
茅苍术试管苗继代培养的ISSR及MSAP分析
2021-04-28 03:52张成才向增旭
中国农学通报 2021年12期
张成才,向增旭
(南京农业大学园艺学院,南京210095)
0 引言
菊科苍术属多年生植物——茅苍术Atractylodes lancea(Thunb.)DC为江苏省茅山地区道地药材,具有燥湿健脾,祛风散寒,明目等功效[1],其质量和疗效也备受历代医家推崇,近年来由于生境变化、人为采挖等导致茅苍术野生资源已濒临灭绝,种苗来源不足是制约人工栽培进展的主要原因之一[2]。为了保护这一种质资源、满足市场需求,将组织培养应用于实际生产中,能在短期内获得大量优质试管苗,满足生产种苗需求,目前已应用于工厂化生产中[3]。但研究发现,组培材料在长期继代培养过程中存在着广泛的遗传变异,而大量变异会影响种苗的品质。因此,探究茅苍术试管苗在长期继代培养中的遗传稳定性具有非常重要的意义。
近年来,生物技术的发展使得药用植物研究更加多元化,利用分子标记技术对组培材料进行遗传稳定性的检测,可准确检测出变异发生的代数,有利于优良种质资源的保存和利用,目前,已成功应用于多种药用植物中[4-5]。其中,简单序列间重复(inter-simple sequence repeat,ISSR)作为目前广泛应用的DNA分子标记技术,具有操作技术简单,灵敏度高等优点。DNA甲基化水平的变化受生理状态、发育阶段及环境刺激多种因素的影响,是一种重要的表观遗传修饰形式[6]。组培材料在经过多次继代培养后甲基化水平往往会发生变化,甲基化敏感扩增多态性(methylation sensitive amplification polymorphism,MSAP)技术常用于DNA甲基化水平和模式的分析,具有简单易行、通用性强、可有效监测DNA甲基化与基因表达关系等优点[7-8]。许梦云等[9]运用ISSR分子标记技术对茅苍术和北苍术进行多态性分析,发现茅苍术比北苍术的遗传多样性更高,与北苍术有很近的亲缘关系,且茅苍术具有明显的遗传分化。王红娟[10]运用ISSR分子标记技术研究二倍体和同源四倍体茅苍术的遗传多样性,发现茅苍术二倍体和四倍体的遗传距离和遗传相似系数差异较大,运用MSAP技术研究其基因组DNA甲基化水平及模式变化,发现四倍体基因组DNA全甲基化率低于二倍体,半甲基化率高于二倍体,与二倍体相比,四倍体DNA甲基化模式发生调整的基因位点占总检测位点的52.53%。本研究以茅苍术试管苗为材料,分别采用ISSR、MSAP两种分子标记技术进行分析,探究不同继代次数对茅苍术试管苗基因组遗传稳定性及DNA甲基化的影响,以期为茅苍术种质资源保存和工厂化生产提供理论依据。
1 材料与方法
1.1 材料
茅苍术野生植株2018年7月采挖于江苏省茅山地区,经南京农业大学园艺学院向增旭副教授鉴定为茅苍术Atractylodes lancea(Thunb.)DC。取茅苍术健壮顶芽建立无菌系转入相同的继代培养基(MS+1.0 mg/L 6-BA+0.1 mg/LNAA)进行继代培养,以25天为培养周期,共继代10代,样本从第1代到第10代分别编号为J、G、2、3、4、5、6、7、8、9。
限制性内切酶、T4 DNA连接酶购自上海泽叶生物科技有限公司。Taq酶和d NTPs购自生工生物工程(上海)股份有限公司。
1.2 茅苍术基因组DNA提取、引物筛选、ISSR-PCR反应体系优化和扩增产物检测
采取改良的CTAB法进行DNA提取,提取产物取2~3 μL,用2.0%琼脂糖凝胶电泳检测,其余置于-20℃保存备用。
ISSR-PCR反应扩增体系(25 μL)为:DNA模板约40 ng,10×Bufer2.5 μL,primer 0.3 μmol/L,4种dNTPs各0.25 μmol/L,MgCl21.5 mmol/L,TaqDNA聚合酶(北京优吉科技有限公司)1.25 U/μL,引物0.6 μmol/L,缓冲液2 μL,dd H2O补足至20 μL。
PCR扩增程序为:94℃预变性5 min,然后进行35个循环:94℃变性 45 s,55~62℃复性 45 s,72℃延伸1.5 min;循环结束后72℃延伸10 min,4℃保存。
PCR反应结束后,将PCR产物在2.0%的琼脂糖凝胶电泳中检测。无DNA条带记为0,有条带记为1,形成ISSR的表型数据矩阵NTSYS软件计算相似系数(DICE系数),并且按照遗传系数进行UPGMA聚类分析。
选取J、3、6、9,4个样品进行引物筛选,从56条随机引物选出稳定扩增并有多态性的10条引物(表1)进行分析试验。所有引物由上海杰瑞生物科技有限公司合成。所有扩增产物在2%琼脂糖凝胶中电泳1 h,凝胶成像仪下拍照并观察。

