
珠三角河网链状硅藻物种组成及生态特征
2021-04-25 06:27葛大艳刘乾甫赖子尼贾慧娟杜宛璘
生态学报 2021年6期
葛大艳,刘乾甫,赖子尼,贾慧娟, 3,杜宛璘, 4,王 超,*
1 上海海洋大学海洋生态与环境学院, 上海 201306 2 中国水产科学研究院珠江水产研究所, 广州 510380 3 上海海洋大学水产与生命学院, 上海 201306 4 天津农学院,天津市水产生态及养殖重点实验室, 天津 300384
链状硅藻是指相邻细胞间通过刺状或突状结构相连形成丝状或链状群体的硅藻物种,例如海洋硅藻中的骨条藻(Skeletonemasp.)、角毛藻(Chaetocerossp.)、根管藻(Rhizosoleniasp.)等,是导致赤潮爆发的常见种类[1- 3];直链藻(Melosirasp.)和沟链藻(Aulacoseirasp.)则是淡水水体中常见的链状硅藻[4],在世界范围内广泛分布,它们通常在富营养化的河流和湖泊中大量繁殖,甚至爆发水华,同时也是贫中营养水体中浮游植物群落的重要组成部分[5]。
20世纪80年代前,淡水水体中常见的链状硅藻物种均被归属于直链藻属(MelosiraAgardh),直到1979年Simonsen分离了颗粒直链藻(Melosiragranulata)及相关物种如颗粒直链藻极狭变种(Melosiragranulatavar.angustissima)、模糊直链藻(Melosiraambigua)等,并在分类学上将它们转移到海藻科(Thalassiosiraceae)的沟链藻属(AulacoseiraThwaites)中,导致直链藻属仅剩下少数物种[6-7]。基于文献计量学的分析结果显示,目前国内外对链状硅藻的研究主要集中在对某一属(通常为沟链藻属)或某一特定物种的分类、多样性和种群生态学等方面的基础性研究上[5, 8- 12]。虽然在不同淡水水体中,链状硅藻中的某些物种常作为浮游植物群落的优势种出现[13- 17],但是我国对淡水链状硅藻开展的针对性研究并不多见。研究对珠江中下游水域中的浮游植物群落的优势种——颗粒沟链藻(Aulacoseiragranulata(Ehrenberg) Simonsen)开展了分类学、种群生态学以及形态性状指示功能等方面的研究[18- 22]。然而,有关珠江下游水域链状硅藻的多样性组成及生态竞争关系等研究仍未见有报道。
珠三角河网水域是珠江汇入南海的重要通道,该水域河道错综复杂,气候温润多雨,物种多样性丰富。该水域的水环境质量状况一直备受关注。根据水利部珠江水利委员会发布的《珠江片段水资源公报2011》显示,珠三角近1/4 河段属劣Ⅴ类水质。旨在掌握珠三角河网水域的链状硅藻物种多样性组成特征,分析物种间的竞争关系,挖掘不同链状硅藻物种对水环境的指示功能。研究结果将为该水域的生物多样性、环境指示生物等方面的研究,以及水环境保护提供科学依据。
1 材料与方法
1.1 调查站位与调查时间
在珠三角河网水域设置了13 个采样站位,基本呈伞状分布(图1)。其中青岐、左滩、外海和新围分布在西江沿线,小榄、小塘、北滘、榄核、横沥、陈村和市桥分布在河网中部,莲花山和珠江桥分布在广州市周边。调查时站位的定位采用GPS 全球卫星定位系统。调查时间分别为2012 年的3 月、5 月、8 月和12 月,其中3 月和12 月代表枯水期,5 月和8 月代表丰水期;以及2015 年的3 月、6 月、9 月和12 月,其中3 月和12 月代表枯水期,6月和9 月代表丰水期。每个时间段的采样在2—3 d之内完成。

