
磷的生物地球化学循环研究进展
2021-05-12 05:35姜允斌郝记华季峻峰
高校地质学报 2021年2期
周 强,姜允斌,郝记华,季峻峰,李 伟*
1. 表生地球化学教育部重点实验室,南京大学 地球科学与工程学院,南京 210023;
2. 罗格斯大学 海洋与海岸科学学院,新泽西 08901-8520 美国
磷是重要的生命元素。它是核酸的磷酸盐酯基骨架,且在三磷酸腺苷(ATP)分子传递化学能的过程中起着至关重要的作用(Defforey et al.,2018)。作为重要的宏量营养元素,磷在维持全球粮食生产方面也发挥着不可替代的作用(Ashley et al., 2011)。然而,当今磷肥的粗放使用模式导致了水体富营养化的全球环境问题;更为严重的是,磷矿资源的加剧开采导致未来磷矿资源的短缺,并威胁粮食安全以及人类生存发展。因此,人类迫切需要从磷的生物地球化学循环过程中找到突破口来解决生存危机。而当前诸如“地球系统科学”等地球科学理论的发展和分析测试方法的进步,为研究磷的生物地球化学循环提供了更加开阔的视野和更加先进的手段。
21 世纪对于“全球变化”的争论,将人们的目光引向了地球圈层的相互作用,孕育了“地球系统科学”的理论,这是本世纪地球科学的重要进展(汪品先等,2018)。当前“地球系统科学”理论中两个要点最为突出:一是生命活动产生的气候效应,二是人类活动形成的地质营力(汪品先等,2018)。在陆地淡水环境中,磷是一个重要的限制性营养元素(Sterner, 2008);而在全球大多数海洋中,初级生产力在短时间内的主要限制性元素是氮,磷则是在较长的地质时间尺度上控制着海洋的初级生产力(Tyrrell, 1999; Bjerrum et al., 2002)。海洋浮游植物可以通过光合作用产生氧气,并将二氧化碳固定在有机物中。所以从某种意义上营养物质磷也影响着大气中的氧气和二氧化碳水平,因此地球表面营养物质磷的生物地球化学循环和全球气候变化是密切相关的(翁焕新,1999)。确立人类活动形成的地质营力,是对以往地球演化历史中塑造地球的主要地质营力:内动力地质作用(构造作用、岩浆作用、变质作用)和外动力地质作用(风化作用、剥蚀作用、搬运作用、沉积作用、成岩作用)的重要补充。目前人类活动正在以前所未有的速度和规模改变地球,日益成为现今最为关键的地质营力。这种地质营力已打破地球环境承载人类活动压力而不影响人类可持续发展的界限,这种界限被称为行星边界。当前九个行星边界中的四个已经被人类跨越,其中一个就是营养元素磷的生物地球化学循环(Steffen et al., 2015)。
然而,当前对磷的生物地球化学循环的研究存在一定的不足:(1)仅研究了磷在生物圈、水圈、表层岩石圈(地壳)等地球子系统中的循环,较少涉及大气圈和深层岩石圈对磷的生物地球化学循环的影响;(2)大多研究集中于现代地球系统中磷的生物地球化学循环,较少考虑到地史时期磷的演化对现今磷的生物地球化学循环模式的影响。在此,本文总结了磷元素的地球化学特征及其在整个地球历史时期各个阶段的演化,并以“地球系统科学”的视角了解人类活动对地球各圈层中磷赋存和转移的影响,为解决磷肥使用导致的环境问题和资源危机提供了新的见解和思路。
1 磷的地球化学
1.1 磷的特性
磷元素本身的化学特性决定了磷在自然界中的存在形式,进而决定着磷在生物地球化学循环中“流通”和“转化”的形式与分析测试的手段。磷是一种典型的非金属元素,是地壳中第11 丰富的元素。在戈尔德施密特元素地球化学分类中磷属于亲石元素,与O、 F、 Cl 的亲和力强(Goldschmidt,1937)。磷可以呈现-3、0、+3、+4、+5 多种价态,自然界中通常以+5 价的氧化态存在。因此,磷酸盐是自然条件下最主要的磷化物,此外,由于磷原子具有sp3型杂化轨道,所以它只能形成一种磷酸根离子PO43-(刘英俊等,1984)。已发现的23种磷同位素中只有31P 最为稳定,其它同位素都具有放射性,其中32P、33P 的半衰期在几十天内,其余的都为极不稳定的放射性同位素(Paytan et al.,2007)。自然界中磷主要以正磷酸根的形式存在于各种有机物和无机物中。因此在分析测试自然样品中磷含量的时候,通常要将样品中的磷消解为正磷酸根的形式。
磷存在于不同的地质作用过程中。内生作用中,磷在鲍文反应序列中的最早期以磷灰石的形式结晶分离出来。磷灰石是火成岩中最常见的副矿物,且火成岩中95%以上的磷都存在于磷灰石中(Smil,2000)。表生作用中,物理风化作用使岩石破碎,可产生细小的颗粒态磷灰石。化学风化作用使磷从磷灰石矿物中释放出来,产生溶解态的磷,并赋存在土壤孔隙水中。
1.2 含磷的天然有机化合物和无机矿物
磷在自然界中能够以无机化合物(正磷酸盐、焦磷酸盐、聚磷酸盐和含磷酸盐矿物)或有机化合物(磷酸单酯、磷酸二酯、膦酸酯、ATP、DNA)的形式存在(表1)。磷酸盐是自然条件下最主要的磷的化合物,因此自然界中磷主要以无机磷酸盐矿物和有机磷酸盐衍生物的形式存在于岩石和土壤中(Paytan et al., 2007)。磷的有机化合物中,ATP 是最重要的磷酸盐有机化合物。ATP 是自然界中最丰富的生物分子(Schlesinger,1997),它是所有已知生命形式的普遍能量“货币”。ATP 及其反应产物为所有生物体的活动提供或储存能量。这些反应发生在细胞内,负责一系列功能,包括新陈代谢、DNA 合成和细胞运动(Oelkerset al.,2008)。植酸又叫肌醇六磷酸,是肌醇磷酸类(含有1~6 个磷酸基的肌醇)的一种(Turner et al., 2002)。植酸是自然界肌醇磷酸类最普遍的存在形式,由植物在陆地生态系统中合成。在土壤中,植酸具有不易被生物分解利用的特性,且易与土壤络合,从而积累成为土壤中有机磷的主要赋存形式之一(Turner et al., 2002)。磷酸单酯、磷酸二酯及膦酸酯等磷的有机化合物也普遍存在于陆地土壤和海洋环境中(Paytan et al., 2007)。
