
基于快速叶绿素荧光鉴定北疆不同时代主栽棉花品种的耐冷性
2021-05-26 16:53李志博徐建伟张小均钟信念闫臣魏亦农
江苏农业科学 2021年7期
李志博 徐建伟 张小均 钟信念 闫臣 魏亦农

摘要:为筛选棉花耐冷资源,建立棉花耐冷性鉴定评价方法,基于快速叶绿素荧光分析技术,采用冷害因子指数结合模糊隶属函数法对北疆不同时代的代表棉花品种在不同生育期的耐冷性进行了鉴定评价,筛选棉花耐冷性的关键快速叶绿素荧光参数。结果表明,随低温胁迫时间的延长,不同生育期棉花功能叶片的Fv/Fm、Fv/Fo、ETo/ABS、RC/CSo、PIABS呈逐渐下降趋势,而Fo/Fm、ABS/RC、DIo/RC呈上升趋势,且相同胁迫时间下吐絮期的荧光值跟其他生育期都有明显差别。同一棉花品种在不同生育期的耐冷性鉴定分级结果不完全相同,全生育期耐冷性达到高抗的有新陆早19号、新陆早33号和新陆早45号,不抗的为新陆早23号。随时代更替,北疆棉花品种耐冷性呈先升后降的趋势,但耐冷性好于其他棉区材料。棉花盛蕾期的耐冷性对全生育期的耐冷性影响最大。除了花铃期,其他生育期的 Fo/Fm 耐冷因子指数对棉花的耐冷性都具有较大的直接和间接作用,可作为棉花耐冷性鉴定的关键因子。本研究探讨了快速叶绿素荧光技术在棉花耐冷性鉴定上的应用,评价了北疆不同时代主栽棉花不同生育期的耐冷性,建立了不同生育期棉花耐冷性鉴定的线性回归模型。
关键词:棉花;快速叶绿素荧光;耐冷性;生育期;北疆
中图分类号: S562.01;Q945.78文献标志码: A
文章编号:1002-1302(2021)07-0078-11
收稿日期:2020-06-18
基金项目:国家自然科学基金(编号:31560074);石河子大学动植物培育专项(编号:YZZX201602);新疆生产建设兵团绿洲生态农业重点实验室开放课题(编号:201902)。
作者简介:李志博(1978―),男,甘肃镇原人,博士,高级实验师,主要从事棉花抗逆生理及分子生物学机制及新品种选育研究。E-mail:lzb_oea@shzu.edu.cn。
通信作者:魏亦农,教授,主要从事棉花遗传育种研究。E-mail:weiyinong@163.com。
低温是影响植物地域分布、生长发育、产量和品质的基本因素[1-3]。棉花属冷敏感性作物,低温冷害是影响棉花生产的最主要灾害。北疆棉区作为新疆棉花生产的一个重要分支,在我国棉花可持续发展中具有重要地位[4],但该区位于天山北坡海拔400 m以下,积温低、无霜期短,属典型的早熟或极早熟棉区,容易受到倒春寒和早霜的影响[4-5],导致棉花种子萌发力降低[6-8],幼苗抗冷性变弱[7-9],生育期延迟[5,10],产量下降[10-11]以及品质变差等[12-13],因此,北疆棉花的早熟性和幼苗期等生育早期的耐冷性鉴定、生理机制等方面得到了早期研究者们的普遍关注[6-9,14]。近年来,随着全球气候变暖,极端气候类型增加,中后期棉花遭受低温冷害的频率增加,北疆棉花早熟性有所降低[15],潜在地增大了该区棉花受低温冷害的风险。鉴定北疆棉花不同生育时期的耐冷性,了解棉花品种的耐冷性演替规律,对建立棉花耐冷性鉴定方法、探索棉花耐冷机制、确定北疆棉花育种目标等具有重要意义。
