
宁夏马铃薯僵顶植原体的分子鉴定
2021-04-30 09:03李正男马强张磊孙平平赵文军周洪友
植物保护 2021年2期
李正男 马强 张磊 孙平平 赵文军 周洪友

关键词马铃薯僵顶植原体;16S rRNA基因;rp基因
马铃薯是我国主要粮食作物之一,是世界上仅次于水稻、玉米、小麦之后的第四大粮食作物,对我国粮食安全具有重要战略意义。我国马铃薯种植区主要划分为西北、西南、内蒙古和东北四大优势产区。宁夏属于西北马铃薯优势产区,2016年,我国马铃薯种植面积为554.52万h㎡,宁夏马铃薯种植面积为16.88万h㎡,马铃薯种植业已经发展成为当地农民脱贫致富的主导产业。随着马铃薯种植面积的扩大,病害的发生已经发展成为制约我国马铃薯高产、优产的主要因素,除传统的晚疫病Phytophthora in fstarts、黑胫病Pectobacte-rium spp.、环腐病Clavibacter michiganensis sub-sp.sepedonicus和病毒病外,由植原体引起的马铃薯僵顶病(potato stolbur)日益发展成为主要病害。
马铃薯僵顶病是一种植原体病害,发生普遍、传播迅速、危害严重。其症状主要表现为叶片上卷、红叶或紫叶、节点膨大、节间缩短、芽增殖、气生薯和早衰,部分发病轻的植株地下薯块虽然无肉眼可见症状,但质地蓬松(海绵薯),在加工过程中品相变差,如制作炸薯片时,会出现薯片褐变,丧失食用和经济价值。马铃薯僵顶病的流行主要有以下两方面原因,第一,由介体昆虫甜菜叶蝉Circuli fer tenel-lus传播,散布迅速;第二,通过薯块营养繁殖传播。据报道,该病害在北美、南美、欧洲、新西兰、俄罗斯和伊朗均有发生,并且引起严重的马铃薯减产。已经确认马铃薯僵顶病可由16SrⅠ、16SrⅡ、16SrⅢ、16SrⅥ、16Sr X、16SrⅫ、16SrXⅢ和16SrXⅧ组植原体引起。在我国云南和内蒙古个别马铃薯种植区内,马铃薯僵顶病的发生率达80%~100%,由16SrⅠ和16SrⅫ组植原体引起。
宁夏作为我国马铃薯的主产区之一,是否有马铃薯僵顶病发生,其病原的分类地位等尚不清楚。为此,我们开展了宁夏马铃薯僵顶病调查和病原鉴定,以期为马铃薯僵顶病的预测及病原菌研究奠定基础。
1材料與方法
1.1材料