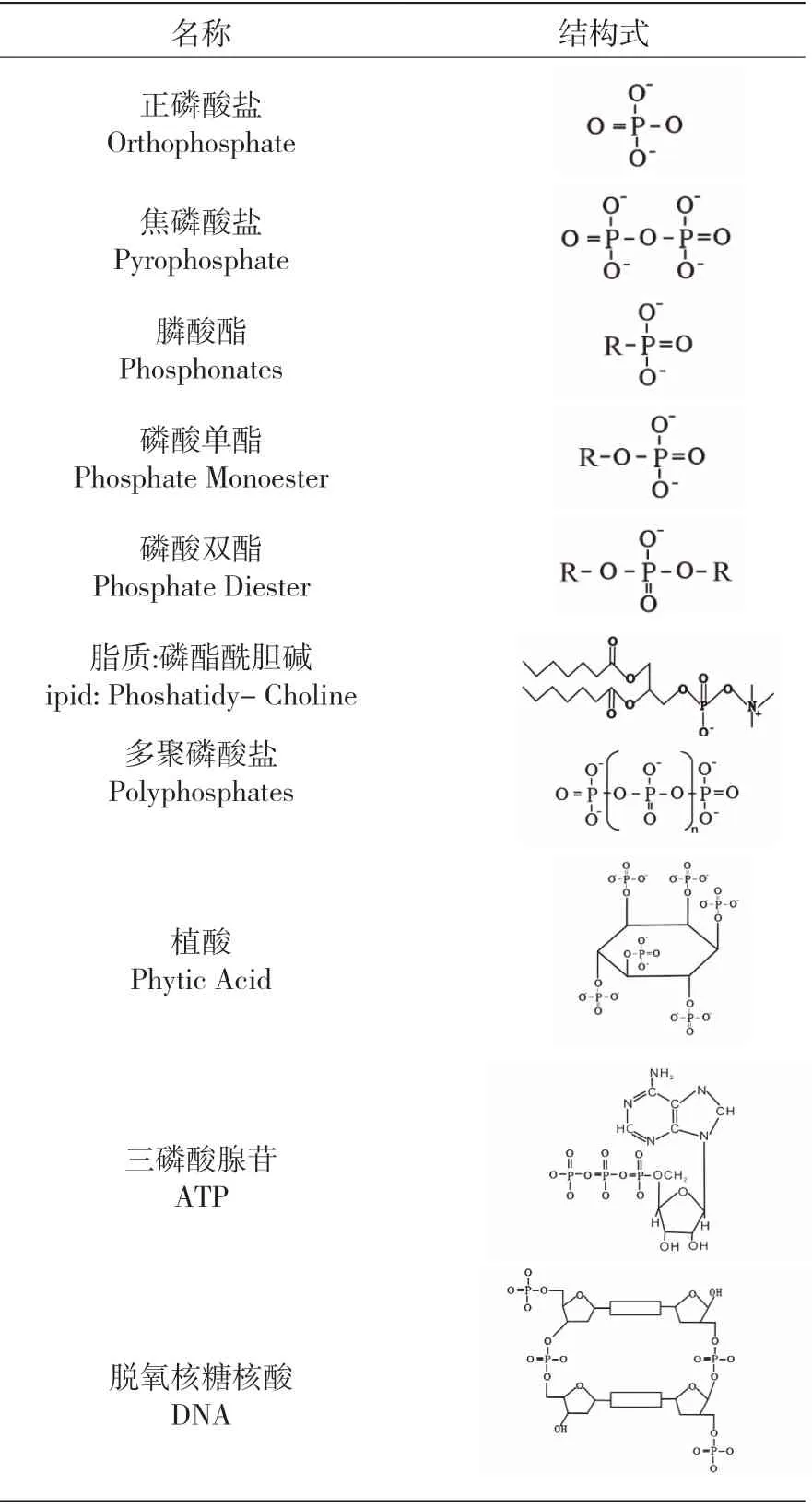
表1 生物学上重要的含磷化合物Table 1 Biologically important phosphorus compounds
磷灰石是地壳中最常见的天然含磷矿物,可在火成岩、变质岩、沉积和生物环境中形成(McClellan et al., 1969)。沉积岩中磷主要赋存在碳酸盐氟磷灰石CFA:Ca5(PO4, CO3, OH)3(OH, F)中。火成岩中的磷主要赋存在氟磷灰石Ca5(PO4)3F 中(Filippelli,2008;Manning,2008)。氟磷灰石是一种在成岩早期形成的副矿物,表现为与铁镁矿物相关联的微小的自形晶体(McConnell, 1973)。除磷灰石之外,自然界还有300 多种含有磷酸盐的矿物,但这部分矿物占地壳中总磷不足5%(Jahnke, 1992)。然而,也有研究表明,磷灰石并不是磷在地壳中赋存的最主要矿物相。由于磷可以取代硅酸盐矿物中的硅,且不同类型岩石硅酸盐矿物中的硅含量大不相同。因此,各种不同类型的硅酸盐矿物中磷的含量也各不相同,例如花岗岩中的长石普遍含有磷(Manning, 2008)。长石是大陆地壳岩石的主要矿物种类(40%~50%),花岗岩中的长石通常含有0.2%~0.3%且最多可含有1%的P2O5(Kontak et al.,1996)。根据简单的质量平衡计算,可估测出长石中的P2O5占地壳P2O5含量的50%~90%(Manning,2008)。除磷灰石外,在自然界中比较常见的含磷酸盐的无机化合物形成的矿物如表2 所示,其中蓝铁矿是自然界中除自生磷灰石外最普遍的沉积型自生磷矿物相(Egger et al., 2015)。富含重金属的磷酸盐矿物,如独居石、磷钇矿和磷稀土矿等广泛以微晶形式分布于火成岩和沉积岩中。火成岩中,独居石在花岗岩石中以副矿物的形式出现,在伟晶岩中以主矿物的形式出现;沉积岩中,独居石和磷钇矿的耐风化性使其在河流和海滩的沙粒中发生富集(Oelkers et al., 2008)。
1.3 不同地质体/天体中磷的含量及磷的地质富集
地球在形成之初,各种元素在地球中的分布是均一的,磷也不例外。磷矿的形成本质上是磷元素在地球不同圈层和储库之间运移、累积的结果。不同地质体/天体中磷的含量如表3 所示。相对于硅元素来说,球粒陨石中磷的丰度低于宇宙中磷的丰度(Anders, 1982)。球粒陨石中磷的含量为0.11%(Anderson, 1983),这一值和整体地球中的磷含量0.10%接近(黎彤,1976)。相对于整体地球,地壳和地幔中则更加富集磷。地幔中磷的含量为0.19%(Anderson, 1983),地壳中磷的含量为0.27%(黎彤,1976)。表明在原始地球圈层分异过程中,磷元素更偏向于进入地球的上部圈层。此外,相对于大陆下地壳,大陆上地壳中磷是亏损的(Wedepohl, 1995)。表明大陆上地壳中的磷在大陆风化的作用下不断流失,并成为地球各圈层中与磷的生物地球化学循环联系最密切的圈层。

表2 自然界中主要的含磷酸盐矿物Table 2 Phosphate minerals found in nature
全球的磷矿主要有原生磷矿床和次生磷矿床2大类。其中,原生磷矿床按成矿作用又包括沉积型磷矿床、岩浆型磷矿床和变质型磷矿床等3 种主要类型(Gerald et al., 2015; 薛珂等, 2019)。目前全球工业开采的所有磷矿石中,沉积岩型磷矿大约占85%(薛珂等, 2019)。沉积型磷矿主要形成于三个地史时期:震旦纪、寒武纪和泥盆纪。关于沉积型磷矿的成因,目前主要流行有三种假说:生物成因说(东野脉兴, 1985, 1992, 1996, 2001; 陈其英等,2000)、上升洋流说(Baturin, 1981)和交代成因说(叶连俊, 1986; Baturin, 1989)。上升洋流说认为上升洋流从深海底部带来大量富含磷的水体,到达浅海区域,由于压力、pH 值、CO2浓度等海水物理化学条件的变化,使得海水对磷酸盐的溶解度下降,进而促使磷质以无机沉淀的方式沉积下来形成磷块岩。生物成因说认为海水中生物吸收了海水中的磷质而大量繁殖,生物死亡后遗体下沉并分解,进而聚集形成磷块岩矿床。交代成因说认为海相磷块岩是由先前形成的碳酸盐岩被后期富含大量磷质的侵入海水交代形成的。笔者认为,大型沉积型磷矿的形成往往取决于生物、洋流和后期交代这三者的共同作用。因此,大型磷矿的形成一般会经历以下过程:(1)上升洋流带来的富磷水体到达浅海区,使浅海区域水体中磷含量骤然升高;(2)海水中生物吸收了海水中的磷质而大量繁殖;(3) 生物死亡后壳体的腐烂分解作用导致水体中pH 值减小、CO2浓度升高,水体溶解磷的能力增强,促进了磷质以化学沉淀的方式发生富集初步成矿;(4)富含大量磷质的海水侵入发生磷酸盐交代作用使磷质发生进一步富集,进而形成大型磷矿。然而不同的磷矿在形成过程中可能主要受控于其中一种或两种作用。例如,中国震旦纪陡山沱期磷块岩和梅树村期磷块岩的形成,都与生物的作用有着密不可分的联系(高磊, 2019);贵州开阳、瓮福含磷岩系存在着下部磷块岩对灯影组白云岩的交代和充填现象(叶联俊, 1986)。

表3 不同地质体/天体中磷的含量Table 3 Phosphorus concentrations in different geologic/celestial bodies
2 磷的地质演化史
