
水稻护颖发育相关基因的研究进展
2021-05-24 09:14罗曦魏林燕郑燕梅魏毅东连玲谢华安吴方喜
南京农业大学学报 2021年3期
罗曦,魏林燕,郑燕梅,魏毅东,连玲,谢华安,吴方喜*
(1.福建省农业科学院水稻研究所,福建 福州 350019;2.农业农村部华南杂交水稻种质创新与分子育种重点实验室/福州(国家)水稻改良分中心/福建省作物分子育种工程实验室/福建省水稻分子育种重点实验室/福建省作物种质创新与分子育种省部共建国家重点实验室培育基地/杂交水稻国家重点实验室华南研究基地/水稻国家工程实验室,福建 福州 350003)
当水稻受到短日照等环境因素诱导后,就开始从营养生长阶段向生殖生长阶段过渡[1]。产生最后一片营养叶(foliage leaf)后,顶端分生组织(shoot apical meristem,SAM)开始向花序分生组织(inflorescence meristem,IM)过渡,随后依次是枝梗分生组织(branch meristem,BM)、小穗分生组织(spikelet meristem,SM)和小花分生组织(floral meristem,FM)的连续转变,最终形成一个完整的花序[2]。小穗是水稻花序的基本单元,小穗的结构按分生顺序依次为:副护颖(rudimentary glumes)、护颖(sterile lemmas)、外稃(lemma)、内稃(palea)、浆片(lodicule)、雄蕊(stamen)、雌蕊(pistil),雌蕊又包括子房(ovary/carpel)和柱头(stigma),子房内部是胚珠(ovule)(图1)。

rg:副护颖 Rudimentary glume;sl:护颖 Sterile lemma;le:外稃 Lemma;pa:内稃 Palea;lo:浆片 Lodicules;pi:雌蕊 Pistil;st:雄蕊 Stamen;标尺 Scale bar:1 mm.图1 水稻小穗结构图Fig.1 Structure chart of rice spikelet
小穗是禾本科特有的花序结构。与其他禾本科作物相比,水稻小穗有其专有的特征:1)水稻每个小穗只有1朵小花;2)护颖是水稻特异器官,其他禾本科作物没有其对应器官。目前,关于水稻护颖的起源有2种假说。一种假说认为:水稻祖先每个小穗上有3朵小花,1朵顶生小花,2朵侧生小花,侧生小花退化只剩外稃,然后进一步退化成护颖[3-4]。另一种假说认为:护颖与副护颖都是退化的苞片结构[5]。而最近,越来越多的证据支持第1种假说[6-7]。
稻穗发育过程中,护颖的分生阶段在花序分生之后,小花分生之前。许多控制小穗发育的基因都会影响护颖的发育。已报道的与护颖发育有关的基因有10多个,这些基因具有不同的结构特点,并且通过不同的分子机制直接或间接影响护颖的表型。水稻的花序和小穗结构特点是决定籽粒产量的重要因素,例如,每穗粒数、千粒重、一次枝梗数、二次枝梗数等性状直接与产量相关,调控水稻护颖发育的一些基因也是这些性状的调控基因。所以,研究水稻护颖发育的分子机制是阐明水稻花发育机制的一个关键切入点,同时也为水稻增产提供理论基础。本文对与护颖发育相关的基因进行分类和总结,目的是找到它们之间的联系,为进一步研究水稻花发育的分子机制提供线索。
1 护颖发育相关基因的突变体表型及其表达模式
