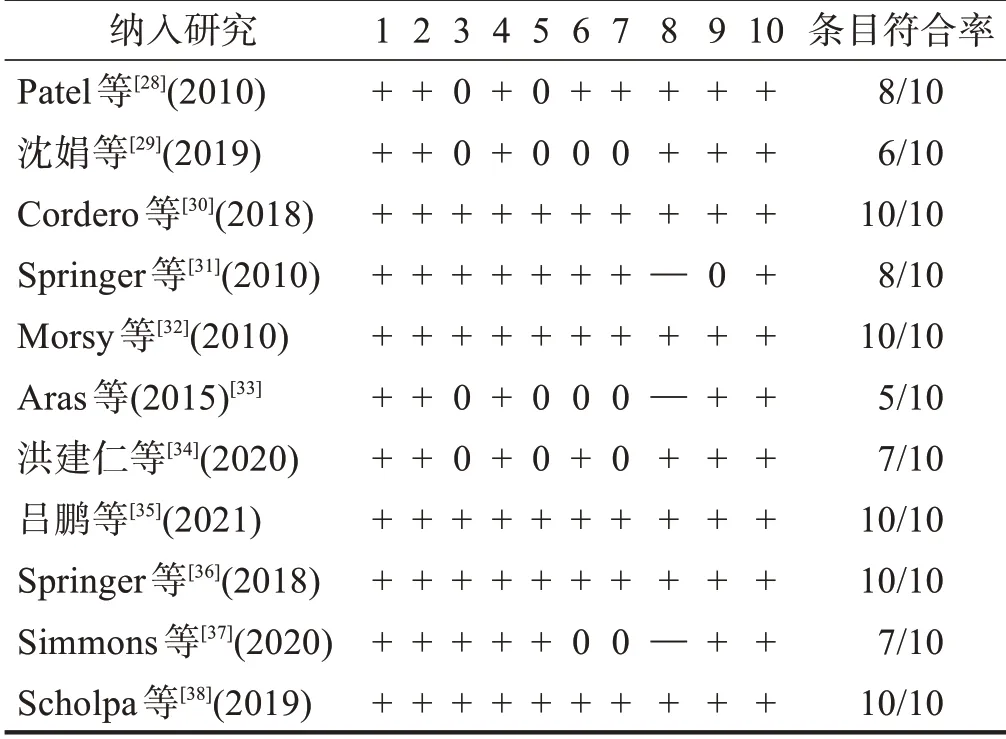
表4 纳入研究的偏倚风险评估结果
2.2.2 CERQual证据质量评估结果
CERQual 证据质量评估结果5 个结局指标中,证据质量为高、中和低。证据质量降级的原因包括原始研究的方法学限制、概括性较差和较多数据无法进行合并和转化等。见表5。
2.3 系统评价结果
11 项研究中,6 项研究选用雄性或雌性Sprague‑Dawley 大 鼠[28‑31,34‑35],其 余5 项研究 选择其 他种属鼠[32‑33,36‑38],8 项研究大鼠体质量150~275 g[28‑29,31‑36],所有研究的脊髓损伤动物模型集中于T9~T11挫伤性脊髓损伤,但使用的脊髓打击器和打击力度存在差异。研究的干预措施存在较大异质性,干预药物的种类、使用时间、频率、浓度和剂量等均不相同。在对照组中,5 项研究使用生理盐水[28‑29,32,34,36],2 项研究使用空白对照和相互对照[33,35]。由于纳入研究在动物种属、动物模型和结局指标测量等方面存在较大的异质性,因此无法对不同研究数据进行Meta 分析,只进行定性描述。
2.3.1 运动功能
9 项研究报告运动功能,其中7 项研究采用BBB评分评估运动功能,发现与对照组动物相比,实验组动物的BBB 评分更 高[28‑30‑31‑36,38]。Springer 等[31]在BBB评分基础上,还利用DigiGait 图像分析系统,显示与假手术动物相比,安慰剂对照组在站立阶段花费的步幅持续时间百分比显著增加,但在Neu2000 治疗后显著减少(与安慰剂对照组相比)。另外,分别有一项研究采用BSM 评分[21]和Rivlin 斜板实验[20],结果显示,与对照组相比,实验组动物运动功能改善。
2.3.2 组织病理学
8 项研究报告脊髓组织病理学,其中3 项研究采用HE 染色[29,34‑35],5 项研究采用铬花青染色[28,32,36‑38],5项研究进行定量分析脊髓受损组织和残余组织体积[28,32,36‑38],与对照组相比,在整个脊髓横断面上治疗组受损组织体积较小,残存体积有所增加。但Spring‑er 等[31]报告,与对照组相比,NIM811 的20 mg/kg 和40 mg/kg 剂量均显著降低病变的平均体积,10 mg/kg的实验组却没有。

表5 纳入研究的CERQual证据质量评估结果
2.3.3 抗氧化功能
6 项研究报告抗氧化功能,其中4 项报告MDA 和SOD[29,31,33,35],分别有2 项报告GSH‑Px[33,35]和CAT[31,35],与对照组相比,实验组在干预后血清MDA 水平降低,SOD、GSH‑Px和CAT水平增加。
2.3.4 抗凋亡功能
2项研究报告抗氧化功能,均采用TUNEL染色进行检测。与对照组相比,实验组在干预后细胞凋亡指数降低[29],逆转凋亡相关蛋白(Smac/Diablo、细胞色素C、FAS 配体、肿瘤坏死因子‑α、缺氧诱导因子‑1α、P53、Bax和Bcl‑2等)的表达[34]。
2.3.5 线粒体生物发生
3 项研究报告线粒体的生物发生,其中1 项报告线粒体呼吸速率和关键线粒体酶复合物的活性,结果显示与对照相比,实验组干预显著保持了线粒体的呼吸和酶活性,即维持线粒体的生物能[28]。1 项报告损伤部位的线粒体DNA 和蛋白质含量,结果显示与假对照组相比,脊髓损伤后3 d,损伤和周围损伤部位的线粒体DNA 含量降低约45%;LY344864 减轻了损伤部位DNA的减少,并且增加线粒体DNA含量(相较于对照组)[37]。1项报告线粒体生物发生相关蛋白PGC‑1α,脊髓损伤后骨骼肌中PGC‑1α 降低,但经实验组干预后可增加[38]。
3 讨论
