
亚精胺对盐胁迫下辽东栎生理特性的影响
2021-05-19 03:22范秀娟
河北林业科技 2021年1期
范秀娟
(山西省国有林场和种苗工作总站,山西 太原030012)
施用化学肥料是促进植物快速生长的有效措施,但是施用不当会引起土壤盐渍化、土壤板结等一系列问题[1]。土壤盐渍化是限制农林产业发展的重要瓶颈,由于改良困难并且耗时长,土壤改良难以快速达到预期目标,因此探寻植物耐盐机理,选择和培育抗盐性强的植物种类是面对土壤盐渍化最有效的途径[2]。辽东栎(Quercus wutaishanica)是我国北方地区阔叶混交林重要组成树种,很多地区都有大量的人工栽培林,并且也较多的应用于城市园林建设中。为适应造林工作的需要,山西省很多苗圃也大量培育辽东栎苗木[3]。育苗实践中,由于连年重茬栽培和不恰当的施肥技术,部分苗圃土壤已经发生了次生盐渍化现象,严重影响了辽东栎的优质壮苗培育。鉴于此,研究亚精胺对盐胁迫下辽东栎生理特性的影响规律,对揭示亚精胺的抗盐调控机理和育苗实践中进行化学调控植物抗盐性具有重要理论与实践意义。亚精胺是一种多胺类物质,盐胁迫下,其可以作为植物的信号物质用于植物抗盐机制的形成[4],大量试验研究表明,外源亚精胺可以显著提高植物的抗盐性[5]。从机理上来看,外源亚精胺对植物盐胁迫的缓解作用主要与亚精胺作为γ-氨基丁酸(GABA)合成的前提物质来促进其合成以维持植物细胞膜系统的稳定,降低丙醛(MDA)含量,保证植物在盐胁迫条件下正常生理生化活动的进行并促进植物生长[6]。亚精胺作为一种缓解植物抗盐性的多胺类物质,在大豆[6]、桑树[2]等植物上进行了初步的研究工作,但是其对辽东栎抗逆生理的相关研究较少。本文通过分析外源亚精胺对辽东栎根系活力及MDA、可溶性糖、可溶性蛋白、游离脯氨酸含量的影响规律,探讨外源亚精胺对辽东栎的抗盐生理机制,为探索辽东栎耐盐栽培的新途径提供理论依据。
1 材料与方法
1.1 试验材料
试验于2020 年4 月至2020 年7 月在山西省国有林场和种苗工作站试验苗圃内进行。试验所选辽东栎为2a 生实生苗。该苗木于2018 年3 月播种,2018 年7 月15 日移栽入长×宽×高为30cm×30cm×35cm 的栽植盆中,2018 年和2019 年在栽植盆中生长,并未发生干旱与水分胁迫,也未发生病虫危害。
1.2 试验设计