在地球系统中,最早的岩石记录出现在38 或40 亿年之前(Nutman et al., 2006),此时原始地壳、原始海洋以及原始大气处于刚刚形成时期(图1)。原始地壳、原始海洋以及原始大气的形成为风化作用奠定了基础,使得陆地上的物质可以经地表河流搬运入海洋。目前观测到的最古老的沉积岩年龄是35~30 亿年(图1),而最早的生命也出现在35 亿年前的海洋中(图1;Schidlowski, 1988),与风化作用的启动时间基本吻合。磷是生命的必需元素。如果磷在风化作用刚开始时就从陆地被持续不断地搬运到海洋中,则使得当时的海洋中存在着可以被早期生命利用的磷,那么经过长时间的累积,磷应该大量出现在当时的沉积地层中。然而,目前尚无太古代沉积磷矿的报道,表明磷矿的形成在太古代地层中是受限制的,并且在中元古代典型的大陆边缘演替中也没有发现磷矿的存在(Holland et al., 2005)。能够进行光合作用的产氧光合生物蓝藻发现于27 亿年前(图1;Brocks et al., 1999),在此之前的地球大气圈和水圈是不存在氧气的。而在此之后的三亿年,光合作用产生的氧气不断在大气和表层海洋中累积,发生了著名的大氧化事件(图1;Kerr, 2005)。大氧化事件的直接产物就是条带状含铁建造(BIF)(图1)。Holland 等(1984)、Canfield(1998)等 认 为BIF的沉积导致矿物对磷的吸附明显提高。Bjerrum 等(2002)分析了BIF 中的磷含量,反演出32~19亿年前海洋中磷酸盐的浓度可能只有现在浓度的10%~25%。他们认为较低的磷浓度会显著降低光合作用和碳埋藏的速率。Reinhard 等(2017)通过对过去35 亿年细粒浅海硅质沉积物(页岩)的分析,反演出磷在地史时期海洋中浓度的变化(图2a)。他们同样认为太古宙缺氧、富铁的海洋中由于磷的清除作用较强,导致海洋中磷的低浓度,从而限制了海洋的初级生产力。

图1 地史时期与磷演化相关的重大地质事件(据汪品先等,2018修改)Fig. 1 Major geological events related to phosphorus evolution in the earth’s history

图2 沉积磷矿及细粒海相硅质碎屑沉积岩中磷含量随时间的变化(a, 据Reinhard et al., 2017修改);大气和海洋氧化还原状态的共同演化模型和海洋初级生产力限制性营养成分(b, 据Planavsky et al., 2010修改)Fig. 2 Sedimentary phosphorite occurrences and phosphorus concentrations of fine-grained, marine siliciclastic sedimentary rocks through time (a, modified after Reinhard et al., 2017); Model for the coevolution of atmospheric and oceanic redox state and limiting nutrients for marine primary productivity (b, modified after Planavsky et al., 2010)
然而在进化出蓝藻之前,初级生产是通过光合铁氧化作用来完成的(Canfield et al., 2006)。光合铁氧化过程中碳固定铁的化学计量为:C:Fe=1:4,而固定106 份碳需要1 份磷 (Jones et al., 2015;Widdel et al., 1993)。因此,1 份磷可以通过光合铁氧化作用氧化424 份铁。Holland 等(1984)估计出的太古宙深水中铁的浓度约为40~120 μM,Jones 等(2015)估算出当时深海中磷的浓度约为0.1~0.3 μM,这意味着磷可以通过光合铁氧化作用耗尽深海中的铁。此外,海水中的硅会与磷竞争在铁氧化物上的吸附位点,而太古宙和古元古代海洋中很可能含有大量溶解的二氧化硅(Konhauser et al., 2007; Planavsky et al., 2010)。因此,太古代时期海洋初级生产力和有机碳埋藏的主要限制因素,与其说是磷,不如说是铁(Konhauser et al.,2007; Planavsky et al., 2010; Jones et al., 2015)。
大氧化事件之后的很长一段时间,深层海水仍然是缺氧且富铁的,因此表层海水中磷的清除作用仍然较强,导致海洋中磷的浓度较低,同时也使得光合生物的产氧速率降低,延缓了进一步氧化事件的发生。大氧化事件并没有给深海带来氧气,反而形成了硫化氢海洋,使BIF 的沉积停滞于18 亿年前(图1;汪品先等, 2018)。硫化氢海洋的形成破坏了磷等生命元素在海洋中的循环途径,使海洋整体初级生产力受限,全球的氧化产生了十余亿年的停滞(汪品先等, 2018)。
元古宙末期,随着罗迪尼亚超级大陆的分解,全球沉积速率大增,大量有机碳的埋藏使得大气中温室气体含量降低,氧气含量增高,导致了几次“雪球地球”式大冰期的发生,并造成了大约7.5亿年前的第二次氧化事件(图1;Kaufman et al.,1997)。在第二次氧化事件之前的大部分时间里,富含自生磷的边缘海相沉积岩是非常罕见的,但在第二次氧化事件发生后的显生宙沉积序列中却很常见(图2a),这一明显区别表明此时期前后磷在海洋中的生物地球化学循环发生了根本性的转变。此时期后,海洋中磷浓度的升高,加上海洋环境条件的变化,导致磷的大规模沉积(Reinhard et al., 2017)。Reinhard 等(2017)、Planavsky 等(2010)等认为:海洋磷酸盐浓度在“雪球地球”事件时期达到峰值(图2b),是造成此事件及第二次氧化事件的主因,并为后生动物的繁盛提供了营养条件。在此,笔者将分为大氧化事件之后到第二次氧化事件之前(以铁氧化物吸附磷沉积为主)和第二次氧化事件之后(以自生磷沉积为主)来讨论海洋中的沉积磷汇的变化。