叶绿素荧光分析技术是一种基于光合作用理论的探测植物光合生理状况及外界因子影响的活体测定和诊断技术,能“内在性”反映叶片光合作用过程中光系统对光能的吸收、传递、耗散、分配等过程[16],在植物的光合作用机制[16]和抗逆生理[17-19]研究上得到了大量应用,其参数常作为逆境条件下植物抗逆反应的指标之一。基于生物膜能量流动为基础的快速荧光诱导动力学因其获得数据量大、仪器便于携带且操作简便等特点[20],近年来越来越多地被应用于植物抗逆性鉴定[17-19,21]、植物生理[19]、遗传育种[21-23]等领域。本研究拟以快速叶绿素荧光诱导动力学测定为基础,对北疆棉区不同世代的代表性棉花在不同生育时期的耐冷性进行鉴定评价,期望筛选耐冷型棉花资源,建立棉花耐冷性鉴定评价的快速荧光动力学方法。
1 材料与方法
1.1 材料
参试材料28份,其中24份为北疆棉区不同时代选育、种植的代表性品种;3份引自中国农业科学院棉花种质资源中心,苗期耐冷性比较突出;1份为新疆南疆棉区主栽的中熟棉花品种。材料均由石河子大学棉花研究所收集和保存,材料基本信息见表1。
1.2 方法
1.2.1 试验设计
各参试棉花材料种子于2019年4月20日人工点播于石河子大学棉花研究所专用育种大田。栽培模式为覆膜高密度膜下滴灌,宽窄行种植,行距30 cm+60 cm+30 cm,株距10 cm。每膜种植4小行,1膜为1小区,小区长8 m、宽 1.5 m。每材料种植1小区,管理方式同常规大田。
分别于苗期、盛蕾期、花铃期、吐絮期在每个材料种植小区中段选不缺苗且生长一致的连续7株棉株,带叶柄剪取功能叶用自封袋密封放在专用取样盒中带回实验室,在RZX型人工智能气候箱中进行6 d的4 ℃低温处理。
1.2.2 快速叶绿素荧光参数测定
用英国Hansatech公司的植物效率仪(PEA)每天测定各棉花材料低温处理(0 d为处理前)的快速葉绿素荧光参数,测定前叶片暗适应20 min。仪器设置激发光强3 500 μmol/(m2·s)。最大光化学效率(Fv/Fm)、用于热耗散的量子比率(t=0时)(Fo/Fm)、单位面积吸收的光能(t=0时)(ABS/CSo)、单位反映中心吸收的光能(ABS/RC)、单位反应中心耗散的能量(t=0时)(DIo/RC)、单位反应中心捕获的用于还原QA的能量(t=0时)(TRO/RC)、单位反应中心捕获的用于电子传递的能量(t=0时)(ETo/RC)及以吸收光能为基础的性能指数PIABS等参数由仪器自动测定读出,用于计算电子传递的量子产额(t=0 时)[ETo/ABS=(ETo/RC)/(ABS/RC)]、单位面积内反应中心的数量(t=0时)[RC/CSo=(ABS/CSo)/(ABS/RC) ][24]。
1.2.3 耐冷因子指数计算及耐冷性鉴定
参考Strauss等[24]和李志博等[23]的鉴定方法并略有改動计算快速叶绿素荧光参数耐冷因子指数(CFI):CFI=∑Wp·LOGRp(CFP),Wp指不同处理时期的权重系数,Rp(CFP)指不同处理时期相对叶绿素荧光参数,为该处理时期相应时间(d)的相对荧光参数的平均值。本试验分为3个胁迫处理时期,其中第1天和第2天为短期处理,第3天和第4天为中期处理,第5天和第6天为长期处理,各处理时期的权重系数依次为1、3、5。