图1 调查站位布设图Fig.1 Survey site layout
1.2 样本采集、处理及数据收集
浮游植物样本取表层(离水面0.5 m)水样1000 mL 装入聚乙烯瓶中,立即加入鲁哥试液固定,使其最终浓度为15‰。水样运回实验室,振荡摇匀后立即移入标记刻度为1000 mL 的玻璃量筒内,加盖静置24 h 后,用管口包裹筛绢的虹吸管或吸管小心吸去不含浮游植物的上清液,如此反复多次,直至剩下80—85 mL沉淀物转入100 mL定量瓶中,再取少许上述虹吸出来的上清液清洗3次沉淀器底部,清洗液仍须转入定量瓶中直至浓缩样品最终体积为100 mL,每瓶样品须标记采样的时间、地点、采样体积等信息后进行保存。
分析时先将浓缩后的样品以左右平移的方式摇匀,再取1 mL 样品注入Sedgewick-Rafte 浮游植物计数框中,在Nikon TS100 倒置显微镜下进行浮游植物的种类鉴定和计数,种类鉴定参考国内相关志书和专业资料[23],每瓶样品计数2次取平均值,每片计数100个视野。本文主要采用了链状硅藻的数据资料,其藻链的长度用每根藻链的细胞数来代表。
使用便携式水质分析仪(YSI6600-02, USA)测定水温、盐度和pH等现场环境因子,用透明度盘测定现场的透明度,另取500 mL 水样现场过滤后置入车载冰箱中运回实验室,用水质流动注射分析仪(SKALAR,荷兰)对硅酸盐、总氮和总磷等营养盐进行分析。
1.3 藻类数据处理与分析
浮游植物生物量的计算方法参照Hillebrand 等[24],通过体积法计算取几何近似值。一升水中的浮游植物的数量(N)可以用下列公式计算:
式中:Cs为计数框面积;Fs为每个视野的面积;Fn为每片计数过的视野数;V为水样沉淀浓缩后的体积;v为计数框的体积;Pn为每片实际数出的浮游植物个数。
各浮游植物的数量用细胞数表示,由于藻类细胞形态比较规则,细胞比重多数接近于1,可用形态相近似的几何体积公式计算细胞体积,细胞体积的毫升数相当于细胞重量的克数,若同时计算各种浮游植物细胞的体积,就可在数量的基础上进而计算出每升水中各种浮游植物的生物量。
时空分布特征用软件Origin 8.5完成;链状硅藻生物量与环境因子的关联性分析采用链状硅藻物种生物量相对百分组成数据,然后与环境因子数据取LOG值后,进行RDA分析,获得二维降序图。链状硅藻链长与环境因子的关联性分析未对数据进行标准化处理,进行RDA分析,获得二维降序图。
2 结果
2.1 环境理化因子
调查站位的理化环境特征见图2。其中丰水期的水温高于枯水期;透明度呈现从西江沿线至河网中部站位依次降低的趋势,枯水期的透明度高于丰水期;电导率和总溶解固体除了在2012年枯水期的莲花山站位出现极大值以外,二者在2012年的值普遍低于2015年;溶解氧在丰水期低于枯水期,在广州市周边站位要低于其余站位;硅酸盐的含量呈现从西江沿线、河网中部至广州市周边站位依次降低的趋势。
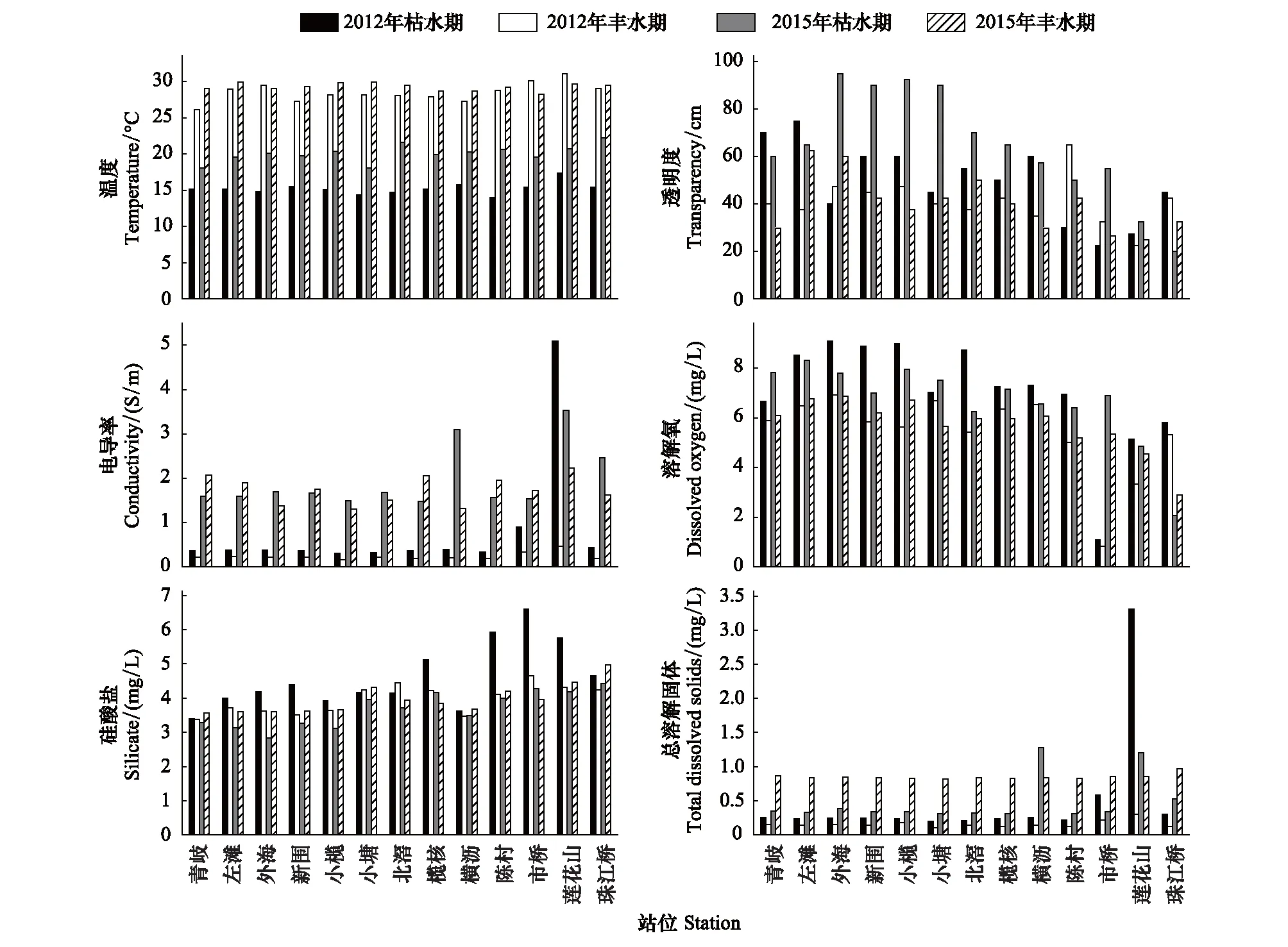