本文总结了12个影响护颖发育的基因(表1)。这些基因分别参与不同阶段的水稻小穗发育进程,这些基因的突变均导致护颖形态或数目的变化。从各基因突变体的护颖表型来看:LONGSTERILELEMMA1(G1)、PANICLE PHYTOMER2(PAP2)/OsMADS34、NONSTOP GLUMES1(NSG1)/LACKING RUDIMENTARY GLUME 1(LRG1)和APICAL DORMANT1(SAD1)抑制护颖向外稃的同源转化;O.sativa VIN3-LIKE2(OsVIL2)、O.sativa INDETERMINATE GAMETOPHYTE1(OsIG1)、O.sativa EXTRA GLUME1(EG1)和ABERRANT SPIKELETS AND PANICLE1(ASP1)不但抑制护颖向外稃同源转化,还影响护颖的数目;而FRIZZY PANICLE(FZP)、SUPERNUMERARY BRACT(SNB)、O.sativa INDETERMINATE SPIKELET1(OsIDS1)和MULTI-FLORET SPIKELET1(MFS1)这4个AP2/EREBP基因促进护颖生长。除了G1基因是调控护颖的特征基因外,其他11个基因在多个器官中都有不同程度的表达。
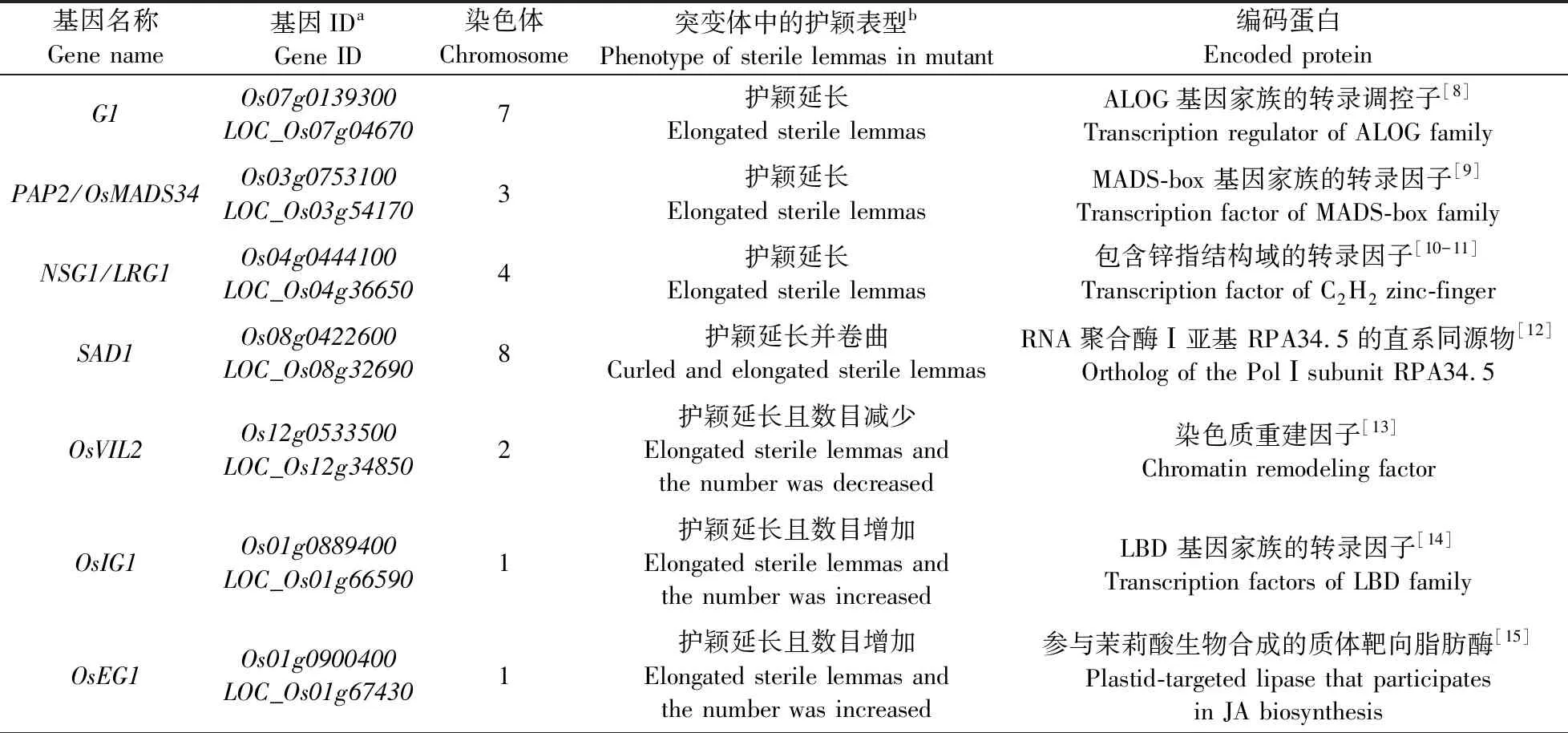
表1 突变体护颖发育异常的基因Table 1 Genes which sterile lemma of mutant with abnormal development

续表1 Table 1 continued
1.1 抑制护颖向外稃同源转化的基因
2009年Yoshida等[8]首先在1个粳稻长护颖突变体中发现G1基因,后来,多个学者又陆续报道多个G1基因的等位基因[21-22],这些研究揭示G1基因具有抑制护颖向外稃转变的作用。Hong等[21]发现,G1基因主要在水稻的护颖中表达,在内稃中也有表达。G1基因首先在一次枝梗、二次枝梗原基分生期的护颖中表达,随着小穗的成熟表达逐渐减弱,当小穗接近成熟时,表达结束。G1基因编码的蛋白具有1个植物特异的、保守的结构域——DUF640,在水稻和拟南芥中各有10个含有DUF640结构域的基因。因为当时已知的具有DUF640结构域的基因只有拟南芥中的参与光敏色素依赖的光信号途径基因LSH1(LIGHT-DEPENDENT SHORT HYPOCOTYLS1)[23]和水稻G1基因,所以,Yoshida等[8]将DUF640结构域命名为ALOG(ArabidopsisLSH1andOryza G1)。Hong等[21]通过网站预测,G1基因没有已知的DNA binding结构域,也没有明显的转录激活功能。推测G1可能作为1个中介或共抑制子调控那些直接控制护颖生长的基因。
MADS-box基因家族是1个成员数量庞大的基因家族,编码的蛋白属于转录因子。根据系统进化关系、基因结构和蛋白结构的不同又将MADS-box基因家族分为Ⅰ型和Ⅱ型两大类型[24]。许多Ⅱ型MADS-box基因都参与植物花器官发育[25]。PAP2/OsMADS34属于MADS-box基因家族的SEPALLATA(SEP)-like亚家族的LOFSEP亚组[9]。PAP2的突变体由于早期发育的小穗分生组织转变为枝梗分生组织,导致一次枝梗数、二次枝梗数增多,并导致副护颖稍微伸长,护颖伸长,叶腋处长出丝状器官,而内稃、外稃、内部花器官没有明显变化,营养阶段也没有受到影响。原位杂交试验结果表明,PAP2基因在花序分生组织、枝梗分生组织、小穗分生组织以及花分生组织中都有表达[9]。