用模糊隶属函数法鉴定评价参试材料的耐冷性。模糊隶属函数公式为Y(u)=(Y-Ymin)/(Ymax-Ymin),与棉花耐冷性负相关用反隶属函数公式:Y(u)=1-(Y-Ymin)/(Ymax-Ymin)中。公式中,Y指不同参试材料的耐冷因子指数值,Ymax指所有参试材料中耐冷因子指数最大值,Ymin指所有参试材料中耐冷因子指数最小值。棉花耐冷性分级参考付凤玲等的标准[25]略有修改:≥0.75 为高抗,0.61~0.74为抗,0.46~0.60为中抗,0.31~0.45为弱抗,≤ 0.30为不抗。某一荧光参数下的耐冷性为该荧光参数耐冷因子指数隶属值,不同生育期棉花的耐冷性为该生育期各荧光参数耐冷因子指数隶属值的平均值,全生育期棉花耐冷性为各生育期棉花耐冷性的平均值。
1.2.4 数据处理与分析
用SPSS 13.0对测试数据进行统计分析,用SIGMAPLOT 12.5进行制图。分析和作图数据均为平均值±标准误。
2 结果与分析
2.1 不同生育期胁迫时间对棉花主要快速荧光参数的影响
由图1可见,随着胁迫时间的延长,各生育期棉花叶片的Fv/Fm、Fv/Fo、ETo/ABS、RC/CSo、PIABS均呈明显下降变化趋势,且同一胁迫时间下,幼苗期、盛蕾期和花铃期间各荧光参数值差异不大,而吐絮期的各荧光参数值都低于其他3个生育期,尤其胁迫第3天起,差异更加明显。不同生育期棉花叶片的ABS/RC、DIo/RC随胁迫时间明显上升,且吐絮期的上升幅度大于其他生育期。不同生育期棉花叶片的TRO/RC、ETo/RC随胁迫时间呈先上升后下降的趋势,且苗期的下降时间较晚,幅度较小。
2.2 不同生育期各棉花的耐冷因子指数及耐冷性评价
根据不同生育期棉花主要荧光参数随胁迫时间的响应趋势,选择了受胁迫时间影响且变化规律明显的荧光参数计算其耐冷因子指数,以耐冷因子指数为参数鉴定了各个棉花材料在该参数下的耐冷性及整个生育期的耐冷性。可以看出,同一生育期不同材料的耐冷性具有明显差异,且同一材料不同荧光参数评价的耐冷性分级结果不完全相同。
苗期(表2):中棉所36号(MH25)和中JX4(MH27) 8个耐冷因子指数隶属值的鉴定结果均为高抗,新陆早54号(MH20)除了ABS/RC和RC/CSo隶属值的鉴定结果为抗、新陆早66号(MH23)除了Fo/Fm和ETo/ABS鉴定为抗外,其余指标的鉴定结果均为高抗;而新陆早7号(MH1)和新陆早24号(MH8)8个耐冷因子指数隶属值的鉴定结果均为不抗,新陆早23号(MH7)也有6个耐冷因子指数隶属值的鉴定结果为不抗。
盛蕾期(表3):新陆早7号(MH1)、新陆早8号(MH2)、新陆早19号(MH5)、新陆早21号(MH6)、新陆早26号(MH10)、新陆早35号(MH12)、新陆早74号(MH24)和新陆中26号(MH28)的8个冷害因子指数的隶属值鉴定结果均为高抗,新陆早24号(MH8)和新陆早25号(MH9)有6个冷害因子指数的隶属值鉴定结果为高抗;但新陆早23号(MH7)、新陆早42号(MH15)和新陆早50号(MH19)有6个耐冷因子指数隶属值的鉴定结果为不抗。