本研究纳入11项动物实验进行系统评价,结果显示,靶向线粒体治疗脊髓损伤可以促进损伤后的运动功能恢复,减少脊髓受损组织和增加残存组织体积而发挥神经保护作用,可能是通过替代性能源、抗氧化、抗凋亡、抑制mPTP 和增强线粒体生物发生等多种作用机制实现。
本研究纳入文献的动物基线和造模方法差异较大。动物的种类、品系、性别、年龄和体质量等基线情况对实验影响较大,纳入研究的动物主要为Sprague‑Dawley 大鼠,脊髓损伤动物模型主要为T9~T11挫伤模型,但造模设备和造模参数存在很大差异。这是由于脊髓挫伤模型是建立最早的脊髓定量损伤模型,是模拟临床急性脊髓损伤的理想模型[39‑40]。该主要用于模拟临床上因暴力撞击而导致的脊髓损伤,其损伤后会造成炎症、缺血和脊髓空洞形成等,是实验性脊髓损伤的标准造模技术[41]。但是由于动物模型需要考虑众多临床需要,脊髓挫伤模型的造模设备和造模参数存在很大差异。脊髓损伤的不同实验模型在研究病变发展、恢复机制和潜在治疗干预措施的不同方面时各有利弊,但灵长类脊髓损伤动物模型可以帮助开发与人类更相关的有效方法[42]。未来应当针对不同动物种属和损伤模型,进一步深入研究以促进临床转化。
纳入的文献干预措施差异较大。干预药物的种类、使用时间、频率、浓度和剂量等均不相同。纳入研究的干预药物包括ALC、α‑生育酚、米诺环素、痿证中药方、NIM811、LY344864 和福莫特罗等[28‑38]。各种药物的药理作用、干预药物浓度(2 mg/kg、25 mg/kg、90 mg/kg 和300 mg/kg)[28,31‑33,37]、开始使用时间(术后即刻、15 min、30 min、1 h 或8 h)[28,32,34,37]、使用频 率(1 次/d、1 次/周 和2 次/周)[29,31]、持续时 间(6 d、30 d、5 周和6 周)[29,31‑32,35]等均不相同。未来应该针对同种脊髓损伤模型和干预药物,对药物浓度、剂量、使用时间和频率进行深入药代动力学分析,尽早进行临床转化。
本研究纳入文献CERQual证据质量等级不高,结局指标抗凋亡功能和增强线粒体生物发生评估为“低”等级,存在选择、实施、测量和失访偏倚。未来应纳入规范设计和实施标准的动物实验。
本研究局限性:①只纳入公开发表的中、英文文献,存在一定语言偏倚;②对灰色文献的检索不够全面,可能存在漏检和产生偏倚;③纳入研究没有进行功效分析以确定样本量大小,基线资料的比较不够详细;④某些研究未交代清楚隐蔽分组和对动物饲养者及结果评价者施盲,可能产生盲法偏倚;⑤由于纳入研究在实验研究设计、干预措施和结局指标等多方面的异质性,本研究无法对纳入研究数据进行定量分析;⑥某些研究质量较低,可能存在发表偏倚。
综上所述,靶向线粒体治疗脊髓损伤可以改善损伤后的运动功能,减少脊髓受损组织和增加残存组织体积而发挥神经保护作用,其可能是通过替代性能源、抗氧化、抗凋亡、抑制mPTP 和增强线粒体生物发生等多种作用机制实现。受纳入研究数量和质量的限制,上述结论尚待更多高质量研究予以验证。
利益冲突声明:所有作者声明不存在利益冲突。
猜你喜欢
军事文摘(2022年8期)2022-11-03 14:22:01
保健医苑(2022年5期)2022-06-10 07:46:38
中国临床医学影像杂志(2021年6期)2021-08-14 02:21:56
海洋通报(2021年1期)2021-07-23 01:55:14
生物学通报(2021年4期)2021-03-16 05:41:26
肝博士(2020年5期)2021-01-18 02:50:18
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
医学研究杂志(2015年7期)2015-06-22 11:01:01
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
中国中医药现代远程教育(2014年21期)2014-03-01 04:32:11