2018年9月10日-17日自宁夏回族自治区固原市彭阳县红河镇采集表现叶片上卷、红叶、节问缩短、气生薯、根系坏死等症状的马铃薯样品,马铃薯品种为‘费乌瑞它。用于分子检测的样品采样方法如下:每株取5~8片新叶,以锡箔纸包裹后迅速液氮冷冻,通过干冰运回实验室,保存于超低温冰箱(-86℃)备用,其中有症状样品10个,无症状样品3个。用于透射电镜观察的样品采样方法如下:取10~15cm有症状植株茎尖,用打湿的报纸迅速包起来,保存在泡沫塑料盒中,带回实验室。在分类上属于16SrⅠ-B亚组的苦楝丛枝样品为本实验室保存。
Ex Taq DNA Polymerase.dNTPs(2.5 retool/L)、克隆载体PMDl9-T simple vector、DH5a感受态细胞均购自宝生物工程(大连)有限公司。琼脂糖凝胶回收试剂盒、高纯度质粒小量快速提取试剂盒购自艾德莱生物技术有限公司。1 kb DNA Ladder购自北京天根生化科技有限公司。
1.2透射电子显微镜观察
从显症马铃薯叶脉取2mm×2mm大小的韧皮部组织,将样品先用3%的戊二醛和4%的多聚甲醛混合物固定4h,然后用1%锇酸再次固定2h,10%~70%的乙醇和0~100%的丙酮逐级脱水,用812环氧树脂包埋,在烘箱内烘干72h后取出修块,修块完成后切半薄切片,定位到韧皮部组织后进行超薄切片,将超薄切片以醋酸铀酰及柠檬酸铅进行双重染色,用JEM-1200X透射电镜观察并拍照。
1.3总DNA提取
采用CTAB法提取冻存的13个马铃薯叶片样品总DNA,溶解于50μL灭菌双蒸水中,采用Eppendorf BioPhotometer D30核酸蛋白测定仪测定DNA浓度,通过琼脂糖凝胶电泳检测DNA的完整性后将总DNA保存于超低温冰箱(-86℃)中备用。苦楝丛枝样品DNA提取方法同上。
1.4马铃薯僵顶病的PCR检测
16S rRNA基因的PCR扩增:以提取的总DNA为模板,用植原体16S rRNA基因通用引物P1(5,-AAGAGTTATCCTGGCTCAGGATT-3)和P7进行扩增。扩增程序为:98℃预变性3min;98℃ 1 min,50℃2min,72℃ 3min,35个循环;72℃10min。将第一轮PCR产物稀释30倍作为模板,用引物R16F2n进行第二轮PCR,扩增程序为:98℃预变性3min;98℃1min,52℃1 min,72℃1min,先进行5个循环,然后以94℃30 s,50℃1min,72℃2min,进行25个循环;72℃延伸10min。
rp基因的PCR扩增:使用植原体rp基因通用引物rpF1C(5,-ATGGTDGGDCAYAARTTAGG-3)和rp(I)RIA(5-GTTCTTTTTGGCATTAA-CAT-3)进行扩增。扩增程序为:98℃预变性11min;98℃ 1min,50℃2min,72℃3min,38个循环;72℃延伸7 min。