NSG1/LRG1是1个最近发现的影响护颖发育的新基因。该基因编码的蛋白具有1个保守的锌指结构域,并且被认为是具有转录抑制活性的转录因子[10-11]。lrg1突变体表现为一侧副护颖延长,而另一侧副护颖未形成;护颖延长和额外的外稃;内部花器官则正常发育。qRT-PCR结果表明,LRG1/NSG1在所有水稻器官和发育时期都有不同程度的表达,但在水稻穗发育时期明显比营养生长期表达量高。在转基因互补植株小穗的副护颖与护颖中检测到强烈的GFP信号[10]。
SAD1编码RNA聚合酶Ⅰ的1个亚基RPA34.5。sad1突变体中,核糖体RNA的数量明显降低。酵母双杂交试验[12]表明,SAD1蛋白与RNA聚合酶Ⅰ调节复合物的中间模块OsMED4正向互作。RNA聚合酶Ⅰ受到细胞分裂素(CK)的诱导,与野生型相比较,sad1突变体中的细胞分裂素明显增多。sad1突变体具有多效性表型:分蘖数减少,植株矮化,根少且短,剑叶变窄变短,没有叶耳、叶舌,穗变小,护颖及内外稃卷曲延长,小穗数量变少,内部花器官异常。整体而言,sad1突变体的营养生长向生殖生长的过渡阶段会延迟[12]。
1.2 抑制护颖向外稃同源转化且影响数目确定性的基因
OsVIL2基因编码1个可促进水稻开花的染色质重建因子[13]。Yoon等[26]的研究表明,OsVIL2的 T-DNA 突变体与其野生型相比,花期延迟,分蘖减少;副护颖数目增加,护颖数目减少并且延长,出现额外的颖壳状器官,内稃退化;内部花器官形态或数目异常。启动GUS基因表达的试验结果表明,OsVIL2基因在苗期叶片和成熟期的剑叶中均有表达,在穗发育的不同时期则组成型表达[26]。
OsIG1(O.sativa)是玉米IG1(INDETERMINATE GAMETOPHYTE1)基因的同源基因,编码属于LBD基因家族的蛋白[27-28](具有保守的LOB结构域的植物特异性转录因子)。Zhang等[14]研究表明,OsIG1-RNAi干扰植株株高变矮,叶夹角增大,叶片褶皱化;小穗梗延长,护颖伸长、增宽,数目增加;出现卷曲的、额外的颖壳状器官,内稃退化;内部花器官形态或数目异常。qRT-PCR结果表明,OsIG1在水稻幼穗中强烈表达,在成熟的花器官中表达,叶片中低表达,在根、茎等组织中表达最低。用OsIG1的启动子启动GUS基因表达分析表明,主要在幼穗、护颖、内外稃和雌蕊中检测到GUS活性。原位杂交试验结果表明,OsIG1在花序分生组织原基、花分生组织原基、护颖原基中有很高的表达,在雄蕊和胚珠中也有表达,随着花器官发育成熟,OsIG1的表达逐渐降低[14]。
EG1(EXTRA GLUME1)基因编码1个参与茉莉酸合成途径的质体靶向脂肪酶[29]。Li等[15]研究表明,EG1基因突变体eg1-1营养生长阶段没有明显差异,穗数也没有明显差异,副护颖没有异常,护颖变长,数目增加,护颖与内外稃之间形成额外的颖壳状器官,浆片也形成类似于护颖的形状,雄蕊和雌蕊都有形态或数目的异常。原位杂交试验表明,EG1基因在发育中的种子和茎中有很低的表达,在营养生长期的顶端分生组织没有表达,进入花序分生阶段,EG1基因在一次枝梗、二次枝梗中大量表达。随后,在各个花器官分生组织中表达,特别是新发育的分生组织表达量较高。随着小穗发育成熟,EG1基因逐渐减少,当小穗和内部花器官发育结束时,只能检测到非常弱的信号。EG1基因通过影响茉莉酸合成途径中的OsJAZ1、OsMYC2等基因的结合,调控OsMADS1的表达。所以,EG1基因突变体小穗的缺陷特征与OsMADS1突变体类似[15]。