图2 环境理化因子的时空变化特征Fig.2 Characteristics of spatiotemporal changes of environmental physical and chemical factors
2.2 链状硅藻的物种组成
本研究共采集到链状硅藻18种(含变型),包括沟链藻属(Aulacoseira)11种和直链藻属(Melosira)7种。物种组成及相关信息详见表1。总体来看,链状硅藻的出现率为100%,对浮游植物群落生物量的贡献超过60%。沟链藻属中出现率最高的是颗粒沟链藻,两年的出现率几乎为100%,对浮游植物生物量贡献均超过50%;其次为颗粒沟链藻极狭变种,两年的出现率均为70%左右,生物量贡献均大于1%。直链藻属中出现率最高的是变异直链藻,2012年的出现率接近60%,2015年超过90%,生物量贡献均为1.7%左右;其次为芬兰直链藻,两年的出现率均大于50%,2015年的生物量贡献比2012年下降了1%。其他物种的出现率往往低于50%,且生物量贡献很小。
2.3 链状硅藻生物量与浮游植物总生物量的关系
对链状硅藻生物量与浮游植物总生物量进行线性回归分析,发现两者存在极显著意义(P<0.001,r2=0.85)的正相关关系(图3)。
2.4 生物量的时间变化特征
从表1列出的18个链状硅藻中选取两周年均有出现的9种作进一步分析(图4)。总生物量的时间变化特征显示,丰水期差异不大枯水期差异明显,且最大值和最小值均出现在枯水期。相对百分组成的时间特征显示,颗粒沟链藻是绝对优势种群,其在9种链状硅藻的生物量总和中所占百分比皆高于70%。颗粒沟链藻极狭变种一般在丰水期的贡献率要高于枯水期。模糊沟链藻日本变型在2012年的贡献率(17.05%)高于2015年(2.08%)。意大利沟链藻仅在个别时间段形成较大优势,其在2012年5月的贡献率为20.60%。远距沟链藻一般在枯水期的贡献率高于丰水期。变异直链藻一般在枯水期形成明显优势,2012年最高贡献率出现在3月为9.49%,2015年最高贡献率出现在12月为8.02%。

表1 链状硅藻物种组成、出现率及生物量贡献

图3 链状硅藻生物量与浮游植物总生物量的相关关系 Fig.3 Relationship between the biomass of filamentous diatoms and total biomass of phytoplankton
2.5 链状硅藻生物量的空间特征
链状硅藻生物量的空间分布特征显示(图5),除2015年12月外,其他时间段均呈现广州市周边站位明显高于其他站位,且最大值均出现在珠江桥站位,尤其是2012年12月(10.02 mg/L)和2015年3月(12.53 mg/L)几乎是其余站位生物量的100倍;西江沿线站位的生物量一般低于河网中部。2015年12月,广州市周边站位的生物量(均值为0.56 mg/L)与河网中部(均值为0.28 mg/L)接近,且高于西江沿线站位(均值为0.038 mg/L)。