花铃期(表4):新陆早24号(MH8)、新陆早36号(MH13)和新陆早45号(MH16)的8个耐冷因子指数的鉴定结果均为高抗,新陆早13号(MH14)和新陆早19号(MH5)有7个耐冷因子指数的鉴定结果为高抗、1个鉴定结果为抗;而新陆早54号(MH20)、中棉所36号(MH25)的8个耐冷因子指数鉴定结果均为不抗。
吐絮期(表5):新陆早19号(MH5)、新陆早7号(MH1)和新陆早60号(MH21)分别有3~4个耐冷因子指数值在28个材料中达到最值,其他耐冷因子指数值在所有材料中的位次也十分靠前,耐冷性综合隶属值鉴定结果为高抗;新陆早33号(MH11)、新陆早35号(MH12)、新陆早54号(MH20)和新陆早74号(MH24)尽管没有耐冷因子指数位于最值,但几乎所有的耐冷因子指数位次都平均靠前,耐冷性隶属值综合鉴定结果也为高抗。新陆早10号(MH3)、新陆早48号(MH18)、中棉所36号(MH25)和中JX4(MH27)的耐冷性隶属值综合鉴定结果为不抗。
从不同生育期的耐冷性鉴定结果来看,同一棉花在不同生育期的耐冷性分级结果不同,有时甚至会有很大的差别,如新陆早7号在盛蕾期和吐絮期的耐冷性鉴定结果为高抗,而幼苗期的鉴定结果为不抗,花铃期的鉴定结果为抗;中JX4在幼苗期和盛蕾期的耐冷性鉴定结果为高抗,而花铃期和吐絮期的鉴定结果为不抗。不同生育期参试棉花材料的耐冷性鉴定分级的比例有所差别,其中抗性(高抗、抗)为盛蕾期>吐絮期>苗期=花铃期,其比例依次为67.86%、46.43%、39.29%、39.29%,而不抗为花铃期>幼苗期>吐絮期>盛蕾期,其比例依次为21.43%、17.86%、14.29%、10.71%。
2.3 不同生育期北疆不同时代棉花耐冷性演替变化分析
以引进的苗期耐冷性强的棉花(MH25、MH26、MH27)为对照,对不同生育期北疆不同时代棉花的耐冷性变化趋势进行了分析(图2)。可以看出,随时代演替,北疆棉区选育的棉花品种在幼苗期、吐絮期的耐冷性逐步增强,盛蕾期逐步下降,而花铃期表现为先上升后下降趋势。与对照相比,北疆棉花早期(幼苗期、盛蕾期)的耐冷性低于对照,中后期(花铃期、吐絮期)的耐冷性高于对照。全生育期来看,北疆不同时代选育的棉花耐冷性呈先上升后下降的趋势,但耐冷性好于对照。
2.4 不同生育期棉花耐冷快速荧光鉴定体系的分析
2.4.1 不同生育期棉花耐冷性关系及棉花耐冷性關键生育期分析 从表6看出,不同生育期棉花耐冷性的隶属值之间两两呈差异不显著,表明同一棉花品种在不同生育期的耐冷性关系不大,不同生育期棉花的耐冷机制存在差异。但不同生育期棉花的耐冷性对整个棉花生长发育过程中的耐冷性影响不一样,盛蕾期、花铃期和吐絮期的耐冷性对整个棉花全生育期的耐冷性有极显著影响,而苗期影响不大,按影响大小依次为盛蕾期>花铃期>吐絮期>苗期。
2.4.2 不同生育期影响棉花耐冷性的关键荧光参数分析 从表7看出,除了吐絮期的RC/CSo,其他生育期的每一个指标因子的评价结果均与该生育期棉花耐冷性的整体评价结果呈极显著正相关,表明每个荧光指标因子间存在着密切的相互作用或共同影响着棉花的耐冷性。
