
油菜响应水分胁迫的生理机制及栽培调控措施研究进展
2021-04-21 07:59杨海云艾雪莹BatoolMaria刘芳蒯婕王晶汪波周广生
华中农业大学学报(自然科学版) 2021年2期
杨海云,艾雪莹,Batool Maria,刘芳,蒯婕,王晶,汪波,周广生
1.华中农业大学植物科学技术学院/农业农村部长江中游作物生理生态与耕作重点实验室,武汉 430070;2.全国农技推广中心油料处,北京 100125
油菜(BrassicanapusL.)是世界上重要的油料作物,也是重要的饲用蛋白原料。油菜籽粒中含有约61.0%的油酸和8.8%的亚油酸,菜籽油营养品质好,是健康的食用植物油之一[1]。我国油菜种植面积和总产量近世界总量的三分之一,每年可提供约520万t优质食用菜籽油[2],油菜的优质高产栽培对于我国乃至世界油料市场的供给意义重大。
水分是作物生长的必需因子,对作物的生存起着决定性的作用。不适的水分条件往往会导致作物生长发育不良,影响作物的产量与品质。水分胁迫主要分为干旱和淹水2种。干旱胁迫在作物生产中普遍发生,水资源短缺是导致干旱的重要原因[3]。目前世界干旱和半干旱地区已达到地球土地总面积的三分之一以上[4],严重制约了全球作物生产。淹水胁迫是因水分过多对植物生长发育产生不良影响的一类水分胁迫。淹水胁迫是目前世界湿润地区、半干旱地区植物生长过程中普遍面临的问题,世界上约16%的农业生产区受洪水危害[5]。播种后强降雨事件的发生和土壤排水不良是造成淹水胁迫的主要原因[6]。
长江流域是我国油菜的主产区,该区域全年降水不均,易出现季节性干旱和淹水问题。我国西南西北的油菜种植区同样也面临干旱天气的影响。据统计,秋旱可使我国长江流域油菜总产量降低 25%~32%[7],而淹水胁迫则威胁到我国20%种植面积的油菜[8]。即使在生长期遭遇短期的水分胁迫,油菜的产量也会受到影响。若遭遇严重的水分胁迫,则会引起油菜整株死亡或严重的产量损失。解析油菜生长过程中响应水分胁迫的生理生化机制,对于优化油菜栽培方式及培育抗性品种具有重要的指导意义。
1 水分胁迫对油菜生长发育的影响及油菜响应机制
油菜对水分胁迫较敏感。首先,油菜的萌发立苗及形态建成受水分条件制约[9]。其次,植株的光合作用、代谢物质积累等生理生化过程在不适的水分条件下会发生紊乱[10],最终影响产量和品质。油菜植株除了改变叶片及根系形态结构,还能通过一系列生理生化响应来适应水分胁迫(图1)。
1.1 水分胁迫对油菜种子萌发出苗的影响
水分是影响作物种子萌发和幼苗形态建成的关键因子。播种后,土壤的水分条件决定了种子的发芽率和发芽速度,影响作物成苗[11]。研究表明,干旱胁迫下种子发芽速度变慢,并且随着干旱胁迫程度的增加,发芽速度严重降低,主要表现为平均发芽时间延长和发芽指数降低[12]。干旱胁迫同样干扰油菜种子萌发进程,干旱条件下种子的活力会逐渐丧失,发芽率下降,平均萌发时间变长,最终造成油菜出苗不齐、成苗率低等一系列问题,导致油菜产量下降。而抗旱性较强的油菜品种能在较低的土壤水分条件下维持较高的出苗速度与出苗率,获得高活力的幼苗[13]。
在油菜-水稻轮作区域,由于稻田土壤湿润,若遇秋季持续降雨,油菜播种时将面临土壤水分过多的问题,引起渍害,导致出苗率降低,秧苗较弱。研究表明,油菜种子经过12 h的缺氧胁迫后,存活率和相对活力指数均呈降低趋势[14]。Cheng等[15]对不同基因型甘蓝型油菜种子露白后进行12 h密闭淹水处理,也得到了相同的结论:与对照相比,淹水处理对油菜萌发出苗具有显著的抑制作用,而电导率可以作为耐淹材料筛选的重要指标。该研究鉴定出了 03P79-1、03P62-5、中双8号等耐淹性较强的品种,可以作为耐淹性资源加以利用。
1.2 水分胁迫对油菜地上部生长的影响及油菜响应机制