从生物量相对百分比的空间特征来看,绝对优势种颗粒沟链藻在广州市周边站位的相对百分组成稳定在80%以上,受时间变动的影响很小;在西江沿线和河网中部的相对百分组成受时间变动的影响较大,枯水期的相对百分组成明显低于丰水期,这在2012年体现得最明显——该年丰水期颗粒沟链藻在西江沿线各站位的平均相对百分组成达到86.67%,在河网中部站位平均值为65.57%;而枯水期在西江沿线站位平均值为37.86%,河网中部站位的为46.80%。2012年的两个枯水期分别是变异直链藻(平均相对百分组成为分别为39.31%和8.11%)和芬兰直链藻(分别为1.87%和36.34%)在西江沿线和河网中部站位与颗粒沟链藻(分别为39.01%和45.65%)形成明显的竞争优势。2015年的两个枯水期,主要是变异直链藻(平均相对百分组成分别为30.12%和16.06%)在西江沿线和河网中部的个别站位与颗粒沟链藻(分别为66.04%和67.20%)形成明显的竞争优势(图6)。
2.6 链长的空间变化特征

图4 链状硅藻生物量的时间特征Fig.4 Temporal characteristics of biomass of filamentous diatoms (a.biomass; b.relative percentage)Agg:颗粒沟链藻Aulacoseira granulata (Ehrenberg) Simonsen;Aga:颗粒沟链藻极狭变种Aulacoseira granulata var. angustissima (O. Müller) Simonsen;Aaj:模糊沟链藻日本变型Aulacoseira ambigua f. japonica (F.Meister) Tuji & D.M.Williams;Aii:意大利沟链藻Aulacoseira italica (Ehrenberg) Simonsen;Ad:远距沟链藻Aulacoseira distans (Ehrenberg) Simonsen;Agas:颗粒沟链藻极狭变种螺旋变型Aulacoseira granulata var. angustissima f. spiralis (Hust.) Czarn. & D.Reinke;Aa:模糊沟链藻Aulacoseira ambigua (Grunow) Simonsen;Mv:变异直链藻Melosira varians C. Agardh;Mf:芬兰直链藻Melosira fennoscandica Cleve-Euler
总体上沟链藻属的链长要优于直链藻属(图7)。颗粒沟链藻链长的时空分布显示,各站位链长的最大值一般出现在丰水期,其链长优势较大的站位主要集中在河网中部及广州市周边。颗粒沟链藻极狭变种在河网中部和广州市周边站位的链长一般高于西江沿线站位。模糊沟链藻日本变型的链长明显大于其他沟链藻属,在河网中部站位优势较明显。其他沟链藻因出现率较低,链长未呈现明显的空间分布规律。直链藻的链长在13个站位差异较小且长度较短,时空特征不明显。
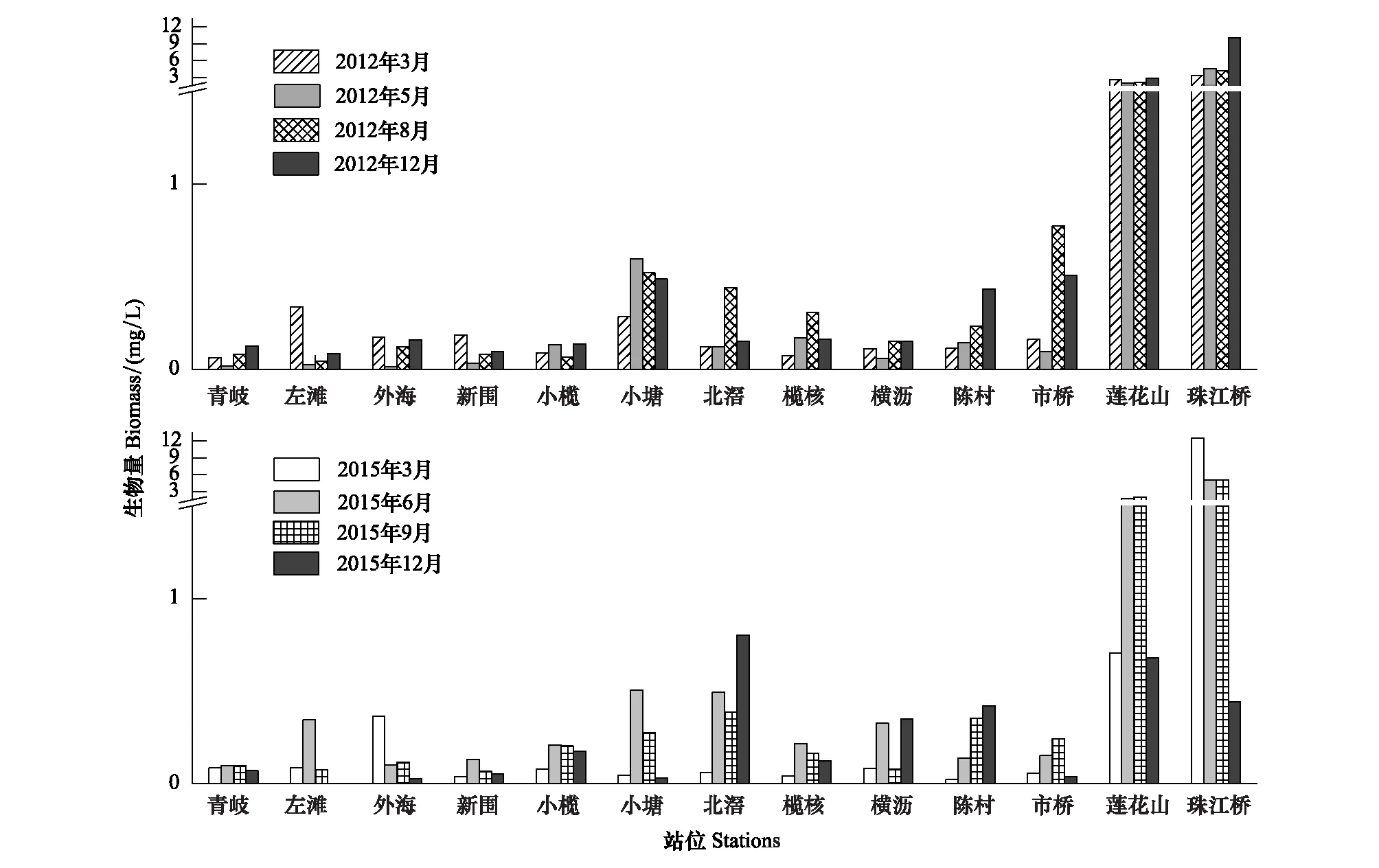
图5 链状硅藻生物量的空间特征Fig.5 Spatial characteristics of biomass of filamentous diatoms

图6 链状硅藻生物量相对组成的空间特征Fig.6 Spatial characteristics of relative composition of biomass of filamentous diatoms