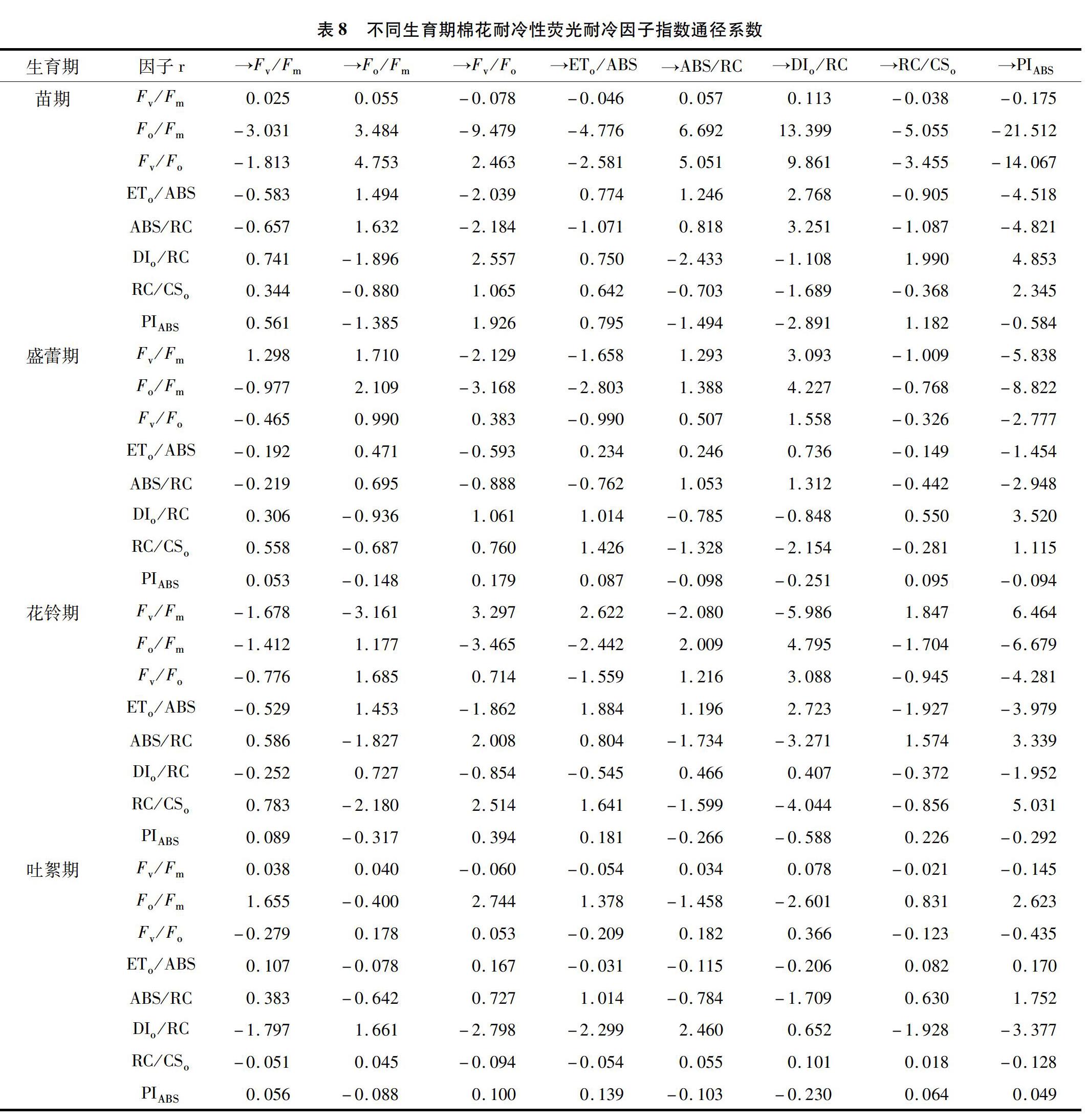
2.4.3 不同生育期影响棉花耐冷性的荧光参数耐冷因子指数通径分析 为更好地了解荧光耐冷因子指数之间的关系及其对棉花耐冷性的作用效应,对耐冷因子指数进行了通径分析,从分析结果(表8)可以看出,苗期Fo/Fm耐冷因子指数的直接通径系数在所有直接通径系数中最大,且其间接通径系数也是所有系数中最大,此外,Fv/Fo也具有较高的直接和间接通径系数;盛蕾期直接效应和间接效应表现最突出的是Fo/Fm耐冷因子指数;花铃期直接通径系数最大的依次为ETo/ABS、ABS/RC、Fv/Fm、Fo/Fm,但它们之间差别不大,且Fv/Fm、Fo/Fm对其他因子的间接系数远远大于ETo/ABS、ABS/RC对其他因子的间接系数;而吐絮期直接通径系数较大的有ABS/RC、DIo/RC、Fo/Fm,但Fo/Fm和DIo/RC耐冷因子指数对其他荧光参数耐冷因子指数的间接效应强于ABS/RC。
总体来看,各个生育期的最大直接通径系数耐冷因子指数略有不同,说明各个生育期棉花耐冷性的耐冷机制和关键因子耐冷指数是有差别的。但几乎各个生育期Fo/Fm耐冷因子指数的直接通径系数及其对其他耐冷因子指数的间接通径系数都最大,表明对各个生育期的棉花耐冷性均具有较强的直接和间接作用,可作为不同生育期棉花耐冷性评价的通用指标。
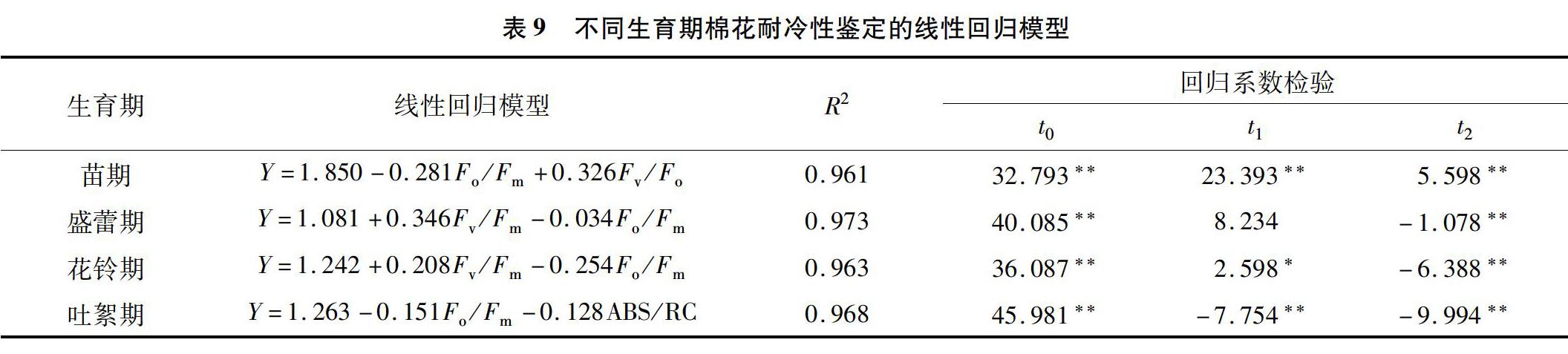
2.4.4 不同生育期基于棉花耐冷性关键影响因子的线性回归分析 选取直接效应和间接效应都比较突出的耐冷因子指数为自变量,棉花的耐冷性为依变量建立不同生育期棉花耐冷性的鉴定模型(表9),每个模型的决定系数都在0.96以上,反映了鉴定模型中所选的耐冷因子指数决定了棉花耐冷性的绝大部分,是影响棉花耐冷性或可作为棉花耐冷性评价的关键因子,而且线性回归模型的回归系数检验几乎都达到了极显著差异水平。
3 讨论与结论
3.1 快速叶绿素荧光技术在棉花耐冷性鉴定评价上的应用
正常的光合能力是植物耐逆性的重要生理基础。叶绿素荧光技术能“内在”地反映植物光合作用信息[16-17],已广泛应用于植物的抗逆资源鉴定和筛选[18-19,21-23],且在植物抗冷性上已显示出良好的应用前景[18,23]。有研究发现,低温下多数植物会产生光抑制,对叶绿素荧光参数产生显著的影响,但叶绿素荧光参数变化与植物种类、处理温度、处理方式等有关系,迄今还没有一个叶绿素荧光参数可以有效地鉴定评价所有植物的抗寒能力[18],因此用叶绿素荧光技术筛选和鉴定抗寒植物时采用的叶绿素荧光参数指标有所不同。以生物膜能量流动为基础的快速叶绿素荧光诱导曲线的数据分析方法(JIP-test),可以捕捉诱导曲线上升部分更丰富的信息[17,20],且O-J-I-P 曲线的形状和参数对外界环境的变化十分敏感[20,23-24],越来越多地应用于植物的耐逆性鉴定评价。本课题组曾初次用O-J-I-P 动力学的Fv/Fm和 PI 参数研究了北疆棉区 22个主栽棉花的幼苗耐冷性差异[23],结果跟Strauss等对南非大豆耐冷性分类的结果[24]相符。本试验用快速叶绿素荧光进一步分析了北疆不同世代主栽棉花在不同生育时期的耐冷性,涉及了多个快速荧光参数,研究结果表明,不同生育期的棉花耐冷性鉴定的适宜快速荧光参数不同。