地球历史中大氧化事件之后到第二次氧化事件之前的大部分时间里,缺氧和富铁的环境一直主宰着海洋内部。在大氧化事件之后,表层海水被氧化,磷在海洋中的清除机制主要受控于磷与铁氧化物吸附共沉淀。在地球历史的绝大部分时间里,产氧光合生物圈一直以蓝藻为主(Knoll,2014),表层氧化、深层还原的富铁海洋会导致海洋表面生物可利用的磷通过与铁结合形成磷酸亚铁相(比如蓝铁矿)或与三价铁氧化物共同沉淀被耗尽。而在养分磷缺乏的条件下,表层海洋生物会表现出极高的C/P 值(White et al., 2006)。因此,在大氧化事件与第二次氧化事件期间的含铁海洋中(图3b),由于铁矿物对磷的清除,导致表层海洋磷的缺乏,浮游植物的高C/P 值,使得边缘海洋沉积物中磷的埋藏量大大降低(Reinhard et al., 2017)。
而在第二次氧化事件后氧含量充足的海洋—大气系统中(图3a),表层海水中营养物质磷的含量变得丰富,浮游植物C/P 值变低,使得磷大量沉积埋藏于边缘海相沉积物中(Reinhard et al.,2017)。这些边缘海相沉积物主要有三种:与有机质有关的磷(Organic P)、吸附在颗粒或氧化物上的磷(Iron-oxide-bound P)、以及CFA 中的磷,其中CFA 中的磷占50%以上(Delaney, 1998)。由于第二次氧化事件之后沉积磷矿的经济效应和科学价值,许多研究集中于阐明这些沉积磷的源汇、通量、地球化学形态及其埋藏速率等问题(Delaney,1998; Filippelli, 1997; Ruttenberg et al., 1993; Schenau et al., 2000; Schuffert et al., 1998)。
两亿年前的印支运动后(图1),逐渐形成了现今的海陆分布格局,这一时期也成为现今海洋磷循环的起点。在现今全球尺度磷的生物地球化学循环系统中,磷的最初来源是土壤发育过程中大陆风化作用产生的磷。大陆风化作用将岩石中的磷灰石以溶解态和颗粒态的形式释放出来(图4,过程1),通过河流、大气、地下水等方式搬运到海洋(图4,过程2)。大部分颗粒态的磷进入海洋后直接沉积在近岸海域的陆架上,约占总输入磷的90%(Filippelli, 2002)。而对海洋的磷循环具有主要贡献的是溶解态的磷,约占总输入磷的10%(William, 2013)。经河流搬运后直接沉淀在近岸海底颗粒态磷与参与海洋磷生物地球化学循环的溶解态磷最终都会转化为磷灰石(图4,过程3)。因此,磷灰石是磷在生物地球化学循环中源和汇之间的“桥梁”。磷灰石在高pH 值和存在大量缓冲物质的海水中溶解速度非常低,因此这些沉积在大陆边缘和深海中的磷灰石会稳定保存在地层中,直到板块俯冲作用最终使这部分磷重新暴露在陆地上(图4,过程4~6)(Filippelli,2002)。然而这一过程需要花费上亿年,这意味着在人类存在的时间尺度上,磷矿是一种不可再生资源。
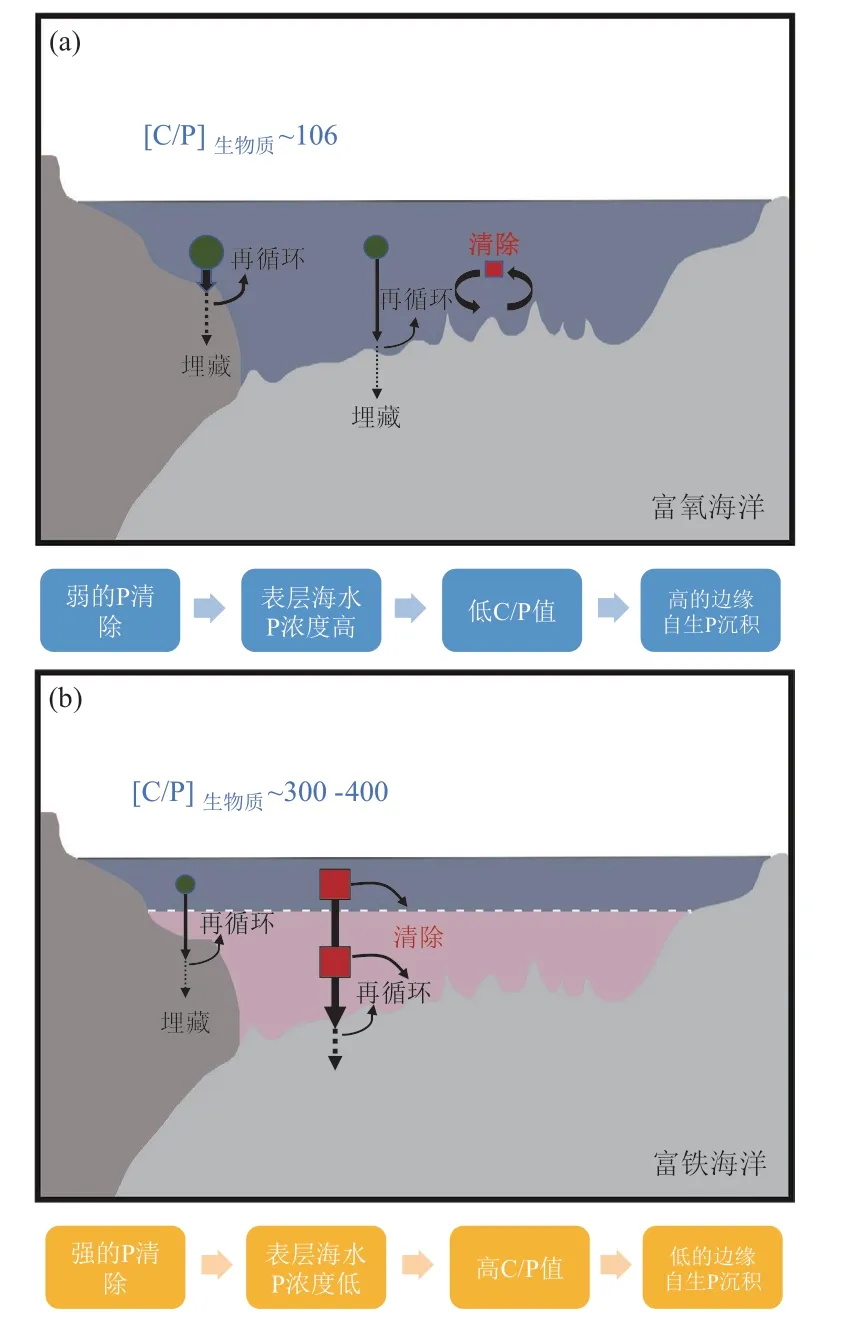
图3 不同海洋环境下磷循环的概念模型Fig. 3 Conceptual model for phosphorus cycling in different marine environments (modified after Reinhard et al., 2017)

图4 磷的地质旋回示意图Fig. 4 The geological cycle of phosphorus
3 全球磷循环
磷在陆地和海洋生态系统中的生物地球化学循环模式是磷在地质历史时期演化的结果,是地球系统中大气圈、水圈、岩石圈(地壳、地幔、地核)和生物圈(包括人类)相互作用、相互影响的动态过程。其中陆地生态系统(包括大气圈、陆地水圈、陆地岩石圈、陆地生物圈)和海洋生态系统(包括大气圈、海洋水圈、海洋岩石圈、海洋生物圈)是磷的生物地球化学循环中最主要的两个循环系统,二者之间相对独立又密不可分。
3.1 磷在陆地生态系统中的循环
陆地生态系统中,几乎所有的磷都以磷灰石或长石中的磷的形式赋存于岩石和土壤。随着土壤的发育,磷越来越多地从矿物中释放出来,并与其他物质结合在一起(图5)。而且随着时间的推移,土壤剖面中可用的磷的总量减少,因为土壤中的磷不断通过地表和地下径流流失(McDowell et al., 2001)。最终,土壤达到稳定状态,此时土壤磷被大量回收至海洋,而通过地表径流流失的磷则被土壤底部磷灰石风化形成的磷慢慢替代(Filippelli, 2008)。

图5 模拟土壤磷的地球化学组成随时间的变化Fig. 5 Modeled changes in soil phosphorus geochemistry over time (modified after Walker et al., 1976)
陆地生态系统的磷循环指磷以各种途径输入、输出生态系统以及磷在系统内部植物—土壤、与各营养级生物之间、生物体内和土壤内部的迁移转化(图6)。在此着重介绍磷以各种途径输入、输出生态系统以及磷在系统内部植物—土壤之间的转化。自然体系下,土壤中磷输入的途径有岩石矿物的风化、空气沉降和地表径流。而在人工生态系统中,施肥是磷素输入的最主要途径。岩石矿物风化是一个缓慢的过程,据统计通过岩石矿物风化进入生态系统的磷的通量为0.05~1.0 kg·hm-2·a-1(赵琼等, 2005)。空气沉降的磷酸盐以前一直被认为在系统磷循环中可以忽略,但是一些研究发现在很多生态系统中空气沉降是系统磷补给的一个重要来源(Chadwick et al., 1999)。通过空气沉降进入系统的磷的通量为0.07~1.7 kg·hm-2·a-1(Newman,1995)。因此凡是影响岩石风化与空气沉降的气候、地形、土壤发育阶段、植被覆盖情况、物候、周围环境条件等因素都对系统中磷的输入有影响。而地面径流对陆地系统磷输入的影响是不考虑的。自然陆地生态系统磷输出的途径有地表径流、沿土壤剖面的淋溶作用和风蚀。系统输出的磷酸盐多以颗粒物的形式随着地表径流被带出系统,进入地表水。在森林生态系统中随着地表径流和地下水渗流输出系统的可溶性无机磷一般不超过0.05 kg·hm-2·a-1(Likens et al.,1977),热带森林生态系统的可溶性磷输出一般不超过0.3 kg·hm-2·a-1(Bruijnzeel et al., 1991)。系统输出的磷主要是正磷酸盐、磷酸二酯和磷酸单酯(Solomon et al., 2000)。