试验共设5 个处理,Y1 为空白对照,无盐胁迫和亚精胺处理;Y2 为盐胁迫对照,NaCl 浓度为150mmol/L;Y3 为盐胁迫+低浓度亚精胺:150mmol/L NaCl+0.60mmol/L 亚精胺;Y4 为盐胁迫+中浓度亚精胺:150mmol/L NaCl+1.2mmol/L 亚精胺;Y5 为盐胁迫+高浓度亚精胺:150mmol/L NaCl+1.8mmol/L亚精胺。其中,Y2、Y3、Y4、Y5 处理NaCl 用量为1500mL/株,其中4 月1 日浇灌栽植盆中500mL/株,5 月3 日浇灌1000mL/株。亚精胺处理分两次进行,第一次为4 月5~8 日每天18:00 后喷施,喷施量为100mL/株;第二次处理为5 月4~8 日每天18:00 后喷施,喷施量为100mL/株。空白对照分别在4 月1日和5 月3 日浇灌500mL/株和1000mL/株的蒸馏水;盐胁迫对照在4 月5~8 日、5月4~8 日期间分别喷施100mL/株的蒸馏水。每处理共计20 盆,3 次重复,随机区组排列,共计300 盆。整个试验期间,防止苗木出现旱涝灾害及病虫危害。
1.3 田间取样及生理指标测定
分别于4 月15 日、5 月1 日、5 月15 日、5 月31 日、6 月15 日到田间取样,叶片取样时,每处理选择3 株,将全部叶片放入液氮中贮存。根系取样时,将栽植盆倒立,所有盆内土壤全部倒出后用水冲洗干净根系后,用塑料袋保湿尽快带回实验室进行根系活力的测定。其中根系活力测定采用TTC 法[7]。将液氮中保存的叶片剪碎混合均匀,然后随机取样进行生理指标的测定,其中MDA 含量采用硫代巴比妥酸法测定[7],游离脯氨酸含量测定采用茚三酮法[7],可溶性蛋白含量测定采用考马斯亮蓝染色法[8],可溶性糖含量测定采用蒽酮法[8]。每项指标测定5 次,取平均值作为最后结果。
1.4 数据处理
图表制作及数据处理均使用Excel2010 版软件,方差分析使用DPS7.05 版软件。
2 结果与分析
2.1 亚精胺对盐胁迫下辽东栎MDA 含量的影响
由图1 可知:各处理辽东栎叶片内MDA 含量在整个试验期内表现出先升高后降低的变化,不同处理之间存在差异。整个试验期间,Y2 处理MDA含量始终为最高值,显著高于空白对照(Y1),表明盐胁迫对辽东栎膜系统造成了一定伤害,从而显著提高了MDA 含量。各亚精胺处理的MDA 含量均低于盐胁迫对照(Y2)处理,说明外源亚精胺对缓解盐胁迫下辽东栎膜系统的危害具有一定的作用,但是不同浓度的亚精胺缓解效果存在差异。