取5μL PCR产物,用1%琼脂糖凝胶电泳检测,EB染色后在凝胶成像系统下成像。PCR反应以健康马铃薯总DNA为阴性对照,采集自陕西省杨凌区西北农林科技大学校园的苦楝丛枝样品DNA为阳性对照。
1.5PCR产物的纯化、克隆及序列测定
采用快捷型琼脂糖DNA回收试剂盒纯化PCR产物,纯化产物与PMDl9-T simple vector 16℃连接1h,将连接产物转化大肠杆菌DH5a感受态细胞,经过蓝白斑筛选后,挑取筛选平板上的白色菌落,在3 mL Amp抗性的LB液体培养基中培养过夜,经过PCR鉴定为阳性的菌液提取质粒,重组质粒送上海生工生物技术有限公司进行测序。
1.6序列一致性比较分析
采用Vector NTI Advancell(Invitrogen Corp.)软件对测序结果进行校正和拼接;将获得的16S rRNA和rp基因序列进行BLASTn比对检索同源序列并提交GenBank;通过DNAMAN软件将测定的16SrRNA基因和rp基因序列与同组植原体代表株系进行核酸一致性比较;通过MEGA 6.0软件分别构建16S rRNA基因和rp基因系统发育进化树。
2结果与分析
2.1马铃薯僵顶病症状
2018年9月10日-17日,对宁夏回族自治区固原市彭阳县红河镇马铃薯僵顶病进行了调查,调查标准主要是看马铃薯田里是否存在叶片上卷、红叶、节问缩短、气生薯和根系坏死症状的植株。结果发现,表现上述症状的马铃薯植株在薯田内零星发生,共采集了10株有症状的马铃薯叶片样品,3株无症状马铃薯叶片样品(图1)。
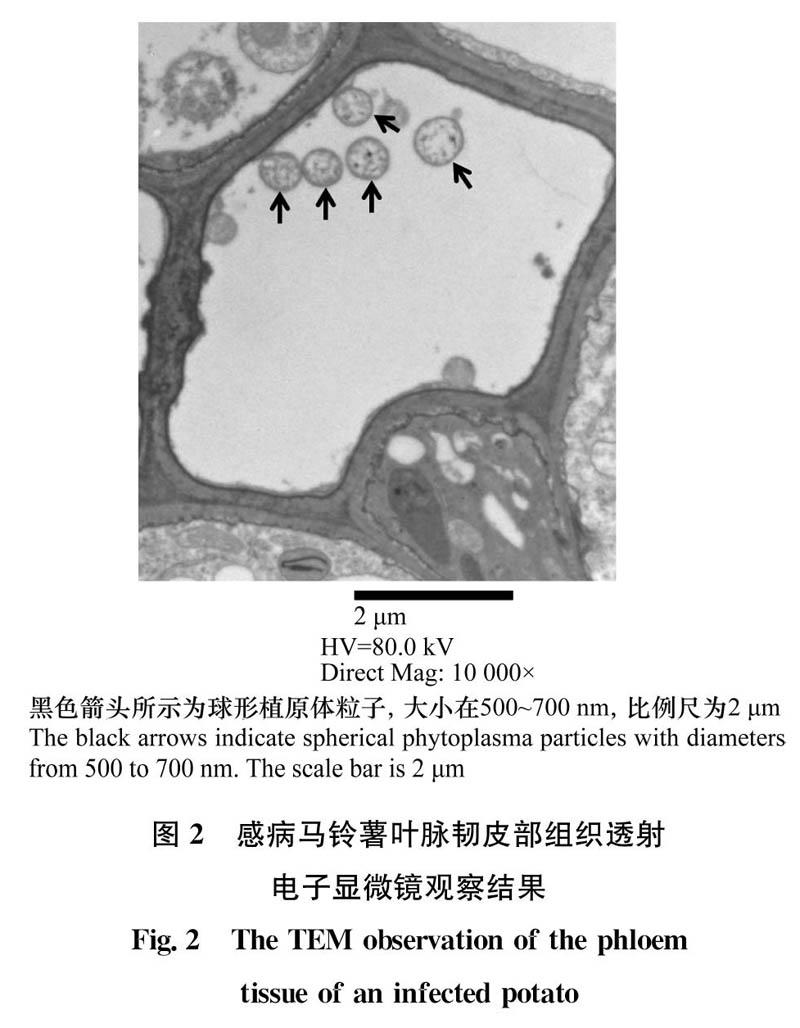
2.2电镜结果
通过透射电子显微镜在感病马铃薯韧皮部筛管细胞内观察到大量球形的植原体粒子,大小在500~700nm(图2)。
2.3 PCR鉴定结果
利用植原体16S rRNA基因和rp基因通用引物从有症状的10个马铃薯样品和阳性对照中均扩增出长度约1.2 kb的16S rRNA基因和rp基因,而3个无症状马铃薯样品中未扩增出特异性条带。证明了表现叶片内卷、红叶、节问短缩、气生薯和根系坏死症状的马铃薯中有植原体存在。
2.4一致性分析和系统发育分析