图7 链状硅藻链长的空间特征 Fig.7 Spatial characteristics of filament length of filamentous diatoms
2.7 冗余分析结果
对不同链状硅藻生物量的相对百分组成与环境因子进行RDA分析(图8)。方差分析检验 (ANOVA test) 结果显示,轴1 (P=0.001)和轴2(P=0.852)对关联性的影响显著。轴1和轴2的特征值为466.47和70.19,分别可解释环境影响因子的76.94%和11.58%,合计解释率为88.52%,可以较好地解释不同链状硅藻生物量的相对百分组成与环境因子之间的关系。基于轴1的关联性结果显示,颗粒沟链藻与变异直链藻分布在轴1的相反方向,呈明显的负相关关系,水温和溶解氧是影响两者相对组成的最重要因素。基于轴2的关联性结果显示,pH是影响芬兰直链藻相对组成的重要因素。
对不同链状硅藻的链长与环境因子进行RDA分析(图8)。方差分析检验 (ANOVA test) 结果显示,轴1 (P=0.001)和轴2 (P=0.007)对关联性的影响显著。轴1和轴2的特征值分别为0.59和0.31,分别可解释环境影响因子的44.76%和23.93%,合计解释率为68.69%,可以较好地解释不同链状硅藻的链长与环境因子之间的关系。基于轴1的关联性结果显示,模糊沟链藻日本变型的链长与硅酸盐和氨氮呈正相关关系,与透明度和溶解氧呈负相关关系。基于轴2的关联性结果显示,颗粒沟链藻极狭变种与意大利沟链藻分布在轴2的相反方向,呈明显的负相关关系,水温、电导率和总溶解固体是影响两者链长的重要因素。

图8 生物量相对百分组成及链长与环境因子的RDA排序图Fig.8 RDA ranking map of environmental factors and relative percentage composition of biomass and filament length S:盐度Salinity;TDS:总溶解固体Total dissolved solids;WT:水温Water temperature;TP:总磷Total phosphorus;TN:总氮Total nitrogen;Trans:透明度Transparency;DO:溶解氧Dissolved oxygen;电导率: Conductivity
3 讨论
不同淡水流域的浮游植物群落结构研究都涉及到链状硅藻的多样性组成结果,例如Genkal等[25]对西西伯利亚平原东南部的河流及湖泊的研究发现,沟链藻7种和直链藻1种;Nardelli等[26]对伊瓜苏河下游和瀑布上游的研究发现,沟链藻9种和直链藻4种。相比而言,珠三角河网水域的链状硅藻物种多样性更为丰富,分析原因在于:(1)沟链藻属和直链藻属的很多物种是普生性种类,在世界范围内广泛分布[10, 12, 27]。珠三角河网地处亚热带沿海,属海洋性亚热带季风气候,全年温暖多雨;加上研究区域受人类活动影响频繁,水体营养丰富,利于链状硅藻的生长。(2)珠三角河网水域位于珠江下游,珠江三大干流(西江、北江和东江)在此汇集后入海,该研究区域的物种多样性存在累积效应,基本汇集了来自整个珠江流域的链状硅藻物种[28],因此物种多样性丰富。