3.2 不同生育期棉花耐冷性差异及耐冷指标筛选
植物耐冷性是植物对低温胁迫强度和时间的综合适应,同时受生长发育时期、处理方法、环境因素等各个因素的影响[1,7,9,14]。不同棉花在不同生育时期的耐冷性不完全一致[7],表明棉花耐冷性具有多样性和丰富的遗传基础,同时也说明不同生育期棉花耐冷机制不一样,需要对不同生育期的棉花耐冷指标分别进行筛选。有研究表明,棉花种子的萌发能力随温度的降低而下降,种子萌发指标与种子耐冷萌发力显著相关[6-7,14],其中发芽指数为低温下影响棉花种子萌发的关键指标[6,14],与辣椒、玉米等萌发期抗冷鉴定采用发芽指数作为评价指标相同,但王俊娟等认为,由于受子叶光合作用的影响,萌发期棉花抗冷鉴定不宜采用萌发指数或出苗率作为评价指标,可以0 ℃处理 4 d,28 ℃条件下恢复正常生长 7 d 的相对子叶平展率作为鉴定指标[7]。在苗期耐冷性鉴定中,很多人将幼苗的外观形态作为耐冷鉴定标准[9],如低温后的死苗率[7],冷害指数[26]等,而且普遍认为,冷害指数和棉花幼苗的耐冷性密切相关[7,9,27]。在生理生化响应上,抗氧化酶活性、膜稳定性及渗透调节物质是植物耐冷研究的主要内容。多数研究表明,主要抗氧化酶POD、CAT活性在不同耐冷型棉花幼苗间有差异变化[7,26],而SOD活性变化不大,同时王俊娟等发现,棉花幼苗地下部根系的抗氧酶活性比地上部对低温冷害的响应更明显[7]。细胞质膜被认为是植物低温胁迫的关键位点,膜稳定或透性相关指标也常被人们用来作为衡量植物耐冷性的指标,棉花幼苗叶片的细胞膜透性物质(MDA)与棉苗耐冷性密切相关,可作为棉花幼苗耐冷性鉴定的指标[7,25],还有研究发现,棉花幼苗的耐冷性跟可溶性蛋白含量、脯氨酸含量等有关。而王冀川等认为,棉花的耐冷性是多个指标因子共同作用的结果,用单一或几个指标评价棉花的耐冷性可能存在误差[27]。
由于受低温处理条件等因素的影响,前人对棉花耐冷的研究多局限于生育前期[6-7,9,14,28](萌发期、幼苗期),而生育中后期是棉花生殖生长、产量、品质形成的关键时期[4,11,15],是棉花温度需求较高的时段,低温冷害的影响可能更大,低温冷害研究也具有重要意义。在建立不同生育时期的棉花耐冷鉴定的快速叶绿素荧光模型基础上,本试验初次对棉花生育期后期的耐冷性鉴定评价指标做了筛选,研究发现,快速荧光参数Fo/Fm、Fv/Fm为花铃期棉花耐冷性的关键指标,Fo/Fm和ABS/RC为吐絮期棉花耐冷性的关键指标。研究还发现,不同生育期棉花的耐冷性对整个生育期棉花耐冷性的影响大小为盛蕾期>花铃期>吐絮期>苗期,进一步说明了后期低温对棉花较大的影响作用。
3.3 北疆棉花耐冷性评价及早熟育种的必要性