ASP1编码1个类似拟南芥中TPL[30]和玉米中REL2[31]的转录共抑制子。asp1突变体具有多效性表型[16],花序分生组织的退化生长阶段延迟,枝梗分生组织到小穗分生组织的过渡加快。在营养生长阶段,asp1突变体腋芽生长不受抑制,并产生不规则的叶序,暗示该基因与生长素密切相关。asp1突变体的小穗也产生多种不正常的表型:护颖和副护颖延长且数目减少;内、外稃缺失或产生圆柱状的结构,但其内部花器官正常可育。原位杂交试验表明,在苗期,ASP1基因主要在叶原基表达,顶端分生组织不表达(SAM);在营养生长阶段,主要在腋芽分生组织(AM)和根冠原基中表达;在生殖生长阶段,在花序分生组织(IM)和枝梗分生组织(BM)中表达强烈,在整个小穗发育时期都有表达,尤其是在副护颖、护颖、外稃、内稃等侧生器官中表达强烈。
1.3 AP2/EREBP转录因子促进护颖发育
APETALA2/ethylene-responsive element-binding proteins(AP2/EREBP)是转录因子超家族,它含有1个或2个非常保守的AP2/ERF结构域。AP2/EREBP分为2个亚族:EREBP亚族(具有1个AP2/ERF结构域)和AP2亚族[32](具有2个AP2/ERF结构域)。该类转录因子主要参与调控植物发育、植物对激素和胁迫的应答[33]。目前,与水稻护颖发育相关的AP2/EREBP转录因子家族基因有4个,分别是FZP、SNB、OsIDS1和MFS1。
FZP基因属于EREBP亚族(ERF类)。Komatsu等[17]研究的FZP突变体fzp-2和fzp-3只在小穗发育中有缺陷,该突变体的小穗没有发育出护颖和内3轮小花分生组织,而是被高次枝梗代替,这些高次枝梗则由若干个副护颖组成。原位杂交试验结果表明,FZP基因的表达区域在小穗分生组织的副护颖原基的叶腋处,所以,FZP被认为是小穗分生组织的特征基因[34]。Ren等[35]研究的FZP突变体fzp-12和fzp-13的穗发育缺陷表型较fzp-2和fzp-3弱,fzp-12的小穗明显变小,而每穗粒数没有减少,护颖退化变小。原位杂交结果表明:FZP基因不但在副护颖处表达,在护颖、外稃、内稃中也有表达,而在内部花器官不表达。
SNB基因属于AP2亚族。Lee等[18]研究发现,snb突变体小穗没有形成护颖,副护颖增多;产生了额外的类似内、外稃的结构,浆片变长,浆片数量增多;雄蕊减少,极少数小穗没有产生内2轮花器官。SNB基因的突变使snb突变体的苞片(即副护颖)产生阶段延长,使小穗分生组织向花分生组织这一过渡期延迟。所以,snb突变体属于时异性突变体。除了穗部结构发生改变,snb突变体在营养生长期、生殖生长期以及抽穗期的相关性状都没有明显改变。qRT-PCR和原位杂交结果表明,SNB基因在营养阶段表达量低,当顶端分生组织向生殖生长阶段过渡时,SNB基因在顶端分生组织边缘开始表达,之后,在枝梗分生阶段和小穗分生阶段,SNB基因持续高表达,但进入花分生阶段,SNB基因表达则逐渐降低。
