
妊娠中后期饲粮添加氮氨甲酰谷氨酸对新生子代滩羊生长发育、脂肪沉积相关基因表达以及母羊血清生化指标的影响
2021-05-20 02:21李荫柱牛文智张桂杰
动物营养学报 2021年4期
钟 锐 李荫柱 刘 博 牛文智 张桂杰*
(1.宁夏大学动物科学系,银川 2750021;2.吴忠市红寺堡区天源良种羊繁育养殖有限公司,吴忠 2751900)
反刍动物脂肪组织形成主要发生在胚胎期[1]。脂肪沉积包括脂肪细胞增殖和肥大,脂肪细胞增殖主要通过妊娠中后期肌内脂肪发育开始时增加脂肪沉积位点来实现,而脂肪细胞肥大主要通过育肥阶段营养供应实现[2]。妊娠期母体营养水平与胚胎发育及其血管生成密切相关,因此妊娠中后期母体营养供给模式可改变胎儿脂肪发育与沉积[3-5],直接影响到成年子代胴体品质和肉品质。随着动物营养学研究的深入,业内人士越来越重视功能性氨基酸在动物生产及营养调控方面的作用。氮氨甲酰谷氨酸(N-carbamyl glutamate,NCG)能有效激活内源性精氨酸(arginine,Arg)合成途径中关键酶——氨甲酰磷酸合成酶-Ⅰ(carbamoyl phosphate synthase-Ⅰ,CPS-Ⅰ),启动内源性Arg合成[6-9],Arg直接参与动物体内多胺和一氧化氮(NO)形成。血液中NO含量升高有助于血管扩张和血液流动,可增强胚胎与母体之间的联系,与胚胎期的脂肪发育启动密切相关。与Arg相比,NCG性质稳定、半衰期长,可提高母体胎盘的最外层及滋养层细胞数量、形态及活性,促进离子、糖、水及氨基酸进入胎儿,促进营养物质利用率并改善胎盘发育[10]。研究表明,在猪饲粮中添加适量的NCG可显著提升胚胎存活率,上调与胚胎着床相关因子的表达,并诱导胚胎和胎儿发育[11-12]。
本团队前期研究发现,饲粮添加0.10% NCG可以显著提高滩羊肌内脂肪含量,改善育肥期滩羊的体脂分配,显著提高肉品质[13-14]。但关于家畜妊娠阶段饲粮添加NCG是否调控新生子代生长发育和脂肪沉积,尚缺乏理论支持。因此,本试验旨在探讨妊娠中后期母体供给NCG对新生子代滩羊生长发育、不同部位脂肪沉积相关基因表达与母体血清生化指标的影响,以期为NCG调控母子一体化高效生产提供科学依据。
1 材料与方法
1.1 试验动物与饲粮
试验选用胎次、年龄和体重[(47.75±1.20) kg]相近的经产滩母羊80只,采用同期发情后进行人工授精配种,成功受孕63只。选择成功受孕的60只母羊进行试验。妊娠70 d按母羊体重随机分为2组,每组5个重复,每个重复6只羊。分娩结束时,每只妊娠母羊均产羔羊1只,共计60只。
试验基础饲粮参考NRC(2007)设计,其组成及营养水平见表1,对照组母羊饲喂基础饲粮,试验组在基础饲粮中添加0.10% NCG(NCG含量≥95%,水分含量≤3%,总砷含量≤5 mg/kg,铅含量≤10 mg/kg)。

表1 基础饲粮组成及营养水平(干物质基础)
1.2 饲养管理
试验羊打耳号后饲养在通风良好的半开放式羊舍内,妊娠前期统一进行饲喂,妊娠中后期进行分群饲喂。预试期7 d,正试期至分娩结束。每天08:00和16:00分2次饲喂,自由饮水。按羊场操作程序进行消毒、驱虫和免疫。每天根据饲粮剩余量调整饲喂量,保证饲粮剩余量<5%,直至分娩结束。
1.3 样品采集与处理
每个组中随机挑选母羊5只,共计10只,妊娠结束后,立即将新生羔羊进行屠宰,擦干新生羔羊体外胎液后测定初生重及体尺指标。母羊分娩后静脉采血5 mL,收集血清,置于-20 ℃冰箱保存,待测。分别采集新生羔羊肾周脂肪(肾脂)、肠系膜脂肪(肠脂)、腹部皮下脂肪(皮脂)和尾部脂肪(尾脂)约10 g,因母羊妊娠结束时间并不统一,为防止样品RNA降解,样品置于液氮中保存,备用。