除了物种多样性丰富,链状硅藻也较易在江河、湖泊、水库和河口等不同类型水域的浮游植物群落中占据优势地位[13-14, 29- 32],在珠三角河网水域也是最优势类群,分析原因在于:(1)链状硅藻具有多变和复杂的形态适应机制,细胞和藻链尺寸大小的变化及其相互之间的协同机制,可以有效地适应水环境的变化,维持种群的稳定性[21-22]。Passy[33]的研究结果发现,硅藻的形态复杂性与物种生态位宽度之间存在很强的正相关关系,这表明群体硅藻形态复杂性的增长可能是其适应未知环境的进化策略。(2)与单细胞微藻或尺寸小的微藻群体相比,链状硅藻的细长形态面临的摄食压力(例如浮游动物)相对较小。Fryxell等[34]在研究中总结出的硅藻链状结构的功能之首便是防止浮游动物摄食。(3)部分链状硅藻物种(例如颗粒沟链藻)在光线较弱的底层环境中依然能够继续生长,保持了种群数量和丰度,随着水体波动再悬浮,维持了种群优势的稳定性。有研究人员发现,风力引起的水体混合作用可以影响高纬度浅层湖泊中的链状硅藻生长状况[35]。(4)人类活动引起的水体富营养化在世界范围内普遍存在。硅藻在营养丰富的条件下具有较高的生长速度,因此其细胞分裂速度比其他类群快[36-37],有研究表明在富营养水体中,模糊沟链藻、颗粒沟链藻和颗粒沟链藻极狭变种等链状硅藻经常出现并占据优势地位[5, 38- 41]。
珠三角河网的结果显示,沟链藻的优势远大于直链藻,这与国内外的研究结果一致。分析原因,主要是由形态结构的差异造成的:(1)沟链藻属物种的结构更加稳固,表现为细胞之间通过短节状的互锁的棘突将圆柱状藻壳连接在一起;而直链藻属细胞之间的连接主要由粘液垫和小的不规则的刺在壳盘缘组成[9]。(2)部分沟链藻属物种的端细胞具有长形刺等辅助结构,可以有效地减缓沉降速度。Conway等[42]研究发现,具有刺状结构的栅藻的沉降速度比不具刺的更慢一些。(3)Wang等[22]研究发现,某些沟链藻属物种(如颗粒沟链藻、模糊沟链藻日本变型和颗粒沟链藻极狭变种螺旋变型等)可形成稳定的弯曲状态,有效应对水体波动并减缓摄食压力。以上形态结构上的特点使沟链藻更适应环境变化,有利于其在表层水体中形成稳定优势。
链状硅藻生物量的时间特征显示,枯水期的差异远大于丰水期,且最大值和最小值均出现在枯水期。分析原因在于:(1)丰水期河水径流量大,对藻类群落有明显的稀释作用,降低了生物量。王超等[43]对西江下游浮游植物群落周年变化的研究发现,种群丰度的最大值均出现在河水径流量的高峰之后的枯水期。(2)丰水期的水流量大,流速快,滞留时间短,藻类对可溶性养分的吸收率较低,不利于生物量的增长;反之,枯水期的低流速和高滞留时间有助于生物量的增长[44-45]。(3)一般情况下,枯水期水体透明度高,链状硅藻的沉降速度大,易造成生物量的沉降损失。但是,生物量最大值出现在了透明度较高的2012年12月(57.7 cm)和2015年3月(77.3 cm)。环境数据显示,这两个枯水期的盐度是另两个枯水期的4—6倍,总溶解固体浓度也呈现相同规律,RDA结果显示(图8),本研究流域的绝对优势种——颗粒沟链藻与盐度和总溶解固体也呈正相关关系。王超等[20]发现,颗粒沟链藻可以在珠江河口的半咸水环境中占据优势地位,说明其对盐度有一定的适应性;也有报道指出,总溶解固体能够促进硅藻的生长,增加硅藻细胞密度[46-47]。表明总溶解固体和盐度很可能是影响链状硅藻生物量的重要因素。