OsIDS1和SNB都属于AP2亚族,OsIDS1突变体表型与SNB突变体表型类似,护颖表现为退化或单侧护颖被副护颖替代、多个副护颖、出现额外的类似内外稃结构等表型[19]。原位杂交表明,OsIDS1的表达模式与SNB类似,但是表达量较低,所以,OsIDS1突变体表型比SNB突变体表型弱。单突变体和双突变体的研究[35]表明,这2个基因在水稻小穗中协同行使功能。
MFS1属于EREBP亚族(ERF类),该基因主要参与小穗分生组织确定性和护颖的特征调控。MFS1基因突变导致小穗发育各种缺陷:小穗轴伸长、护颖退化、额外的外稃、内稃退化及数量不定的内3轮花器官[20]。原位杂交试验结果表明,MFS1基因先在枝梗分生组织和小穗分生组织中大量表达,然后在护颖原基和内/外稃原基中强烈表达,最后在浆片、雄蕊、雌蕊中表达,当雌蕊形成后,MFS1基因在护颖和内/外稃中检测不到转录信号,在浆片、雄蕊、雌蕊中表达量逐渐降低。
2 护颖发育相关基因之间的调控关系
植物花的发育主要经历3个阶段:开花决定、花的发端和花器官发育。首先,植物受到外界和内生信号的诱导,从营养生长转为生殖生长;其次,不同花诱导途径的信号相互作用,激活特异的花分生组织特征基因,使顶端分生组织向花序分生组织转变;最后,花序分生组织特征基因激活花器官特征基因的表达,从而激活器官建成基因的表达,最后形成花器官[36-37]。
根据时空表达模式(图2)、蛋白的功能特点和突变体表型(表1),本文将护颖发育相关基因分为3类:OsVIL2和SAD1属于开花决定基因;OsIG1、EG1和NSG1/LRG1属于花序分生组织特征基因;OsMADS1/LHS1、PAP2/OsMADS34、G1和AP2/EREBP(FZP、SNB、IDS1和MFS1)属于花器官特征基因。

图2 护颖发育相关基因的表达模式Fig.2 Expression patterns of genes related to sterile lemma development箭头表示顶端分生组织到花分生组织的发育进程。SAM:顶端分生组织;IM:花序分生组织;BM:枝梗分生组织;SM:小穗分生组织;FM:花分生组织。绿色矩形和红色矩形分别代表正向调控和负向调控,灰色矩形代表已确定或未知调控。矩形的宽度表示基因在相对应的器官或分生组织中表达。The arrow indicates the process of shoot apical meristem(SAM)to floral meristem(FM). IM:Inflorescence meristem;BM:Branch meristem;SM:Spikelet meristem;FM:Floral meristem. The green,red rectangles indicate positive,negative regulation and grey rectangles indicate determinacy or unknown regulation. The width of the rectangle indicates the expression domain of genes correspond to the different floral organs or different meristems.