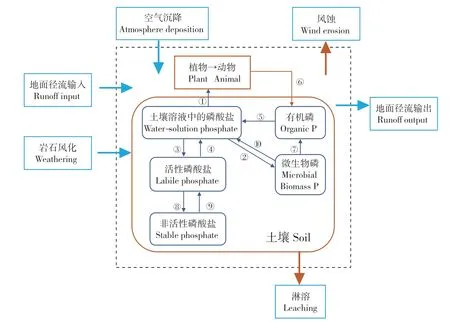
图6 陆地生态系统磷循环概述(据赵琼等, 2005修改)Fig. 6 Schematic overview of phosphorus cycle in terrestrial ecosystem
土壤中不同形态磷的转化和植物对磷的吸收利用一直是陆地生态系统磷循环的研究重点。磷在植物—土壤之间的转化过程,首先是因为植物通过呼吸作用释放的二氧化碳造成植物根部环境酸度的增加,使得结晶性差的含磷矿物迅速溶解,将磷释放到根部孔隙空间(Schlesinger,1997)。此外,植物根系分泌的乙酸盐、草酸盐、苹果酸盐和柠檬酸盐等低分子有机酸,会腐蚀土壤矿物质,将养分释放到土壤溶液中(Manning et al.,1992; Drever et al.,1997; Van Hees, 2002)。 然 后,被溶解的磷以无机磷的形式被植物吸收,并转化为有机形式。土壤总磷含量为0.5~2.0 mg·kg-1(Sanyal et al., 1991),通常以化合态或吸附态的正磷酸盐的形式存在,并以HPO42-和H2PO4-的形式被植物吸收。植物细胞中的磷酸根离子浓度为2~20μmol·L-1, 显著高于其在土壤溶液中的浓度, 因此植物对磷的吸收是一个主动吸收的过程(Schachtman et al., 1998)。此外,磷与铁、铝氧化物和氢氧化物的吸附结合过程在陆地生态系统中具有重要意义,且在土壤科学中得到了广泛的研究(Ruttenberg et al., 2014; Bennett et al., 2001;Guzman et al., 1994)。
3.2 磷在海洋生态系统中的循环
相比于大气和陆地,更多的研究集中于磷在海洋中的循环。磷输入海洋包括三个途径,分别为河流输入、大气输入和地下水输入。对于近大陆边缘环境,磷主要通过河流被运输到海水中(Ruttenberg et al., 2014);此外海底地下水在沿岸营养成分的循环中也起着重要作用(Moore et al., 2000; Slomp et al., 2004; Paytan et al., 2006);而大气沉积在远离大陆的海域中发挥更大的作用(Mahowald et al., 2008)。
首先,通过河流进入海洋的磷可大体分为颗粒有机磷(POP)、颗粒无机磷(PIP)、溶解有机磷(DOP)和溶解无机磷(DIP)(Benitez-Nelson,2000; Delaney, 1998)。PIP 占其中的绝大多数,其通量约为8.4~15 Tg/a;POP 约为0.9 Tg/a;DOP 约为0.2~0.5 Tg/a;DIP 约 为0.2 Tg/a(Paytan et al.,2007)。可见,随地表流进入海洋的溶解态磷(DOP与DIP)的通量约为1 Tg/a,加之潜在的可溶性磷(来自POP 与PIP,1~2 Tg/a),使溶解态磷的总通量约为2~3 Tg/a(Filippelli, 2002)。人类活动前,河流每年搬运9~17 Tg 磷到海洋中(Paytan et al., 2007)。在人类活动后河流每年向海洋中搬运的磷的总量约为21~31 Tg(Meybeck, 1982; Smil,2000)。相应的溶解态磷的总通量达到了4~6 Tg/a(Wallmann, 2010; Liu et al., 2008),约为人类活动前的两倍。
其次,与风成粉尘颗粒相关的气溶胶是海洋磷的另一个来源。大气输入的磷约占海洋磷输入总量的5%,其通量约为1 Tg/a(Duce et al.,1991)。这种通过大气输入的磷在远离海岸的区域更为重要(Mahowald et al., 2008),通常以有机和无机化合物的形式存在于气溶胶中,且有机和无机化合物比例大致相同(Ridame et al., 2002; Chen et al., 2007)。其中无机磷主要与铁氧化物结合,或与钙、镁、铝、铁等元素结合(Ridame et al., 2002; Bergametti et al.,1992),而有机磷的存在形式尚不清楚。
输入到海水中的磷以溶解态和颗粒态两种形式存在于整个水体中,它们可以通过海洋中磷的生物地球化学循环过程实现相互之间的转化(图7)。海洋中溶解态的磷包括无机磷(一般为可溶性正磷酸盐形式)、有机磷化合物和大分子胶体磷(Paytan et al., 2007)。其中最主要的是可溶性活 性 磷(SRP: Soluble Reactive Phosphorus), 主要由DIP(磷酸氢根离子和少量磷酸根离子,其相对丰度取决于pH 值)组成,但也可能包括其他一些容易水解的无机和有机磷(Benitez-Nelson,2000)。总磷减去SRP 为可溶性的非活性的磷(SNP:Soluble Nonreactive Phosphorus)。主要为DOP(蛋白质、碳水化合物和脂质),也包含无机聚磷酸盐(Paytan et al., 2007)。海水中的溶解态总磷约为90000~93000 Tg(William et al., 2013; Filippelli, 2002;Paytan et al., 2007),其中深层海水中溶解态磷的储量约为89900 Tg,表层约为3100 Tg(Paytan et al., 2007)。由于浮游植物的光合作用,大多数表层海水的磷酸盐浓度接近于零(Filippelli, 2002)。因此,海水中的DIP(主要为磷酸根离子,此处将DIP 与SRP 视为等同)在海洋中的深度剖面展示出表面耗竭和深层富集的特征趋势(Paytan et al.,2007)。在深海区域,由于下沉颗粒物的不断积累和再生,DIP 浓度也随着深层海水年龄的增大而增长。