Y4在整个试验期间MDA 含量分别比Y2 处理降低了27.78%、22.89%、24.10%、35.42%、29.66%,差异显著,表明该浓度的亚精胺对降低辽东栎盐胁迫危害具有显著作用;Y3 显著高于Y4 处理,Y3 在5月31 日显著低于Y2,其余时间Y3 均与Y2 之间无显著差异,说明低浓度的亚精胺对缓解辽东栎盐胁迫作用不大。5 月15 日、5 月31 日,Y4 显著低于Y5,4 月15 日、5 月1 日、6 月15 日Y4 与Y5 之间无显著差异,表明亚精胺浓度从1.2mmol/L 提高至1.8mmol/L 不会对辽东栎MDA 含量产生显著影响。
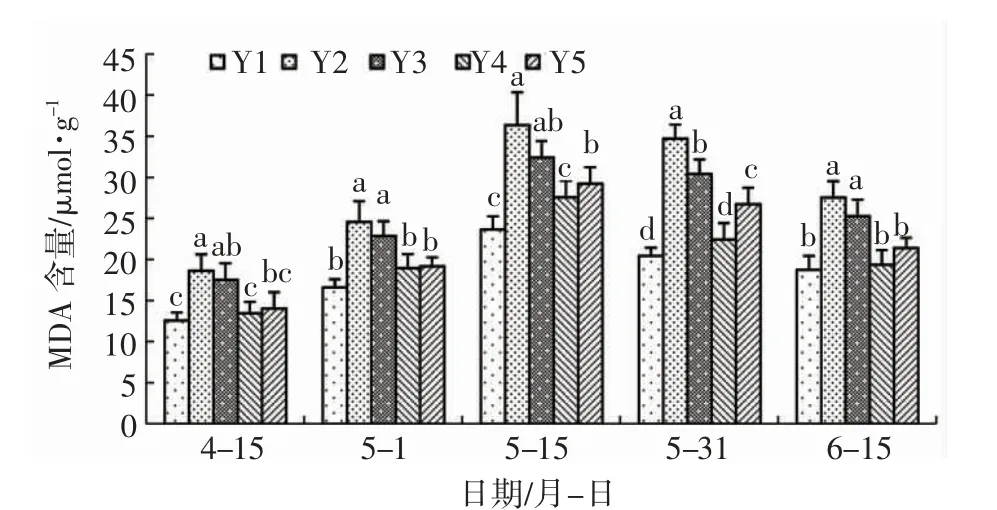
图1 亚精胺对盐胁迫下辽东栎MDA 含量的影响
2.2 亚精胺对盐胁迫下辽东栎可溶性糖含量的影响
由图2 可知:整个试验期间,盐胁迫处理与空白对照相比可溶性糖含量显著升高,表明盐胁迫显著诱导了辽东栎叶片内可溶性糖的积累。盐胁迫下,添加外源亚精胺与单独盐胁迫处理相比提高了辽东栎叶片内的可溶性糖含量,但是不同浓度的亚精胺对可溶性糖含量的影响存在差异。整个试验期间,Y4 可溶性糖含量分别比Y2 提高了20.65%、16.61%、18.27%、15.54%、23.83%,差异显著,表明该浓度的亚精胺显著提高了盐胁迫下辽东栎叶片内的可溶性糖含量;5 月1 日、5 月15 日、6 月15 日,Y4 显著高于Y3 处理,表明亚精胺浓度从0.6mmol/L 增加到1.2mmol/L 会显著增加辽东栎叶片内的可溶性糖含量;Y3 处理在整个试验期间可溶性糖含量均高于Y2,但两个处理之间无显著差异,表明该浓度的亚精胺对辽东栎叶片内可溶性糖含量影响较小;Y5 与Y4 之间无显著差异,说明亚精胺处理浓度由1.2mmol/L 基础上提高至1.8mmol/L 并未显著提高辽东栎叶片内可溶性糖含量。
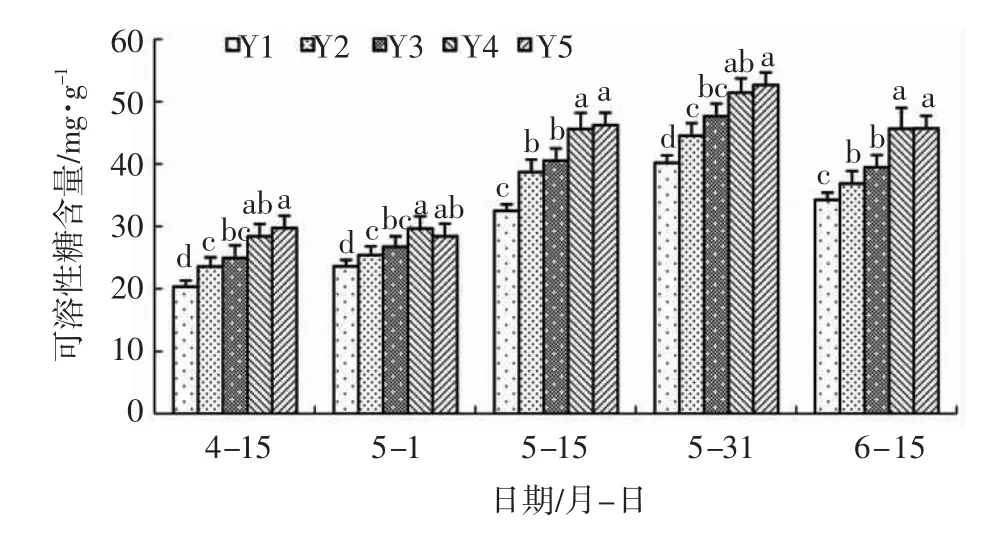
图2 亚精胺对盐胁迫下辽东栎可溶性糖含量的影响
2.3 亚精胺对盐胁迫下辽东栎可溶性蛋白含量的影响