从10个PCR鉴定为植原体阳性的马铃薯样品中随机选择2、4、7、9号样品进行16S rRNA基因和rp基因克隆,16S rRNA基因和rp基因均选择3个阳性克隆进行测序,4个样品彼此问16S rRNA基因和rp基因的一致性为100.0 %,我们将该植原体命名为马铃薯僵顶植原体宁夏株系(potato stolburphytoplasma strain Ningxia,PSp-Nx),16S rRNA片段长度为1246 bp,G+C含量为46.79%,登录号为MK696087;rp基因长度为1182 bp,登录号为MK695939。通过BLASTn程序,将获得的马铃薯僵頂植原体宁夏株系16S rRNA基因序列和rp基因序列在GenBank中进行同源序列搜索,分别搜索到99条同源序列,99条序列均属于僵顶组(16SrⅫ组)成员,其16Sr RNA基因与‘Ca.P.ffagariae槭树株系(MK501642)16S rRNA基因一致性最高,为99.70%,rp基因与‘P.fragariae云南马铃薯YN-2G株系(KJl44889)rp基因一致性最高,达到100.0%。
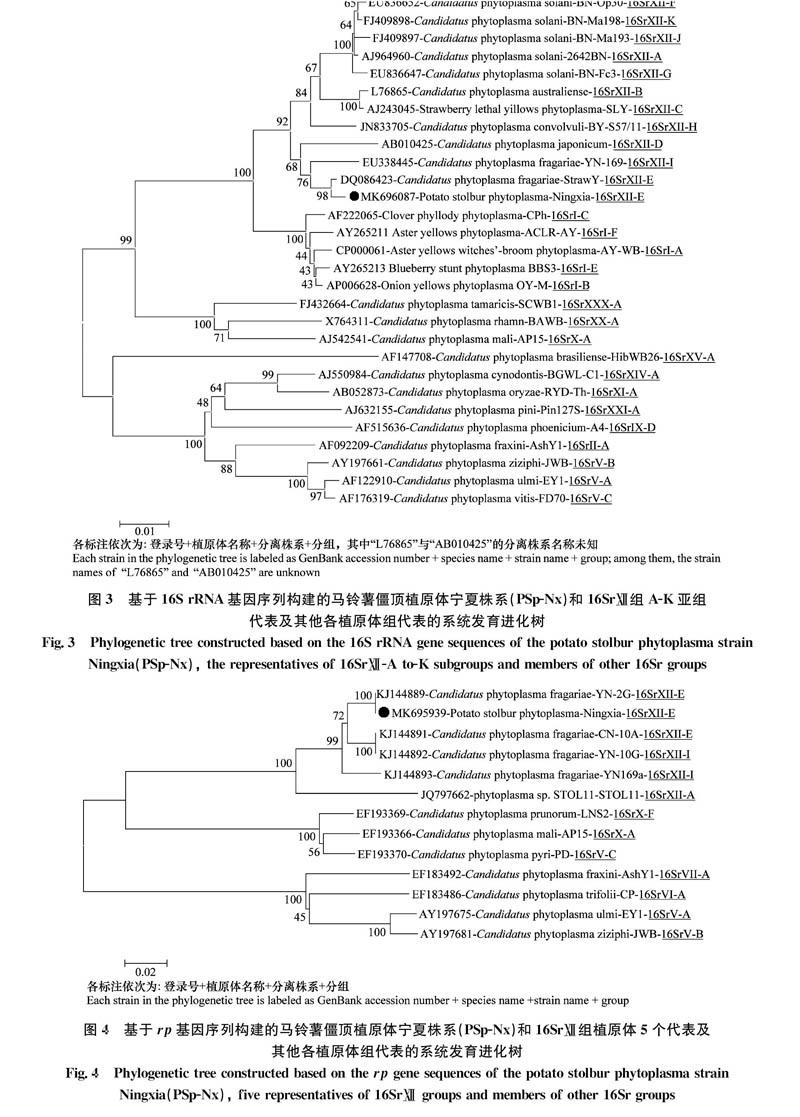
利用MEGA 6.0软件将马铃薯僵顶植原体宁夏株系16S rRNA基因序列与僵顶组植原体11个亚组(A-K)代表及其他各植原体组代表的16SrRNA基因序列进行比较,构建系统发育进化树(图3)。从图中可以看出马铃薯僵顶植原体宁夏株系与僵顶组植原体11个亚组代表株系聚集在一起形成一个大的分支,其中又与16SrⅫ-E亚组的代表株系‘ca.P.fragariae草莓株系(DQ086423)聚集在一起形成独立分支。马铃薯僵顶植原体宁夏株系rp基因序列与僵顶组植原体5个代表及其他各植原体组代表的rp基因序列进行比较,构建系统发育进化树(图4)。从图中可以看出马铃薯僵顶植原体宁夏株系与僵顶组植原体5个代表株系聚集在一起形成一个大的分支,其中又与分类属于16SrⅫ-E亚组的‘ca.P.fragariae云南马铃薯YN-2G株系(KJl44889)聚集在一起。
基于对16S rRNA基因和rp基因的核酸一致性分析和系统发育分析,均证明马铃薯僵顶植原体宁夏株系属于16SrⅫ组,并且该分离物与16SrⅫ-E亚组亲缘关系最近。
3讨论
在本研究中透射电镜和分子检测结果均证明在宁夏发生的马铃薯僵顶病与植原体有关;其16SrRNA基因和rp基因的一致性分析和系统发育分析证明该植原体属于16SrⅫ组,并且与16SrⅫ-E亚组亲缘关系最近。