1.4 指标测定
1.4.1 新生子代滩羊的初生重与体尺
测定所有新生子代滩羊的初生重和体尺。
初生重:擦干新生羔羊体外胎液后称重。
体高:肩胛骨最高点到地面的垂直距离。
体长:肩端至坐骨结节末端距离。
胸深:鬐甲至胸骨下缘的垂直距离。
胸围:肩胛骨后缘绕胸1周的长度。
1.4.2 母羊血清生化指标
所选10只母羊妊娠结束后,静脉采血5 mL,置于离心管中,1 980×g、4 ℃离心15 min,收集血清,置于-20 ℃冰箱保存。采用全自动生化分析仪(TBA-40FR,日本东芝公司),通过比色法测定血清总蛋白(TP)、白蛋白(ALB)、球蛋白(GLB)、总胆固醇(CHO)、葡萄糖(GLU)、尿素氮(UN)含量,试剂盒购自南京建成生物工程研究所,按照说明书要求进行操作。
1.4.3 新生子代滩羊不同部位脂肪组织中脂肪沉积相关基因表达
采用TRIzol法提取新生子代滩羊肾脂、肠脂、皮脂和尾脂的总RNA,取少量RNA用1%琼脂糖凝胶电泳验证。参照TIAN Script RT Kit反转录试剂盒说明书操作获得cDNA,并以此为模板,采用实时荧光定量PCR方法测定脂蛋白脂酶(LPL)、脂肪酸合成酶(FAS)、硬脂酰辅酶A去饱和酶(SCD)和葡萄糖转运蛋白4(GLUT4)mRNA相对表达水平。引物采用Primer 5.0软件设计,PCR引物序列见表2。以β-肌动蛋白(β-actin)为内参,在ABI7500荧光定量PCR仪上进行实时荧光定量PCR反应。根据目的基因和β-actin的阈值循环(Ct),采用2-△△Ct法计算样品中各脂肪沉积相关基因mRNA相对表达水平[15]。

表2 PCR引物序列
1.5 数据统计方法
试验数据用Excel 2010软件进行初步整理计算,采用SAS 8.2 TTEST过程统计分析,组间差异利用t检验进行比较,P<0.05表示差异显著,0.05≤P<0.10表示有显著趋势。
2 结 果
2.1 NCG对新生子代滩羊初生重及体尺的影响
由表3可见,与对照组相比,试验组新生子代滩羊初生重显著上升了6.1%(P<0.05),体长和体高有上升趋势(0.05≤P<0.10),胸围和胸深无显著差异(P>0.05)。
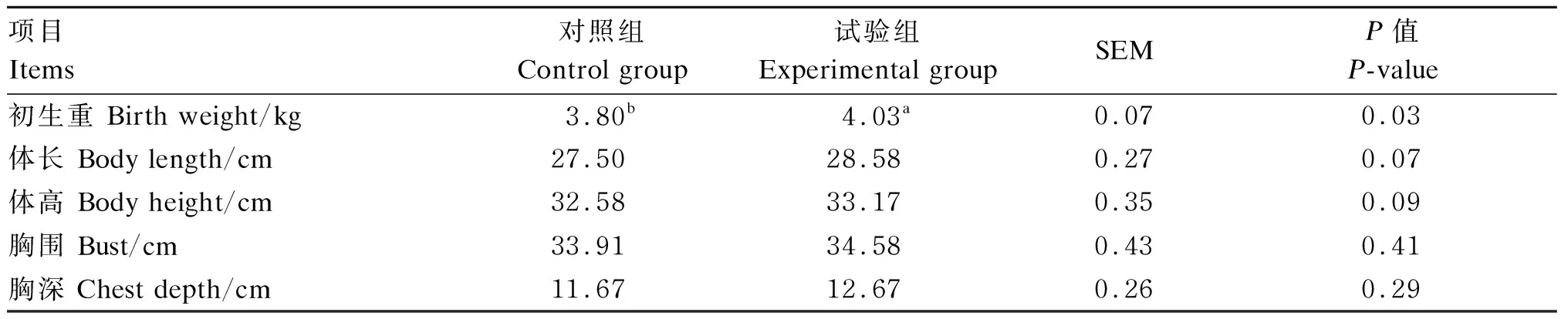
表3 NCG对新生子代滩羊初生重及体尺的影响
2.2 NCG对新生子代滩羊不同部位脂肪组织中脂肪沉积相关基因mRNA表达的影响
由表4可见,与对照组相比,试验组新生子代滩羊肾脂中LPLmRNA相对表达水平显著提高了18.6%(P<0.05),而肾脂中GLUT4 mRNA相对表达水平显著下降了13.3%(P<0.05);试验组新生子代滩羊肠脂中LPLmRNA相对表达水平有降低趋势(0.05≤P<0.10);新生子代滩羊肠脂、肾脂和尾脂中SCDmRNA相对表达水平有上升趋势(0.05≤P<0.10),皮脂中SCDmRNA相对表达水平无显著差异(P>0.05);新生子代滩羊肠脂、肾脂、皮脂和尾脂中FASmRNA相对表达水平无显著差异(P>0.05)。

表4 NCG对新生子代滩羊不同部位脂肪组织中脂肪沉积相关基因mRNA表达的影响
2.3 妊娠前、后母羊体重和日采食量
由表5可见,对照组与试验组妊娠前、后母羊体重和日采食量均无显著差异(P>0.05)。

表5 妊娠前、后母羊体重和日采食量
2.4 NCG对妊娠母羊血清生化指标的影响
由表6可见,与对照组相比,试验组母羊血清TP和GLB含量有上升趋势(0.05≤P<0.10),血清CHO、GLU和UN含量有下降趋势(0.05≤P<0.10)。