由图3 可知:盐胁迫下,辽东栎叶片内可溶性蛋白含量在整个试验期间均显著高于空白对照,表明盐胁迫会诱导辽东栎叶片内可溶性蛋白积累。整个试验期间,Y4 分别比Y2 提高了29.32%、36.39%、47.15%、52.64%、40.34%,差异显著,表明1.2mmol/L浓度的亚精胺可以显著提高盐胁迫下辽东栎可溶性蛋白含量。Y3 分别低于Y4 处理1.23mg/g、1.59mg/g、3.62mg/g、4.42mg/g、2.21mg/g,差异显著,表明亚精胺浓度从Y3 提高至Y4 后显著增加了盐胁迫下辽东栎可溶性蛋白含量;4 月15 日至5 月15 日,Y2与Y3 之间无显著差异,表明0.6mmol/L 亚精胺在处理早期不会对盐胁迫下辽东栎可溶性蛋白含量产生显著影响;5 月31 日至6 月15 日,Y3 显著高于Y2 处理,表明该浓度的亚精胺在后期会显著提高盐胁迫下辽东栎可溶性蛋白含量。5 月31 日,Y5低于Y4 处理2.21mg/g,差异显著,表明亚精胺浓度从Y4 提高至Y5 在5 月31 日显著降低了可溶性蛋白含量;4 月15 日、5 月1 日、5 月15 日、6 月15 日Y4 与Y5 之间无显著差异,说明亚精胺浓度从Y4提高至Y5 后在这些时间段并未对辽东栎可溶性蛋白产生显著影响。
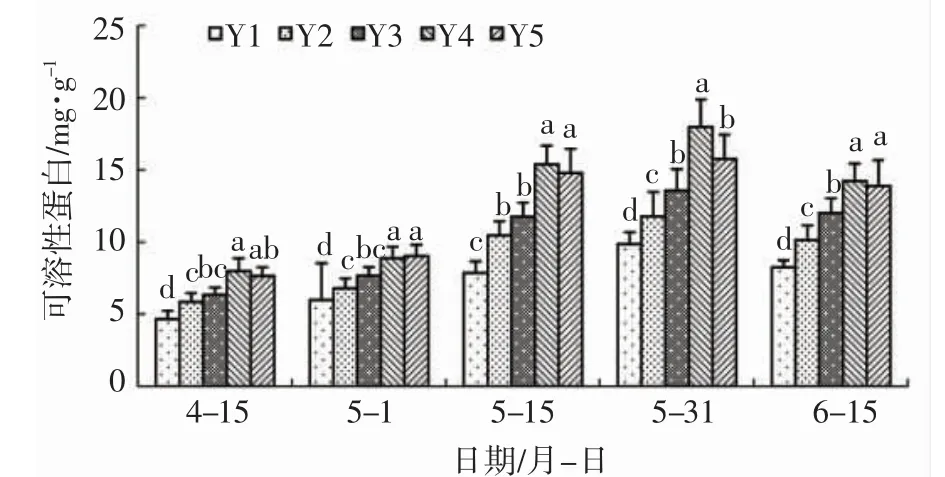
图3 亚精胺对盐胁迫下辽东栎可溶性蛋白含量的影响
2.4 亚精胺对盐胁迫下辽东栎游离脯氨酸含量的影响
由图4 可知:辽东栎游离脯氨酸含量在整个试验期间表现逐渐升高,不同处理对游离脯氨酸含量的影响存在差异。Y2 在整个试验期间均显著高于Y1,表明盐胁迫显著提高了辽东栎叶片内的游离脯氨酸含量。喷施亚精胺处理游离脯氨酸含量均高于盐胁迫处理,其中Y4 与Y2 相比分别提高了19.10%、25.75%、16.33%、22.17%、31.40%,差异显著,表明该浓度的亚精胺会显著提高辽东栎叶片内的游离脯氨酸含量。5 月1 日,Y3 与Y4 之间无显著差异,表明5 月1 日亚精胺浓度从Y3 降低至Y4 不会对辽东栎游离脯氨酸含量产生显著影响;Y3 与Y2 之间无显著差异,表明该时间段喷施低浓度的亚精胺对辽东栎游离脯氨酸含量影响较小;5 月15日至6 月15 日,Y3 显著低于Y4 处理,表明5 月15日后亚精胺浓度从Y3 增加至Y4 水平会导致辽东栎游离脯氨酸含量显著增加;5 月31 日至6 月15日,Y3 显著高于Y2 处理,表明低浓度的亚精胺处理在5 月31 日后可以显著提高盐胁迫下辽东栎的游离脯氨酸含量。Y5 与Y4 之间无显著差异,表明亚精胺浓度从Y4 提高至Y5 并未对辽东栎游离脯氨酸含量产生显著影响。