因此,大西洋的年轻深水的DIP 浓度通常约为1.5 μM;而那些较老的太平洋深水DIP 浓度则约为2.5 μM(Broecker et al., 1982)。相对于DIP,海洋中DOP 随深度分布的特征是表层海水中浓度较高,主要因为合成这些有机化合物的海洋生物大多生存在表层海洋中(Clark et al., 1999; Karl et al., 2001; Aminot et al., 2004)。这些DOP 大部分被细菌水解为DIP,随后被生物体迅速吸收和利用,只有一小部分被转移到深海。因此,随着海水深度的增加,DOP 的浓度通常较低。有趣的是,随着深度的变化,所有海洋盆地中DOP 的浓度波动较小,表明深海中大部分DOP 在海水中的停留时间相对较长(Kolowith et al., 2001)。海洋中的颗粒态磷主要由活的和死的浮游生物、POP(40%)、稳定的PIP(25%)、和不稳定的PIP(21%),以及少部分不参与任何反应的碎屑磷(13%) 组成(Faul et al., 2005)。下沉颗粒有机物的C/N/P 接近于Redfield 比值,说明其来源于海洋生物(Paytan et al., 2007)。Ruttenberg(1992)通过对浮游植物、下沉颗粒物、悬浮颗粒物以及沉积物中磷的连续提取(SEDEX)和核磁共振(NMR)分析,确定了这些磷储库的组成、相互关联和转化以及时空差异。不同磷储库在颗粒物中的总通量和相对分布存在显著的差异。浅层POP 较多(80%在透光区),而深层POP 的持续水解导致PIP 库的相对增加(POP <总颗粒态磷的25%)。而总磷、无机磷和有机磷在不同深度的通量随季节变化,且随深度的增加而显著降低。以上现象共同表明海洋磷循环在时空上的复杂性和高度的可变性。

图7 水体与沉积物中磷库之间的转化模式Fig. 7 Transformations between phosphorus pools in the water column and sediments (modified after Paytan et al., 2007)
海洋中的磷不断地在有机和无机态、颗粒和溶解态之间发生着转化。DIP 被浮游植物吸收并转化为有机磷化合物进入食物链(Cotner et al.,1992)。在此期间被吸收的磷主要通过生物的排泄物和死亡残体以DOP 和DIP 的形式回到海水中(Anderson et al., 1970; Cotner et al., 2002)。有机结合态的磷(POP 与DOP)必须先经过水解生成正磷酸盐才能够被生物体直接利用,此过程发生在整个水体中。细菌和浮游植物合成的酶(磷酸单酯酶、磷酸二酯酶、核酸酶、核苷酸酶、激酶)可以催化POP 与DOP 水解释放磷酸盐,然后将其吸收(Cotner et al., 2002; Azam et al., 1983)。因此,细菌和浮游植物是海水中POP 与DOP 向DIP 转化的主要驱动者(Paytan et al., 2007)。
3.3 地球系统科学视角下磷的生物地球化学循环模型
磷在地球系统中的循环可以大致分为陆地生态系统和海洋生态系统。陆地生态系统参与循环的储库主要有:磷矿(8300 Tg)、土壤(95000 Tg)和陆生生物(470 Tg)(Yuan et al., 2018);海洋生态系统参与循环的储库主要有:表层海水(3100 Tg)(Paytan et al., 2007)、海洋生物(100 Tg)(Yuan et al., 2018)和深层海水(Paytan et al., 2007;Filippelli, 2002; William, 2013)。陆地生态系统和海洋生态系统之间相对独立又密不可分。磷在陆地生态系统中的循环是一个地表岩石中的磷不断被地表径流搬运流失的过程。因此,陆地生态系统是全球磷生物地球化学循环的源。磷在海洋生态系统中的循环是一个海水中的磷不断通过类似生物泵的效应传递到海底沉积物中的过程。因此,海洋生态系统是全球磷生物地球化学循环的汇。人类活动前,磷在源和汇之间的转化主要受到气候环境、风化作用和构造作用的控制。人类的工业和农业活动作为一种新的地质营力极大的影响和改变了磷的生物地球化学循环过程,使陆地生态系统输入海洋生态系统的磷的通量提升了0.5~3倍(Ruttenberg et al., 2014)。这个过程不但加速了磷矿资源的消耗,而且这些额外输入海洋的磷会造成海洋初级生产力激增,产生大规模的水体富营养化现象,从而改变大气成分,进而产生气候效应。气候变化不但会对大陆风化的速率产生影响,还会对人类的工业和农业活动产生影响(图8)。这个过程是十分复杂的,很难去控制这个动态循环的过程。因此,需要找到调控当前磷循环的有效方法,以维持未来的粮食生产并保持河流、湖泊和海洋的健康(Liu et al., 2016)。
4 磷的生物地球化学循环的环境意义
4.1 磷的生物地球化学循环与全球变化
“雪球地球”冰期之后磷酸盐向海洋中的输入增加导致海洋初级生产力和有机碳埋藏的速率大幅增高,并导致海洋—大气系统的氧化还原条件的转变及海洋、大气中氧气含量的增高(Planavsky et al., 2010; Reinhard et al., 2017; Filippelli, 2002)。这意味着磷是控制海洋初级生产力最主要的营养限制性元素。然而这只是地球化学家的观点。生物学家认为:向缺乏营养的表层海水样本中添加硝酸盐通常会刺激浮游植物的生长,而添加磷酸盐则不会,因此是氮控制了海洋的初级生产力(Tyrrell,1999)。Tyrrell 等(1999)通过在不同营养条件下对两种浮游植物(Nitrogrn-fixers:可通过光合作用固氮的浮游植物,如蓝藻; Non-Nitrogrn-fixer: 不可固氮浮游植物)的培养,得出了氮和磷控制海洋初级生产力的模型。此模型指示,在地质历史时期的长时间尺度上,海洋初级生产力的决定因素是磷,而非氮。