之前的研究表明在我国北方地区16SrⅠ组和16SrV组植原体发生最为普遍,南方地区16SrⅠ组和16SrⅡ组植原体发生最为普遍_2,但近年来越来越多的研究报道僵顶组(16SrⅫ组)植原体在我国的发生,其中Gao等报道了16SrⅫ组一个新的亚组在山东牡丹上的发生,Yang等报道了16SrⅫ-A亚组在陕西丹参上的发生,Wu等报道了16SrⅫ-c亚组在云南黄槐树上的发生,Cheng等报道了16SrⅫ-E、16SrⅫ-I亚组在我国云南和内蒙古马铃薯上的发生,由此,我们可以看到16Sr组植原体在我国不同地区正在广泛发生。由16Sr组植原体引起的病害已经发展成为北欧最严重的植原体病害,因此16Sr组植原体在我国的发生值得引起重视。
在我国西北马铃薯产区的陕西、西南马铃薯产区的云南、内蒙古马铃薯产区分别有16SrⅠ-B、16SrⅥ-A、16SrⅫ-E、16SrⅫ-I亚组植原体侵染马铃薯引起僵顶病的报道,但在宁夏和黑龙江等马铃薯产区还没有马铃薯僵顶病发生的报道,本研究对宁夏马铃薯产区进行了调查,并证明在宁夏有16SrⅫ-E亚组植原体引起的马铃薯僵顶病。结合上面研究结果证明与植原体相关的马铃薯僵顶病在我国马铃薯产区普遍发生。虽然在本研究中鉴定到的马铃薯僵顶植原体宁夏株系与在云南和内蒙古鉴定到的马铃薯僵顶植原体均属于16SrⅫ-E亚组,但马铃薯僵顶植原体宁夏株系16S rRNA基因却与在比利时发生的‘ca.P.fragariae槭树株系(MK501642)一致性最高,为99.70%,说明马铃薯僵顶植原体宁夏株系与云南和内蒙古株系存在一定变异。在世界范围内已经有16SrI、16SrII、16SrIII、16SrVI、16SrX、16SrⅫ、16SrⅫ在马铃薯上引起僵顶病的报道,结合我国的研究结果可以看出引起马铃薯僵顶病的植原体病原多样,植原体由刺吸式口器昆虫传播,这类昆虫食性广泛,它们可能会在感染了不同组植原体或者同组不同亚组植原体的植株上取食,这可能是导致马铃薯僵顶病植原体病原多样性的原因,在油菜、葡萄、长春花等植物上也存在分类上属于不同组的多个植原体组植原体侵染的报道。我国不同马铃薯产区僵顶病的发生情况、病原多样性和效应因子鉴定将是我们后续工作的重点。
猜你喜欢
少儿科学周刊·儿童版(2021年21期)2021-12-11
中国食品(2020年9期)2020-05-26
宁夏画报(2019年5期)2019-09-19
中小企业管理与科技·下旬刊(2019年3期)2019-07-08
人生与伴侣·共同关注(2018年4期)2018-04-28
宁夏画报(2017年12期)2018-01-30
食品安全导刊(2017年12期)2018-01-04
食品界(2017年7期)2017-08-24
儿童故事画报·发现号趣味百科(2016年7期)2017-02-08
宁夏画报(2016年5期)2016-06-28