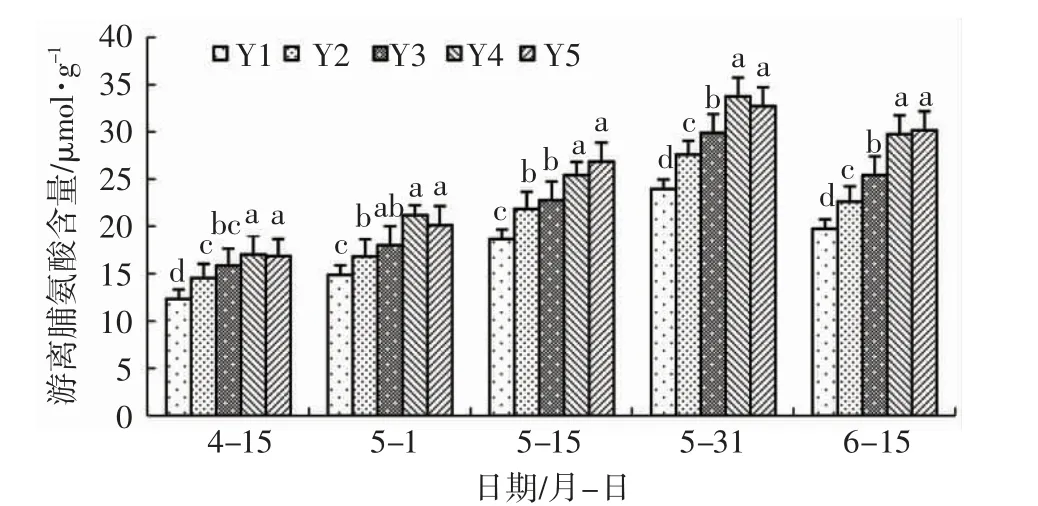
图4 亚精胺对盐胁迫下辽东栎游离脯氨酸含量的影响
2.5 亚精胺对盐胁迫下辽东栎根系活力的影响
由图5 可知:盐胁迫下,辽东栎根系活力显著降低,其中Y2 处理根系活力始终处于最低值,与空白对照相比差异显著,表明辽东栎根系活力在盐胁迫下会显著降低。亚精胺一定程度上会提高盐胁迫下辽东栎的根系活力,但是浓度不同对根系活力的影响也存在差异。整个试验期间,Y4 根系活力分别比Y2 处 理 提 高 了17.29% 、19.56% 、25.69% 、31.20%、28.81%,差异显著,表明该浓度的亚精胺会显著提高盐胁迫下辽东栎的根系活力;4 月15 日至5 月15 日,Y3 与Y4 处理之间无显著差异,表明亚精胺浓度从Y3 提高至Y4 在5 月15 日之前不会对根系活力产生显著影响;但5 月31 日至6 月15日,Y4 显著高于Y3,表明亚精胺浓度从0.6mmol/L增至1.2mmol/L 会显著提高辽东栎根系活力。Y5 与Y4之间无显著差异,表明亚精胺浓度增至1.5mmol/L不会显著提高辽东栎根系活力。
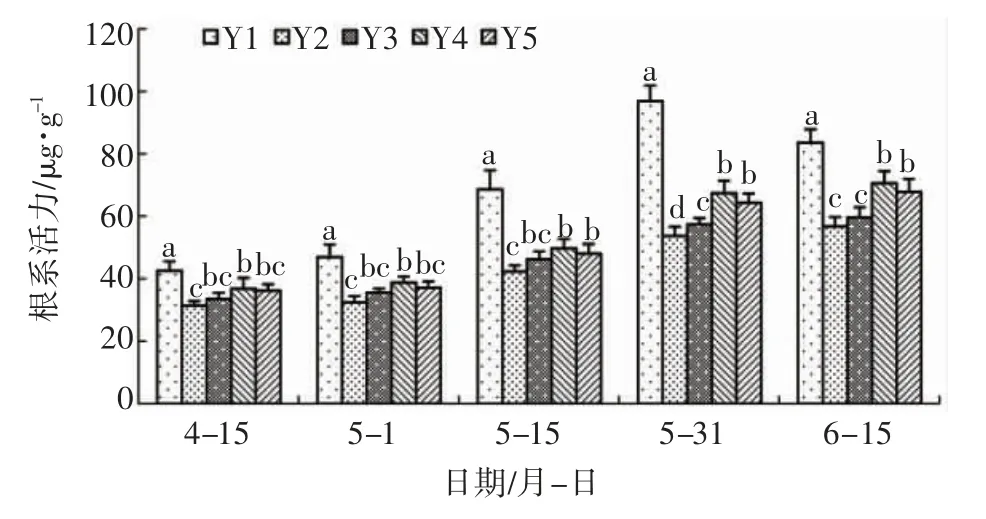
图5 亚精胺对盐胁迫下辽东栎根系活力的影响
3 讨论
MDA 是膜脂过氧化的主要产物,其含量的升高会导致植物体膜结构的完整性和稳定性受到破坏,影响植物的正常生理功能发挥,因此MDA 含量的变化可以作为判断逆境条件下膜系统受伤程度的指标[9]。从本试验结果来看,盐胁迫显著提高了辽东栎叶片内的MDA 含量,这也说明盐胁迫对其膜系统造成了一定的危害[10]。亚精胺处理与盐胁迫处理相比降低了辽东栎叶片内的MDA 含量,说明亚精胺对缓解辽东栎盐胁迫具有一定的作用,郭双[11]研究认为这与亚精胺可以提高植物体渗透调节物质含量,对膜系统起到保护作用有关。从不同浓度的亚精胺对辽东栎MDA 含量影响来看,中等浓度的亚精胺与高浓度和低浓度处理相比更有利于MDA含量的降低,这与赵东晓[2]的研究结果一致,说明亚精胺在缓解逆境胁迫中选择适宜的浓度也非常关键。可溶性糖和可溶性蛋白是植物体内重要的渗透调节物质,盐胁迫下其含量提高有利于维持细胞水分平衡[12],盐胁迫显著提高了辽东栎两种渗透调节物质含量,本试验结果与前人相关研究结果一致。外源亚精胺处理后,辽东栎可溶性糖、可溶性蛋白含量均表现出升高的变化,这与桑树[2]、黄瓜[13]的研究结果相似。