表1 ISSR分析试验所用引物序列
1.3 MSAP分析
用识别四碱基的HpaII和MspI同裂酶分别与识别六碱基的核酸内切酶EcoR I组合对样本DNA进行双酶切,DNA酶切反应体系为20 μL。在酶切片段的两端加上人工设计的与EcoR I和Hpa II/Msp I酶切位点互补的人工接头(表2)。接头连接体系为20 μL:2 μL 10×T4 Buffer、0.4 μL 20 μmol/L EcoR I和Hpa II/Msp I接头,其余用水补齐,16℃过夜。

表2 MSAP分析试验所用接头和引物信息
预扩增与选择扩增反应参考韩盼盼等[11]的方法,并做适当调整。3730XL测序仪对PCR产物进行毛细管电泳后利用GeneMarker 2.2软件将10个样品、14对引物组合扩增产物测序仪得到的原始数据进行分析,根据无带和有带情况转化为0,1数据矩阵,然后进行甲基化分析。
2 结果与分析
2.1 茅苍术ISSR分子标记遗传多样性分析
2.1.1 ISSR引物检测的位点数和多态性 根据PCR扩增结果,以引物ISS6的扩增电泳图为基础,用Quantity one软件绘制电泳模式图(图1)。10条引物在10个样品中共扩增出了80条清晰条带,其中多态位点47个,多态百分数为25%~87.50%,平均多态百分数为56.58%。各引物扩增的条带在6~12条之间,平均每条引物扩增的条带数为8条;从单个引物扩增得到的多态位点来看,引物0531-018扩增得到的多态位点最多,为12个,10条引物扩增得到的多态位点平均为4.7个。引物P10(873)和ISSR4的扩增图谱如图2所示,茅苍术试管苗从继代第四代开始,条带在个别位点有减弱或增强的变化,并且随着继代次数的递增,变化愈明显。在引物P10(873)扩增图谱中,第4、7、10代条带在200 bp左右位点发生缺失,但总体呈逐渐减弱趋势。
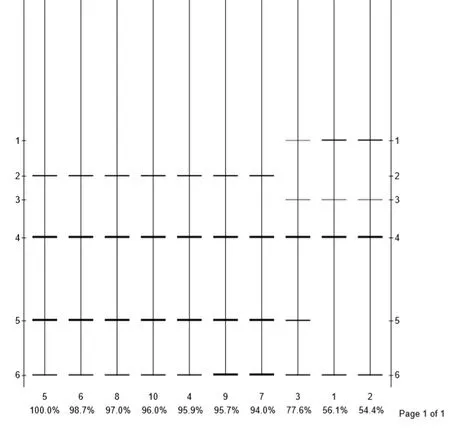
图1 电泳模式图(引物ISSR6部分)
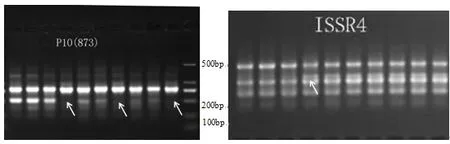
图2 引物P10(873)和ISSR4的扩增图谱
利用Popgene.32软件对10个样品的10条引物分别进行遗传多样性分析,结果如表3所示。观察等位基因数Na范围为1.2500~1.8750之间,有效等位基因数Ne在1.2455~1.5437的范围之内,Nei’s基因多样性的范围在0.1239~0.3177之间,Shannon信息指数I为0.1721~0.4718,多态百分数在25.00%~87.50%之间,其中ISSR35的多态百分数最高为87.50%,而ISSR4的多态性百分数最低为25.00%,由以上数据可得,10个样本之间遗传多样性较低。