新疆是我国最大的棉花生产基地,北疆亚棉区是新疆棉区一个重要组成部分[4]。相对其他亚棉区,北疆棉区积温低,棉花生育期短[5,10,15],因此,研究者们对北疆棉区适宜棉花材料的耐冷性进行大量的鉴定和筛选,早熟也成为北疆棉区棉花育种的主要目标。近年来,随着全球气温普遍增温,棉花适宜生育期有延长趋势[15],生育期较长的品种相对更容易获得高产,引起育种者们有忽略棉花早熟性的倾向,如李春平等在分析北疆棉花早熟育种问题时认为,北疆棉区棉花风险面积比例较小,应选育生育期在 125 d 或稍长的品种[29],而本研究也发现,2011—2017年间审定的适宜北疆棉区的代表棉花品种比2001—2010年间的代表棉花品种的耐冷性降低,因此,北疆棉花在坚持棉花品质和产量为首要育种目标时,依然应该把早熟性放到棉花育种目标的首要地位,不但是抵御棉花遭受的低温冷害伤害的需要,而且在未来棉花机械采收的叶片脱落和脱叶剂的施用效应上可能发挥着重要的作用。
快速叶绿素荧光技术可以应用于棉花耐冷性鉴定评价。不同生育期棉花耐冷性鉴定的快速荧光指标不同,建立了不同生育期棉花耐冷性鉴定的数学模型。在参试材料中,全生育期耐冷性表现高抗的棉花有新陆早19号、新陆早33号和新陆早45号,不抗的为新陆早23号;随时代更替,北疆不同世代选育的棉花品种耐冷性呈先上升后下降的趋势,但耐冷性好于其他棉区材料。
参考文献:
[1]Margesin R,Neuner G,Storey K B. Cold-loving microbes,plants,and animals-fundamental and applied aspects[J]. Die Naturwissenschaften,2007,94(2):77-99.
[2]Yadav S K. Cold stress tolerance mechanisms in plants. A review[J]. Agronomy for Sustainable Development,2010,30(3):515-527.
[3]刘 辉,李德军,邓 治. 植物应答低温胁迫的转录调控网络研究进展[J]. 中国农业科学,2014,47(18):3523-3533.
[4]新疆棉花高产优质高效理论与实践[M]. 乌鲁木齐:新疆科学技术出版社,2004.
[5]周小云,胡文冉,杨 洋,等. 北疆春季棉花延迟型冷害的综合防治措施[J]. 棉花科学,2016,38(5):46-49.
[6]高利英,邓永胜,韩宗福,等. 黄淮棉区棉花品种种子萌发期低温耐受性评价[J]. 棉花学报,2018,30(6):455-463.
[7]王俊娟,王德龙,阴祖军,等. 陆地棉萌发至幼苗期抗冷性的鉴定[J]. 中国农业科学,2016,49(17):3332-3346.
[8]Gai Y P,Li X Z,Ji X L,et al. Chilling stress accelerates degradation of seed storage protein and photosynthetic protein during cotton seed germination[J]. Journal of Agronomy and Crop Science,2008,194(4):278-288.