2.1 开花决定基因
OsVIL2参与水稻春化途径,通过对靶基因的染色质修饰来调控开花时间[13]。转录组测序表明,OsVIL2突变体中OsIG1和G1的表达显著下调,表明OsVIL2是OsIG1和G1的上游基因[26]。
SAD1编码1个RNA聚合酶Ⅰ(PolⅠ)的亚基,而RNA聚合酶Ⅰ受到细胞分裂素(CK)的诱导[38]。与野生型相比较,sad1突变体中的细胞分裂素明显增多,这可能是由某种反馈机制调节导致[11]。有研究表明,miR156和miR172这2个microRNA在植物营养生长向生殖生长转变过程中起关键作用[39]。Li等[12]研究表明,SAD1正向调控miR172,负向调控miR156。
上述基因除了蛋白功能还具有开花决定基因的特征,而且osvil2和sad1突变体植株都表现为营养生长向生殖生长的过渡期大幅延长,且顶端分生组织的生长受到抑制,所以,我们认为OsVIL2和SAD1属于开花决定基因。
2.2 花序分生组织特征基因
OsIG1、EG1和NSG1/LRG1都在花序分生组织中高表达,而在顶端分生组织中不表达或低表达,所以,我们认为这3个基因都属于花序分生组织特征基因。Zhang等[14]发现,osig1突变体中的EG1表达水平降低,表明EG1是OsIG1的下游基因;osig1突变体中OsMADS1表达量升高,而Li等[15]研究的eg1突变体中OsMADS1表达量下降。根据Cai等[29]研究结果,eg1突变体2 mm以内幼穗中的OsMADS1表达量下调,而3~4 mm幼穗中的OsMADS1表达量下调不明显,说明EG1调控OsMADS1受到时空性的限制。总之,OsIG1、EG1、OsMADS1这3个基因是从上游到下游的调控关系。Xu等[10]通过qRT-PCR和原位杂交试验发现,lrg1突变体中G1基因的表达量明显下降,而OsMADS1表达量显著上升。这个结果表明,NSG1/LRG1正向调控G1,负向调控OsMADS1。Zhuang等[11]研究发现,NSG1/LRG1能直接与OsMADS1的启动子结合,然后再与TPR类(ASP1即OsTPR2)的共阻遏蛋白结合,通过下调OsMADS1所在染色质上组蛋白的乙酰化水平,抑制OsMADS1的表达。
2.3 花器官特征基因
OsMADS1、PAP2/OsMADS34、G1、AP2/EREBP(FZP、SNB、OsIDS1、MFS1)这4个基因/类基因在水稻小穗分化过程中的时间和空间表达特异性强(图2),属于花器官确定性基因。虽然这些基因分别调控相对应的花器官,但没有严格的分界线。从表达方式上来看,这些基因的表达是时空性的、局部连续的、部分重叠的,这些基因之间也互相调控。
OsMADS1虽然没有直接调控护颖的发育,但OsMADS1是小穗分生组织向小花分生组织过渡阶段的关键调控因子。Khanday等[40]研究发现,OsMADS1以直接减少PAP2/OsMADS34(穗分生抑制子)的表达和激活OsMADS55(花分生正向调控子)的方式促进小穗分生组织向小花分生组织过渡。Dai等[41]研究发现,OsMADS1通过抑制microRNA172的表达调控水稻内/外稃的发育。