表6 NCG对妊娠母羊血清生化指标的影响
3 讨 论
3.1 NCG对新生子代滩羊初生重及体尺的影响
动物出生体重及体尺是反映其生长发育状况的重要指标[10,13]。胎儿发育依靠血液从母体内获得大量营养物质来满足自身需要,母体营养不良会导致胎儿生长发育受阻[16]。因此妊娠期母羊的营养调控至关重要。NCG能激活CPS-Ⅰ及二氢吡咯-5羧酸合成酶(P5CS),从而促进内源性Arg的合成。Arg是动物生产中条件性必需氨基酸且积极参与胚胎、胎盘及胎儿的生长发育[6,17-18]。大量研究表明,在妊娠母猪饲粮中添加不同水平NCG或Arg在改善母猪繁殖性能的同时可以提高窝产活仔数和初生窝重,保证胎儿的存活、生长和发育[19-20]。本研究发现,NCG显著提高了新生子代滩羊的初生重,而且体高和体长均有上升趋势,可能是饲粮中添加NCG提高母体血液和子宫液中Arg浓度,进而促进胎盘NO和多胺的合成[10,21]。NO含量增加可促进血管内皮生长因子(vascular endothelial growth factor,VEGF)的产生,诱导血管舒张并促进血流速度,多胺刺激血管生成、胚胎发育和胎盘生长,提高胎盘对胎儿营养物质供应[22-23]。此外,NCG的补充还可影响VEGF中miR-15b、miR-222和脐静脉内皮表面一氧化氮合酶(NOS)基因的表达,从而改善脐静脉功能和容积,使胎儿从母体获得更多的营养和氧气[22-25],进而改善胎儿生长发育。
3.2 NCG对新生子代滩羊不同部位脂肪组织中脂肪沉积相关基因表达的影响
脂肪的合成主要有2种方式,其一为小肠黏膜细胞由甘油单酸酯与脂肪酸合成脂肪,其二为肝细胞与脂肪细胞由甘油二酸酯合成脂肪,然后通过血液循环运输到肌肉等不同组织中沉积[26]。体脂的沉积是动物贮存能量的主要方式,其沉积量取决于脂肪合成代谢与分解代谢速度的高低[27]。反刍动物脂肪组织的形成多发生在妊娠中后期[4],而肠脂、肾脂、皮脂和尾脂组织过度发育会导致脂肪过度累积,影响滩羊的胴体品质。探究与脂肪沉积相关的候选基因和分子机制已成为脂肪沉积研究中的热点内容,不增加或降低无效脂肪的形成并增加脂肪内风味物质的沉积,才能改善胴体品质[28]。研究表明,LPL、FAS、SCD和GLUT4等在脂肪代谢调节过程中起着关键作用[29-31]。
LPL是脂质代谢和转运的关键酶,主要在脂肪组织、心脏肌肉和巨噬细胞中表达[32-33],其活性的高低反映脂肪的蓄积程度,对组织的脂肪的沉积起负向调控作用。LPL能够使动物机体内甘油三酯降解为甘油和游离脂肪酸,以供机体组织储存和利用。LPL可调节肌肉和脂肪甘油三酯的分配,是决定脂肪细胞大小的重要因素。LPL基因也通过调节不同组织内脂蛋白的沉积而影响胴体品质[34]。本研究中,LPLmRNA相对表达水平在肾脂中显著上升,抑制肾周脂囤积,与Bonnet等[35]研究结果相一致。这说明妊娠中后期滩羊母羊饲粮中添加NCG能够影响新生子代羔羊LPL的表达,可调控羔羊的脂肪分配,对提高新生子代滩羊的胴体品质有积极作用。
FAS是合成动物内源性脂肪酸和脂质积累的关键酶,在靶组织中参与脂质代谢,可改变脂肪酸生物合成速率。主要分布于高脂肪代谢和对激素敏感的组织中,可催化乙酰辅酶A和丙二酸单酰辅酶A用以长链脂肪酸的合成[36-37]。因此,FAS表达可调控脂肪酸合成速度,其活性与胴体脂肪含量呈正相关[34]。本研究中,NCG没有提高新生子代滩羊肠脂、肾脂、皮脂和尾脂中FASmRNA相对表达水平。这说明妊娠中后期滩羊母羊饲粮中添加NCG没有增加新生子代滩羊肠脂、肾脂、皮脂和尾脂长链脂肪酸合成量,对肠脂、肾脂、皮脂和尾脂中脂质积累物无影响,新生子代滩羊的胴体品质未受影响。