王显瑞[14]研究认为这可能与亚精胺提高了蔗糖磷酸合成酶和淀粉合成酶的酶活性有关。游离脯氨酸含量的升高可以缓解植物盐胁迫造成的水分胁迫,适宜浓度的亚精胺处理可以显著提高盐胁迫下辽东栎叶片内的游离脯氨酸含量,这与刘球[15]的研究结果一致,李州[16]研究认为出现这种现象的原因与亚精胺可以提高脯氨酸代谢过程中的关键酶,如:Δ′-吡咯啉-5-羧酸合成酶(P5CS)、Δ′-吡咯啉-5-羧酸还原酶(P5CR)活性和分解代谢关键酶 ProDH 的活性促进氨基酸的代谢有关,也有部分研究者[17]认为亚精胺是游离脯氨酸的合成过程中的底物,外源亚精胺提高了底物含量,有利于促进游离脯氨酸的合成,从而提高其含量。盐胁迫降低了辽东栎根系活力,这与吴正东[18]的研究结果一致,而亚精胺一定程度上会缓解植物的盐胁迫危害,提高植物根系活力,促进植物根系生长,但是不同植物对亚精胺的浓度响应存在差异[19]。
4 结论
(1)盐胁迫显著提高了辽东栎叶片内MDA 含量,亚精胺显著降低了盐胁迫下辽东栎叶片内MDA含量;1.2mmol/L 亚精胺对降低丙二醛含量效果优于0.6mmol/L 和1.8mmol/L 处理。
(2)盐胁迫会显著提高辽东栎叶片内的渗透调节物质含量,亚精胺处理与盐胁迫对照相比显著提高了可溶性糖、可溶性蛋白、游离脯氨酸含量;1.2mmol/L 亚精胺促进渗透调节物质含量增加效果优于0.6mmol/L 处理;1.8mmol/L 与1.2mmol/L 亚精胺处理之间无显著差异。
(3)盐胁迫与空白对照相比显著降低了辽东栎根系活力,亚精胺有提高盐胁迫下辽东栎根系活力的作用,其中1.2mmol/L 亚精胺处理与盐胁迫对照相比差异显著。
(4)1.2mmol/L 亚精胺可以较好的缓解辽东栎盐胁迫。
猜你喜欢
中老年保健(2022年3期)2022-11-21
实用手外科杂志(2022年2期)2022-08-31
安徽化工(2021年3期)2021-05-29
科学导报·学术(2020年29期)2020-10-21
辽宁省博物馆馆刊(2020年0期)2020-08-13
棉花学报(2020年3期)2020-08-08
延边大学学报(社会科学版)(2020年2期)2020-03-25
美与时代·美术学刊(2019年9期)2019-11-29
科学与财富(2019年15期)2019-10-21
东坡赤壁诗词(2019年3期)2019-07-05