PAP2/OsMADS34是正向调控小穗分生组织的特征基因,也是水稻花序结构和护颖特征的重要调节基因[9,42]。OsMADS1与PAP2都属于禾本科特有的LOFSEP亚族基因,而且这2个基因都参与小穗分生组织的发育过程,但又有分工上的不同。首先,PAP2的作用是启动小穗分生组织发育,并且参与副护颖和护颖的发育进程;其次,OsMADS1维持小穗分生组织发育,参与内稃、外稃以及内3轮的花器官发育。进一步研究表明,PAP2是OsMADS1的直接靶基因并被该基因负向调控[40]。已知pap2突变体的护颖变长,我们推测PAP2基因负向调控护颖建成基因表达。这个推断或许可以解释OsMADS1异位表达导致护颖变长的原因。
已知G1基因只在护颖和内稃中表达,G1基因功能的缺失导致护颖向外稃的同源转化,而其他性状不受影响,所以,G1基因是护颖的特征基因。Gao等[42]在pap2突变体中发现,G1基因的表达(qRT-PCR)并没有受到影响,认为G1与PAP2分别独立调控护颖的发育。然而,Liu等[22]发现,g1突变体中的PAP2基因表达(qRT-PCR)显著降低,认为G1调控PAP2的表达。所以,需要更多的试验证据来确定G1与PAP2之间的调控关系。
FZP、SNB、OsIDS1和MFS1这4个基因都具有AP2/ERF结构域,它们的突变都会影响小穗分生组织的确定性,而且都没有发育出护颖或护颖退化。fzp和snb突变体都形成更多的副护颖,并且具有明显的小穗分生组织和小花分生组织边界。fzp突变体在副护颖的中间形成腋生分生组织(axillary meristem,AM),而花分生组织被高次枝梗代替[35]。snb突变体在多重副护颖之上形成花器官,而且没有观察到fzp突变体副护颖中间的腋生分生组织。除此之外,在snb突变体中,FZP基因的转录水平没有明显变化。根据上述试验结果,Lee等[18]认为,FZP和SNB基因在水稻的小穗发育过程中分别独立表达,而且FZP基因行使功能的时期在SNB基因之前。Ren等[35]根据MFS1的小穗分生组织确定性丢失比SNB迟,推断MFS1基因行使功能在SNB基因之后。FZP和SNB基因在mfs1的突变体中表达量显著降低,护颖特征基因G1表达量减少,说明MFS1基因参与这些基因的表达调控。与mfs1突变体一样,G1基因在fzp突变体中的表达也显著降低,而FZP过表达植株的护颖形态伸长[17]。G1基因正常表达会抑制护颖伸长,G1基因表达显著降低则应该表现为长护颖的形态,然而fzp、snb、osids1、mfs1这些突变体都没有发育出护颖或护颖退化。我们推测,AP2/EREBP基因没有直接调控G1的表达,而是直接正向调控其下游的器官建成基因。已有研究[43-45]表明,AP2亚族的基因具有维持植物分生组织分生能力的作用,EREBP亚族也参与植物激素应答、胁迫应答和发育调节[46-47]。Ren等[35]研究表明,FZP通过控制细胞增殖和细胞增大来调节谷物大小。至于fzp和mfs1突变体中G1基因表达为什么会降低,我们推测是由G1基因与护颖器官建成基因的一种反馈调节机制造成的。
AP2家族成员是miR172的靶基因[48-54],miR172的过表达植株表现为与snbosids1双突变体类似,但是有更严重的表型缺陷[19],这意味着miR172通过抑制AP2类基因表达方式参与水稻花发育的调控。
3 水稻护颖发育相关基因之间可能的调控模式