链状硅藻生物量的空间分布显示,莲花山和珠江桥站位显著高于其他站位(2015年12月除外),主要原因在于(1)广州市周边水域受人类活动影响大,珠江桥站位的总氮和总磷含量是其他站位的2—3倍,富营养化程度极高,有助于链状硅藻形成极高的丰度。(2)西江沿线生物量一般低于河网中部站位,主要原因为西江沿线站位受到西江径流稀释及冲击的作用,加之西江沿线站位营养盐含量低,水体透明度较高等特点,导致硅藻出现沉降损失[28]。而2015年12月的低生物量很可能与降雨量及光照条件有关,当月有15天有雨,部分时间有大到暴雨,晴天只有3天(当地天气见网站http://lishi.tianqi.com/guangzhou/),温度下降,光照条件不足以及降雨量增加,很可能是导致生物量骤降的主要原因。整体上,链状硅藻生物量的空间分布格局主要与人类活动造成的水体营养状况有关。
不同链状硅藻生物量相对百分比的时空特征显示,颗粒沟链藻是绝对优势种群。O′Farrell等[48]研究显示,颗粒沟链藻喜好浑浊、富营养化的水体,是中营养-富营养化水体的良好指示物种。至于颗粒沟链藻在枯水期的优势度出现较大波动,除了因透明度增大导致的沉降损失、以及低温不利于其生长等因素外,变异直链藻的种群增长也造成了颗粒沟链藻的相对弱势,图8中颗粒沟链藻与变异直链藻沿轴1呈对立分布,也显示了两者的竞争关系。变异直链藻是典型的底栖硅藻,在水温较低时更适生存,本研究中其与水温呈负相关关系也证明了这一点,贾兴焕等[49]发现,附石藻类细胞密度在冬春季节要高于夏秋季节,表明低温更适合附石藻类的生长。在枯水期水位较低,水体扰动的情况下,变异直链藻易悬浮至水体表层,形成种群优势。此外枯水期溶解氧含量上升更适于变异直链藻生长,Nardelli等[9]在研究中指出变异直链藻的生长需要适度氧气,图8显示变异直链藻与溶解氧呈现极显著的正相关关系也说明了变异直链藻对氧气的依赖性;而颗粒沟链藻则相反,该物种则更适于生存在溶解氧含量较低的中污水体或重污水体中[50]。因此在某些站位,变异直链藻与颗粒沟链藻形成明显的竞争优势。
链状硅藻链长的时空特征显示,沟链藻属的链长整体上大于直链藻属,主要源于形态结构上较稳固的优势,前面已有论述。时间特征显示,颗粒沟链藻在丰水期的链长一般大于枯水期,其主要原因为(1)链长越长,沉降率越高,丰水期水体透明度较低,一定程度上降低其沉降率[4],因此表层水中易出现长链。本研究中颗粒沟链藻链长与水体透明度呈负相关关系也证实了这一点;(2)丰水期河流流量大,水流湍急,可以使颗粒沟链藻从底层再悬浮至表层水体,而在底层的颗粒沟链藻大多是易沉积的长链[51];(3)丰水期光照时间较长,也可能引起颗粒沟链藻链长的增加,实验表明[4]光周期的增加会导致硅藻生长和链长增加,O′Farrell等[48]在研究中指出,当营养盐含量充足,光照条件受限制时,颗粒沟链藻的藻链长度会因此减短。其余藻链长度的季节变动不稳定,很可能与这些沟链藻属物种的形态特异性(如颗粒沟链藻极狭变种的直径很小,模糊沟链藻日本变型呈螺旋形态)有关。