Reinhard 等(2017)将初级生产者的C/P、表层海水中磷的净清除率以及磷、碳、氮循环和氧气之间的耦合关系(P-C-N-O2),通过建立生物地球化学模型(CANOPS)联系起来。这些模型计算表明磷通常在极低或极高的大气氧分压(pO2)下控制海洋初级生产力和大气中氧气的水平。然而在地史时期,两次氧化事件之间有十余亿年时间氧气含量处于现今氧含量的0.1~0.4 倍之间,此时若要平衡P-C-N-O2之间的循环,N 的固定率必须显著提高。表层环境下固氮浮游植物通过消耗氧气固氮产生硝酸根(NO3-),海洋内部则由于反硝化作用发生氮的丢失。这一过程限制了大气中氧含量的增涨(Reinhard et al., 2017)。因此,在地史时期两次氧化事件之间长达近二十亿年的时间里,磷和氮的动态耦合过程控制并稳定地球系统表面氧气水平和生物圈生产力;而在目前大气的氧气水平下,磷是控制海洋初级生产力最主要的营养限制性元素(Reinhard et al., 2017)。
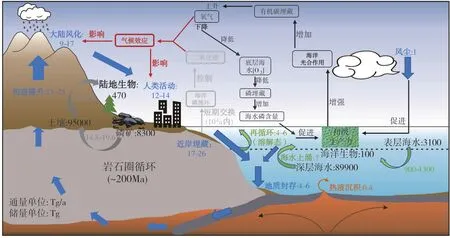
图8 全球尺度磷的生物地球化学循环概述及其与气候效应的耦合关系Fig. 8 A global scale schematic overview of biogeochemical cycles of phosphorus and its coupling with climate effects
全球尺度的气候变化与大气中氧气、二氧化碳的含量是紧密相关的(图8)。因此,海洋初级生产力的变化可以导致气候变化。反过来,全球气候变化又对海洋初级生产力具有重要影响。Holland(1994)认为,在大气圈氧气分压和氧气的产生速率之间,海洋磷酸根离子起了一个重要的连接作用。大量的磷酸根离子从河流或大气进入海洋后,可以与有机碳一起被埋藏于沉积物中,在这个过程中所产生的氧气量是非常巨大的。因此,通过海洋磷循环,由有机碳埋藏而产生的氧气可以与大气圈氧气水平相关联。显生宙以来,磷通过一种潜在负反馈机制(当大气氧含量降低时会导致底层海水氧浓度降低,从而降低海洋中磷的埋藏通量,使海水中磷浓度升高并促进海洋初级生产力,海洋初级生产力的增高会增强海洋中初级生产者的光合作用强度,进而增加有机碳埋藏,最终导致大气氧含量重新升高回到正常水平)在百万年尺度上控制着大气中氧气的平衡(Algeo et al., 2007)。Broecker(1982)则强调,二氧化碳的变化与滨海沉积物中碳的短期(一万年内)交换有关,后者在一定程度上是受磷的变化所致。以上研究表明:磷的生物地球化学循环控制着海洋碳循环,从而对大气圈氧气和二氧化碳水平具有重要的影响。例如:晚泥盆世陆地维管植物的爆发,使大陆生物化学风化加强,造成大量营养元素从陆地输入海洋,引起大规模赤潮进而导致海洋缺氧,且因海洋初级生产力激增而大幅增强的生物泵将大气中的二氧化碳更多的固定在海洋中,造成全球变冷。海洋缺氧与全球气候的变化,最终导致晚泥盆世大灭绝事件的发生(徐冉等,2006;陈代钊等,2006;沈树忠等,2017)。然而,晚泥盆世大灭绝之后生物圈的再次繁盛,很可能受控于磷对大气中氧气的负反馈调节机制。
海洋中一份磷可以固定106~170 份碳(Filippelli, 2008)。在当今二氧化碳排放过量导致全球变暖的背景下,过量排入海水的磷有助于固定更多的大气二氧化碳。但鉴于多种因素对海洋生态系统的复杂影响,很难预测未来由于人类活动引起的海洋中磷输入的倍增对全球气候的影响(Ruttenberg et al., 2014)。但这部分额外的磷的输入造成的富营养化现象对海洋生态系统的巨大破坏是必须引起警觉的。
4.2 磷的生物地球化学循环与可持续农业
磷元素对生命的重要性是无可代替的(Ashley et al., 2011)。然而自然体系下,由于磷灰石的溶解度非常低,以磷灰石的形式存在于土壤中的磷难以被生物利用,导致磷成为了地球表面大多数环境中动植物生产力的关键限制元素(Oelkers et al., 2008)。因此为了维持粮食的高产量,现代农业需要从磷矿中提取磷来满足作物生长需求。据估计,从磷矿中提取的磷约有90%用于生产农业化肥(Brunner, 2010)。而施用于农田的磷除约60%来自磷矿外,还有部分来自有机残留物,如粪肥和作物残渣(Liu et al., 2008)。
向农田中施用大量的磷会增加土壤肥力,少部分能够被作物利用增加其产量,但大部分除了以矿物吸附态(主要为磷与铁、铝氧化物和氢氧化物的吸附,参见Ruttenberg et al., 2014)或有机磷固定在土壤中外,均通过淋滤从土壤中损失(Manning, 2008; Kochiank, 2012)。这部分磷会和其它人类活动产生的磷(生活污水污水、畜禽养殖、纸浆制造)一起汇入河流、地下水和河口等淡水水体,引起水体富营养化(Powers et al., 2016)。进入海洋的磷会稳定固存在洋底沉积物中且在人类现有的技术下无法利用。因此,磷矿资源可能在50~100 年内就会枯竭(Cordell et al., 2009)。
已有研究提出以下缓解磷矿资源危机的措施:(1)培育贫营养耐受的植物品种(Kochiank,2012);(2)筛选和培育可高效从土壤中释放磷酸盐及可进行“元素分类”的微生物(Newman,2001; Hutchens et al., 2006; Oelkers et al., 2008;Stipp, 2008);(3)探索可持续循环利用磷的途径,例如通过改进农业技术提高粮食生产系统的磷效率及推广回收肥料、污泥和餐厨垃圾中的磷的技术(Yuan, 2018);(4)在全球范围内寻找更多可供开采的磷矿(Cooper et al., 2011)。
5 结语与展望