综上所述,我们总结了一个水稻护颖发育相关基因的调控模式(图3)。首先,OsVIL2受到外界或内生信号的诱导,激活植物特异转录因子OsIG1。同时,OsSAD1也受到外界或内生信号的诱导促进miR172的积累。其次,OsIG1激活质体靶向脂肪酶基因EG1,并参与茉莉酸(JA)生物合成。NSG1/LRG1是OsMADS1和G1的上游基因,该基因直接抑制OsMADS1表达,并促进G1表达,NSG1/LRG1与其他花序分生组织特征基因的互作未知。EG1通过直接或间接作用进一步调控OsMADS1表达。按照经典的被子植物花发育ABC模型[55]分类,G1、PAP2、OsMADS1和AP2/EREBP都属于A类基因,这些基因直接或间接调控护颖建成基因的表达,最终形成水稻护颖器官。目前,还没有水稻护颖相关建成基因(可能与细胞分裂和激素调节有关)的报道。根据已有研究结果,我们推断AP2/EREBP类基因正向调控护颖建成基因的表达,而G1和PAP2抑制护颖建成基因的表达,从而维持正常的护颖表型。最后,OsMADS1开始表达,并且抑制PAP2在外稃、内稃中表达。而SAD1促进miR172的积累,miR172又抑制AP2/EREBP类基因的表达,同时miR172又受到OsMADS1的抑制。
ASP1编码1个转录共抑制子,而且asp1突变体在营养生长到生殖生长阶段具有多效性表型,我们认为,ASP1与miR172具有类似的作用,它们都是调节水稻花发育和生理进程的调节子。目前已知ASP1属于TPR(TOPLESS-RELATED)类蛋白[30],而TPR类蛋白能与具有EAR结构域的蛋白结合[56]。Zhuang等[11]研究证实,ASP1、NSG1/LRG1(具有EAR结构域)和HDAC(组蛋白脱乙酰基酶)结合成复合体共同抑制OsMADS1的表达。
4 研究展望
关于水稻护颖发育的分子机制,仍然需要更多的试验数据来阐明。我们认为有以下几个方面值得进一步探索:1)NSG1/LRG1与OsIG1、EG1之间是互作关系还是单独执行功能;2)EG1与OsMADS1之间是否还有其他基因参与调控;3)需要更多的试验证据来确定G1和PAP2之间的关系;4)哪些内生信号直接激活了G1的表达;5)G1调控的下游基因有哪些。鉴于目前组学相关技术的飞速发展,水稻护颖发育相关基因可通过转录组、蛋白组、代谢组的联合分析方法,快速明确这些基因之间的调控关系;再通过蛋白互作试验或转录表达试验进一步验证某2个基因之间的具体关系。水稻护颖发育分子机制的阐明,不但对揭示水稻花发育机制具有重要的理论意义,而且通过护颖相关基因的编辑或聚合,可以增加水稻的每穗粒数、千粒重、枝梗数等重要的产量性状,为水稻增产提供一条新的思路。
猜你喜欢
亚热带农业研究(2022年1期)2022-08-08
蔬菜(2021年7期)2021-11-27
流行色(2021年8期)2021-11-09
中国生殖健康(2020年7期)2021-01-18
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
中国生殖健康(2020年6期)2020-02-01
汉语世界(The World of Chinese)(2019年5期)2019-11-11
少儿科学周刊·少年版(2018年12期)2018-01-26
科学中国人(2017年36期)2017-06-09