表3 10个样本遗传多样性分析结果
2.1.2 基于ISSR标记的茅苍术遗传多样性分析 根据“0”,“1”矩阵数据,利用Popgene32软件分别计算10个样本遗传相似系数矩阵,结果如表4所示。遗传相似系数为0.4750~0.9875,其中相似系数最大的为0.9875(样品4号与5号),亲缘关系最近,最小为0.4750(样品6号与G号),亲缘关系最远。根据遗传相似系数矩阵,对10份供试材料的ISSR数据的结果进行聚类分析,得到树状聚类图(图3)。样本间遗传相似系数平均值为0.7392,在遗传距离为0.74处,将10个样本划分为两个大类,第一类包括样本J、G、2号,即为继代培养的1~3代,其余样本均为第二类。综合以上数据可知,在继代培养的初期,样本遗传稳定性较高,随着继代次数的增加,遗传稳定性有所下降。

图3 10个样品基于遗传相似系数的UPGMA聚类分析

表4 基于ISSR标记的10个样品的遗传相似系数矩阵
2.2 茅苍术基因组DNA甲基化的MSAP分析
2.2.1 基因组DNA甲基化水平分析 依据HpaⅡ和MspⅠ对甲基化不同的敏感程度,MSAP片段可分为4种类型[16-17]。本试验选取14对引物组合(E34/MSP49,E34MSP60, E38MSP44, E38MSP50, E38MSP61,E41MSP40, E41MSP50, E44MSP41, E46MSP39,E50MSP39, E50MSP41, E50MSP59, E77MSP41,E77MSP50)对双酶切后的10个样本进行扩增。以继代次数第1代和第10代茅苍术试管苗(样品J与样品9)为例,如表5所示,样品J的总甲基化率79.09%,全甲基化率为70.17%,半甲基化率为8.92%,而样品9总甲基化率78.83%,全甲基化率为65.27%,半甲基化率为13.56%。经过长期继代培养后,总甲基化率降低了0.26%,全甲基化率降低4.9%,但半甲基化率提高了4.64%,表明甲基化水平虽然有所下降,但仍处于较高水平。

表5 样品J与样品9基因组DNA甲基化水平分析
2.2.2 样品J与样品9基因组DNA甲基化模式分析 茅苍术试管苗经过长期继代培养后,存在3种甲基化模式(表6),其中,甲基化条带数无变化所占比例为67.3%,去甲基化模式占18.87%,而甲基化模式比例为13.6%。与第1代试管苗(样品J)相比,继代培养第10代试管苗(样品9)的去甲基化比例为18.87%,甲基化模式的比例为13.46%,即在继代培养过程中共有32.7%的DNA基因组甲基化模式发生了改变。以上数据表明,在茅苍术试管苗经过长期继代培养过程后,去甲基化模式和甲基化模式并存,但以去甲基化模式为主,说明长期继代培养后有较多的基因被重新活化和表达。