SCD是一种定位于内质网的跨膜蛋白,是合成单不饱和脂肪酸的关键酶,可在脂肪酸C9位置引入不饱和双键,将饱和脂肪酸转化成不饱和脂肪酸[38],影响着羊肉大理石花纹,脂肪柔软度及羊肉内风味物质的沉积等[36]。在哺乳动物中,SCD主要在肝脏、脂肪等脂肪生成的组织中表达,其次在肾脏、肺脏等组织中表达,其对肝脏脂肪的沉积起着重要作用[38]。本研究结果表明,NCG可提高新生子代滩羊肠脂、肾脂和尾脂中SCDmRNA相对表达水平。这说明妊娠中后期滩羊母羊饲粮中添加NCG会诱导子代滩羊SCD的表达。
GLUT4在哺乳动物GLU跨膜转运过程中起重要作用,对调节能量平衡和调控脂肪沉积作用显著[39]。GLUT4作为胰岛素中介导信号转导的关键因子,主要分布在脂肪组织细胞,这些细胞在胰岛素刺激下GLU摄入量迅速升高,其表达不足会使胰岛素敏感性降低,导致对GLU的利用减少[28,35,40]。本研究通过对滩羊GLUT4在不同脂肪组织中mRNA表达的研究发现,NCG使新生子代滩羊肾脂GLUT4 mRNA相对表达水平显著下降,可能是妊娠中后期滩羊母羊饲粮中添加NCG可降低子代滩羊肾脏周围脂肪组织细胞对GLU的吸收利用,进而抑制子代滩羊肾脏周围脂肪的生成,对改善羔羊体况发育起积极作用。
3.3 NCG对母羊血清生化指标的影响
血清生化指标是反映动物机体免疫功能、酶活性及脏器功能完整性的重要指标[41]。血清TP含量的高低反映了蛋白质的代谢水平,TP含量变高,有利于促进动物生长,提高饲料转化效率。血清UN含量是衡量机体内蛋白质代谢和氨基酸平衡的重要指标,蛋白质代谢良好则血清UN含量降低,相反饲粮中1种或多种氨基酸缺乏或过量时会造成血清UN含量的上升[42-43]。研究表明,瘤胃上皮细胞和十二指肠细胞具有利用NCG进行尿素再循环的能力[44],此外,饲粮中添加NCG能有效降低哺乳仔猪及断奶仔猪血清UN含量[45]。山羊羔羊灌服适量NCG可提高血清TP、ALB和Arg含量[9]。Arg可提高泌乳奶牛对蛋白质的利用,增加血清TP含量[10]。GLU是动物机体糖代谢平衡的重要指标,GLU含量的恒定可优先满足大脑、胎儿和泌乳等需求。GLU含量的动态平衡是依靠GLU来源和去路的协调来维持,动物在采食消化吸收期间,由肠道吸收大量GLU,其去路主要为各种组织中分解供能或转变为脂肪储存[46],此外,母畜妊娠胎儿时必须利用GLU,泌乳时需大量的GLU合成乳糖[47]。本研究中,妊娠中后期滩羊母羊饲粮中添加NCG后,妊娠母羊血清TP含量增高,血清UN、GLU含量降低,且均处于正常范围内,可能是由于妊娠期母羊饲粮添加NCG促进母羊体内蛋白质的合成效率,同时激活CPS-Ⅰ将体内多余的氨化为尿素,并提供更多GLU满足胎儿和泌乳需求。
4 结 论
妊娠中后期滩母羊饲粮中添加0.10% NCG可提高新生子代羔羊初生重,改善生长发育,调控脂肪沉积相关基因LPL、SCD和GLUT4的表达。
猜你喜欢
动物营养学报(2022年8期)2022-09-01
美食(2021年11期)2021-12-04
中国饲料(2021年17期)2021-11-02
中国饲料(2021年17期)2021-11-02
畜牧兽医科技信息(2018年12期)2018-02-14
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
广西林业科学(2016年3期)2016-03-16
动物营养学报(2015年9期)2016-01-07