链长的空间特征显示,颗粒沟链藻和颗粒沟链藻极狭变种的链长在河网中部和广州市周边站位大于西江沿线,根据图9显示二者与pH、溶解氧和透明度呈明显的负相关关系,与水温、盐度和总溶解固体呈正相关关系,主要原因是:(1)浮游植物通常在pH值为7—9的弱碱性水质中生长,酸性或碱性过强都会抑制其光合作用[52],Celestino等[52]等发现适合链状硅藻生长的最佳pH范围是7.99±0.30,O′Farrell等[48]发现在pH值范围为6.5—7.8的研究水域中,颗粒沟链藻的平均细胞直径与pH值呈反比,在本研究流域中pH值范围为7.25—8.37,颗粒沟链藻和颗粒沟链藻极狭变种的藻链长度与pH值呈反比,表明pH是影响其藻链尺寸的重要因素。(2)二者链长与溶解氧的负相关关系以及与水温和总溶解固体的正相关关系很可能是因为链长对生长周期的响应。研究表明,颗粒沟链藻由中间连接细胞和末端分裂细胞组成,其连接细胞通过短而互锁的棘突紧密连接在一起,从而导致链的形成,末端分裂细胞的壳盘缘上具有长而尖细的长刺使连接细胞彼此滑落分离,因此藻链的长短依赖于种群中分裂细胞的产生频率来控制,而沟链藻生长速率的增加往往伴随着分裂细胞产生频率的下降,从而导致藻链长度增加[53]。颗粒沟链藻和颗粒沟链藻极狭变种通常在溶解氧含量较低的富营养水体中生长速率较高,总溶解固体含量和水温的升高也能促进二者生长速率的升高(前文已有论述),从而导致藻链长度的增加。(3)水体透明度升高会导致藻链沉降速率增加,从而引起表层水中藻链长度的下降。模糊沟链藻日本变种的链长在河网中部站位优势较明显,根据图9显示该种与硅酸盐及氨氮呈极显著正相关关系。Poister等[53]指出水中磷元素含量较低时,不仅会导致模糊沟链藻含量降低,还会引发其链长减短,而增加的磷可利用性有助于形成具有更高沉降速度的、链长更长的模糊沟链藻群落。Conley等[54]指出水柱中可溶性反应性硅(DRSi)不断消耗时会引发链状硅藻沉降,进而通过沉淀作用获得更多养分,较大的硅藻正是导致较高沉降率的重要因素,它的这种生存策略也会降低系统中其他生产力对养分的利用,从而具有极强的种间竞争力。
4 结论
(1)珠三角河网流域共鉴定出链状硅藻18种(含变型),包括沟链藻属11种和直链藻属7种,该流域的沟链藻属比直链藻属更具有优势地位。在该流域中颗粒沟链藻是绝对优势种,也是水体富营养化的重要指示物种。
(2)颗粒沟链藻和变异直链藻在某些站位形成竞争优势。水温、营养盐、溶解氧及透明度等环境因子是影响链状硅藻生物量的重要因素。
(3)一般情况下沟链藻属的链长要大于直链藻属,透明度、pH值、营养盐和水温等环境因子是影响藻链大小的重要因素。
(4)链状硅藻在空间上的差异性是由于不同采样地的地理位置不同造成环境因子具有差异性,进而影响硅藻的生物量及链长。
猜你喜欢
——中核工程创新驱动改革,奋力打造核工程产业链“链长”
中国工程咨询(2022年8期)2022-08-31
河南科技(2021年14期)2021-11-28
中国纺织(2021年12期)2021-09-23
黑龙江水利科技(2020年8期)2021-01-21
中国卫生产业(2020年25期)2020-11-30
飞天(2020年9期)2020-09-06
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
电脑知识与技术(2018年7期)2018-06-06
分析化学(2018年2期)2018-03-02
作文周刊(中考版)(2017年21期)2017-07-10