表6 样品J与样品9基因组DNA甲基化模式分析
3 讨论和结论
植株离体培养材料在长期的继代培养过程中,往往会发生遗传变异和生理变化,刘福平研究发现,经组织培养产生的变异频率高于自然突变和常规繁殖方法。在一个组培周期内可产生1%~3%的无性系变异[12]。而不同植株、初始继代材料、培养条件等都会影响组培材料在继代培养中的遗传稳定性[13]。已有研究显示,火龙果继代到第4代时发生变异[14],而番木瓜却能够继代培养32代无明显变异[15],由此可见不同组培材料的遗传稳定性具有明显的个体差异。另外一些组培材料经长期继代培养后表观遗传修饰发生变化,但核苷酸序列并未改变。因此,在研究遗传稳定性时结合甲基化现象,能够更为全面的分析组培材料在继代过程中产生的遗传变异。
茅苍术ISSR分子标记遗传多样性分析结果显示,10个样本间遗传多样性较低,遗传相似系数最低的为第1代(样品J)与第10代(样品9),亲缘关系较远。同时通过聚类分析,可将10个样本分为两大类,第1~3代为第一大类,其余样本为第二类。在整个继代过程中,茅苍术试管苗主要遗传位点皆相同,但从继代第4代开始,在个别位点处谱带发生增强或减弱的现象;在引物P10(873)的扩增图谱中,第4、7、10代条带在200 bp左右发生位点缺失,从整体来看,随着继代次数的增加,变化愈明显。以上数据表明茅苍术试管苗在继代培养过程中虽然整体遗传稳定性较高,但从第4代开始仍发生了少量变异,变异程度随继代次数增加而递增,与火龙果的产生变异的继代次数相同,不同物种继代的遗传稳定性有异同。通过MSAP分析长期继代培养对对茅苍术试管苗甲基化水平和甲基化模式的影响,发现经过长期继代培养后,试管苗总甲基化率降低了0.26%,全甲基化率降低4.9%,但半甲基化率提高了4.64%,表明甲基化水平虽然有所下降,但仍处于较高水平,这与Li等[16]研究结果一致。与第1代试管苗(样品J)相比,继代培养第10代试管苗(样品9)的去甲基化比例为18.87%,甲基化模式的比例为13.46%。甲基化状态通常与基因表达失活有关,反之亦然[17]。茅苍术试管苗在继代培养后去甲基化模式和甲基化模式并存,但以去甲基化模式为主,DNA甲基化水平发生了一定程度的变异。以上数据表明,在长期继代培养后共有18.87%的基因被重新活化和表达,13.46%的基因被抑制表达。
茅苍术试管苗继代培养过程中所用培养基为MS+1.0mg/L6-BA+0.1mg/LNAA+30g/L蔗糖+7.5g/L琼脂(pH 5.8~5.9),继代出现变异现象可能与培养条件(培养基成分如激素、蔗糖含量等)以及培养时间的长短不同等因素有关,这些因素影响继代试管苗的遗传稳定性和DNA甲基化水平及模式,去甲基化模式高于甲基化模式可能也是植物适应性反应的迹象,从而导致更多的基因得到表达,今后将通过调控这些因素,降低茅苍术试管苗继代变异方面进一步研究。
本研究以茅苍术幼嫩顶芽为初始材料,建立茅苍术高效离体快繁体系并进行长期继代培养,运用ISSR、MSAP分子标记技术对不同继代次数的试管苗进行分析,茅苍术继代10代遗传稳定性较高,从继代第4代开始出现少量变异,第1代与第10代样本之间遗传相似系数最低,亲缘关系较远。继代长期培养后茅苍术试管苗的甲基化水平有所下降,去甲基化模式和甲基化模式并存,但以去甲基化模式为主。综上所述,本研究结果可为茅苍术种质资源保存和工厂化生产提供一定的理论依据。
猜你喜欢
河北农机(2020年10期)2020-12-14
今日农业(2020年13期)2020-08-24
中成药(2018年5期)2018-06-06
辽宁林业科技(2017年4期)2017-06-22
好孩子画报(2016年7期)2016-12-12
现代检验医学杂志(2015年2期)2015-02-06
沈阳医学院学报(2014年4期)2014-12-27
中小学实验与装备(2014年2期)2014-09-21
遗传(2014年3期)2014-02-28
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