[9]王俊娟,王 帥,陆许可,等. 棉花幼苗对低温胁迫的响应及抗冷机制初步研究[J]. 棉花学报,2017,29(2):147-156.
[10]谢 芳,童 忠,刘 静. 新疆乌苏市近20a气候变化及其对棉花生育期的影响[J]. 中国棉花,2018,45(10):36-38,40.
[11]李玉强. 低温冷害对尉犁县棉花生长发育的影响分析[J]. 农业灾害研究,2018,8(5):42-43.
[12]Zheng M,Wang Y,Liu K,et al. Protein expression changes during cotton fiber elongation in response to low temperature stress[J]. Journal of Plant Physiology,2012,169(4):399-409.
[13]Jimmy Z,James R,Matthew I. Impact of temperature and relative humidity on the near infrared spectroscopy measurements of cotton fiber micronaire[J]. Journal Indexing and Metrics,2018,88(20):2279-2291.
[14]徐建伟,张 晨,曾晓燕,等. 近十年新疆北疆主栽棉花种子低温萌发能力差异评价[J]. 新疆农业科学,2017,54(9):1569-1578.
[15]胡莉婷,胡 琦,潘学标,等. 气候变暖和覆膜对新疆不同熟性棉花种植区划的影响[J]. 农业工程学报,2019,35(2):90-99.
[16]Baker N R. Chlorophyll fluorescence:a probe of photosynthesis in vivo[J]. Annual Review of Plant Biology,2008,59:89-113.
[17]Kalaji H M,Rastogi A,Zˇivcˇák M,et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping:an example of barley landraces exposed to various abiotic stress factors[J]. Photosynthetica,2018,56(3):953-961.
[18]周蕴薇,刘艳萍,戴思兰. 用叶绿素荧光分析技术鉴定植物抗寒性的剖析[J]. 植物生理学通讯,2006,42(5):945-950.
[19]Kalaji H M,Jajoo A,Oukarroum A,et al. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions[J]. Acta Physiologiae Plantarum,2016,38(4):102.
[20]李鹏民,高辉远,Strasser R J. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J]. 植物生理与分子生物学学报,2005,31(6):559-566.
[21]张淑英,褚贵新,梁永超. 不同铵硝配比对低温胁迫下棉花幼苗光合及叶绿素荧光参数的影响[J]. 干旱地区农业研究,2018,30(5):34-39.
[22]Zushi K,Matsuzoe N. Using of chlorophyll a fluorescence OJIP transients for sensing salt stress in the leaves and fruits of tomato[J]. Scientia Horticulturae,2017,219:216-221.
[23]李志博,华显伟,魏亦农,等. 叶绿素荧光动力学 O-J-I-P 参数在棉花幼苗耐冷性评价上的应用[J]. 棉花学报,2010,22(2):132-137.
[24]Strauss A J,Kruger G H,Strasser R J,et al. Ranking of dark chilling tolerance in soybean genotypes probed by the chlorophyll a fluorescence transient O-J-I-P[J]. Environmental and Experimental Botany,2006,56(2):147-157.
[25]付凤玲,李晚忱,潘光堂. 模糊隶属法对玉米苗期耐旱性的拟合分析[J]. 干旱地区农业研究,2003,21(1):83-85,125.
[26]李志博,曹連莆,魏亦农,等. 荒漠绿洲区棉花幼苗耐寒生理特性及其评价指标分析[J]. 棉花学报,2010,22(6):568-573.
[27]王冀川,黄 琪,徐雅丽.棉花抗冷指标的灰色关联度分析[J]. 江西棉花,2001,23(2):6-9.
[28]夏 军,王远远,乔 露,等. 营养元素引发对低温下棉苗保护性酶活性及叶绿素荧光参数的影响[J]. 江苏农业科学,2019,47(16):89-92.
[29]李春平,刘忠山,张大伟,等. 北疆棉花早熟育种探讨[J]. 中国棉花,2014,41(1):5-7.
猜你喜欢
少儿科学周刊·儿童版(2022年10期)2022-06-27
中国土壤与肥料(2021年5期)2021-12-02
小读者(2020年4期)2020-06-16
——军旅写生作品展
军营文化天地(2016年5期)2016-07-26
西南农业学报(2016年5期)2016-05-17
河北书画研究(2016年6期)2016-02-03
西藏科技(2015年5期)2015-09-26
新疆农垦科技(2014年12期)2014-02-28
新疆农垦科技(2014年10期)2014-02-28
奋斗(2013年12期)2013-01-27