综上分析,磷的生物地球化学循环是一个具有反馈机制的过程,这个过程以气候为纽带,动态调节磷在各个圈层之间的通量。同时,气候的变化是多种因素相互耦合的复杂结果,涉及C、N、Fe 的生物地球化学循环;地球行星轨道的变化;海洋和陆地的分布等。因此,在研究磷的生物地球化学循环时,不但要关注各个圈层之间磷的通量和运移转化机制,也要意识到其它对气候产生影响的因素会间接影响到各圈层之间磷的运移。必须指出深层海水中的磷占海洋总磷的百分之九十(Paytan et al., 2007),而上涌的深层海水中生物可利用磷的量并不清楚,其对海洋初级生产力的贡献也很难估计。因此,未来有必要对上涌深层海水、人类活动以及大陆风化对海洋初级生产力的贡献进行定量研究。另外,海底热液作为深部地幔岩石圈与海洋水圈联通的“介质”,对磷在海洋中的沉积也有重要作用,这部分磷会随洋壳向板块俯冲边缘运移,也可能与深层海水交换并随上升流重新回到表层海水中被生物利用。此外,随热液进入海水的氟会极大的降低磷灰石的溶解度,这也解释了为什么CFA 是地层中最普遍的磷灰石类矿物(Ruttenberg et al., 2014)。因此,针对洋中脊及海底火山附近由于热液而发生沉积的磷的通量和运移过程也需要开展更为系统地研究。
近年来先进仪器分析技术的出现使我们能够直接确定磷的化学形态并研究其在环境中的变化过程,极大地提高了我们对各个储库中磷的性质及磷在储库间运移转化机制的认识,并且更加精确地估计了磷在地球各个储库中的储量及各个储库之间的转化通量。在过去的20 年中,一些研究利用一系列显微镜、光谱和波谱技术,在分子尺度上表征磷物种的形成。例如:Diaz 等(2008)应用基于同步辐射的X 射线光谱显微镜对同一位置的海洋沉积物进行了研究。她报道的光谱特征与聚磷酸盐、自生磷灰石以及两者之间的过渡相一致,表明多聚磷酸盐在成岩过程中转变为自生磷灰石。Brandes 等(2007)利用磷近边X 射线荧光光谱(P-NEXFS)绘制了不同的有机和无机磷化合物的参考结构模型,并以亚微米尺度的分辨率研究了加拿大埃芬汉入海口沉积物中磷的形态(Brandes et al., 2007)。Cade-Menun(2005)总结了土壤和水环境中观测到磷的固体与液体核磁共振(NMR)信号。一般来说,31P-NMR 信号带处于25×10-6~-25×10-6之间,包括正磷酸盐(orthophosphate, ortho-P)5×10-6~7×10-6、焦磷酸盐(pyrophosphate, pyro-P)-4×10-6~-5×10-6、多聚磷酸盐(polyphosphate,poly-P)-20×10-6、磷酸单酯(phosphate monoesters)3×10-6~6×10-6、磷酸二酯(orthophosphate diesters)2.5×10-6~-1×10-6以及膦酸酯(phosphonates)20×10-6(Cade-Menun, 2005)。目前全球工业开采的磷矿石,大约有85%来自沉积岩型磷矿,然而这些研究大都集中于探讨沉积型磷矿的成矿环境和形成条件,很少有研究从分子水平的微观尺度去探究沉积型磷矿最主要的矿物成分CFA的形成机制。因此,未来可以利用固体核磁共振、同步辐射等技术,探究CFA的形成机制,为沉积型磷矿的成因提供更微观的证据,这将是未来的热点研究方向。
此外,同位素技术在磷的生物地球化学循环研究中的应用(包括稳定同位素和放射性同位素)使我们对磷在不同储库之间循环的生物地球化学过程及不同形态之间的转化速率有了更加深入的了解(Helfenstein et al., 2018)。磷酸盐中的P-O键可以抵抗无机条件下的水解。在大多数自然系统的温度和pH 值下,如果没有生物的作用,磷酸盐不能与水交换氧(O’Neil, 2003)。因此,任何观察到的磷酸盐氧同位素组成的变化,要么反映了同位素组成不同的磷酸盐源的混合,要么反映了磷酸盐在活的细胞中循环过程中由于氧的交换而发生的磷酸盐氧同位素组成值的改变。以往的工作表明,细胞内焦磷酸酶的无机磷循环会导致与温度相关的同位素分馏,使在细胞内循环的磷酸盐中的氧同位素组成值达到平衡(Blake et al.,2005; Chang et al., 2015)。此外,Pucéat 等(2010)在实验室对温度控制生物成因磷灰石与水中氧同位素分馏的过程进行了研究。磷有两种放射性同位素32P(半衰期14.3 d)和33P(半衰期25.3 d)(Ruttenberg et al., 2014)。通过测量33P/32P 比值在雨水和不同的海洋磷汇中的比值,可以提供关于溶解有机磷和磷酸盐的周转速率的定量信息,以及磷在+5 和+3 价态之间的循环速率的定量信息(Van Mooy et al., 2015)。因此,近年来放射性磷同位素越来越多的用于研究海洋环境中磷的转化(Benitez-Nelson et al., 2002; Van Mooy et al., 2015;Sokoll et al., 2017)。
越来越多的研究通过这些先进的技术和方法揭示磷在陆地土壤和海洋沉积物中的赋存形态及迁移转化机制,从而找到引起磷短缺的资源危机问题和磷过剩的环境污染问题的关键生物地球化学循环过程。并通过这些原理来开发和利用城市磷矿资源(主要是餐厨垃圾、肥料、动物粪便及工业废弃物)、调控工农业活动减少溶解态磷向海水中的输入、提高土壤中磷的利用效率发展节约高效的可持续农业,最终实现全球磷资源的可持续利用与开发。
致谢:特别感谢中组部千人计划青年项目和南京大学登峰人才计划资助。
猜你喜欢
云南化工(2021年9期)2021-12-21
有色金属(矿山部分)(2021年4期)2021-08-30
矿产勘查(2020年5期)2020-12-19
矿产综合利用(2020年1期)2020-07-24
疯狂英语·新读写(2018年3期)2018-11-29
中国化肥信息(2018年5期)2018-08-04
电镀与环保(2016年2期)2017-01-20
环境科技(2016年1期)2016-11-08
三峡大学学报(自然科学版)(2016年6期)2016-04-16
无机化学学报(2014年12期)2014-02-28