1)地上部生长。大量研究表明,植物的株高、茎粗、地上部鲜质量及干质量均受水分条件影响。在营养生长初期,干旱胁迫下油菜的苗高和鲜质量与水分条件显著相关。与正常水分条件下生长的幼苗相比,经10% PEG-6000渗透胁迫后,油菜幼苗苗高降低40.68%,鲜质量降低34.2%[16]。苗期对水分胁迫的耐受性对油菜整个生育期的良好发育至关重要,Khan等[17]研究发现干旱条件下油菜苗期地上部干质量与产量或产量抗旱系数呈极显著正相关。也有研究表明,油菜薹期至花期对水分胁迫最为敏感[18],花期若遇干旱,敏感型油菜品种的株高显著下降[19],而耐旱油菜品种往往能够更好地维持地上部的生长,株高几乎不受水分影响[20]。
淹水胁迫引起的水氧竞争会干扰油菜地上部正常的生长发育。油菜幼苗在淹水3周后,苗高、茎粗、绿叶数、单株叶面积较正常水分处理下降,地上部分鲜质量、干质量显著降低[21]。何激光等[22]分别在油菜苗期、花期、蕾薹期、角果成熟期进行了3、5 和7 d的淹水处理,发现淹水会使油菜的株高、茎粗明显降低,但降低程度在不同时期存在差异,油菜地上部分的生长受苗期和蕾薹期淹水处理影响最大。
2)叶片光合生理。叶片是作物产量的重要决定因素之一,直接影响光合作用期间的同化物产量。光合作用是作物生长发育的物质基础,常以直接或间接的方式为植物的生长发育提供能量。而植物叶片形态与功能的维持对水分条件十分敏感,并易受到水分胁迫的不利影响。
①叶片形态。叶片形态的变化决定了油菜的有效光合面积。水分胁迫首先会引起叶片相对含水量(RWC)的变化,从而干扰叶片正常生长和气孔传导,进而影响光合作用,最终导致作物干物质积累量降低[23]。谢小玉等[24]发现经过干旱处理后,油菜苗期叶片RWC、叶面积(LA)均会降低,且随干旱胁迫时间延长和胁迫程度增加,叶片RWC和LA降幅不断增加。因此,干旱胁迫下叶片含水量变化[25]、萎蔫指数[26]可作为油菜苗期抗旱筛选指标。淹水胁迫同样也会改变叶片形态,受淹后油菜LA与SPAD值显著下降[27]。高华东等[28]发现,在薹期进行淹水处理会使油菜叶片黄化萎蔫,且不同品种间的叶片衰老程度存在差异。
②叶片功能。在水分胁迫下,植物体渗透调节及抗氧化系统对于维持叶片正常功能十分重要,间接决定了叶片光合性能。在苗期干旱胁迫下,油菜叶片丙二醛(MDA)、脯氨酸(Pro)、可溶性糖、可溶性蛋白含量以及超氧化物歧化酶(SOD)、过氧化物酶(POD)活性相对值均表现出上升趋势[24]。研究发现,叶片可溶性糖与油菜品种抗旱性密切相关,碳水化合物的积累有助于叶组织的渗透调节,这种渗透调节有利于提高油菜抗旱性[29-30]。而在淹水胁迫下,油菜叶片中蔗糖和可溶性糖含量升高,这是由于蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)活性增加。说明一定程度的淹水促进光合产物向蔗糖方向分配,增加可溶性糖含量,提高渗透调节能力,缓减生理失水,从而适应水分胁迫。研究还发现,淹水胁迫后,膜脂过氧化程度加剧,MDA含量显著上升。同时,油菜叶片中活性氧代谢失衡,SOD活性、抗坏血酸(AsA)和谷胱甘肽(GSH)含量升高,其中敏感品种较抗性品种变化幅度更大[31]。
叶绿素是植物吸收和利用光能的主要色素[32-34],其含量的变化可以反映初始光合反应特点,尤其是叶绿素a直接影响了光合活性及光饱和光合速率(Pn)[35]。干旱条件下植株叶片中叶绿素会遭到破坏,同时光合酶的含量和活性均会降低[36]。与正常水分条件相比,缺水条件下光系统Ⅱ的最大量子产量(Fv/Fm)值下降,光合速率迅速降低[37]。不同基因型甘蓝型油菜光合特性受缺水的影响存在差异。Dai等[38]的试验结果表明,干旱降低了敏感型油菜Pn、气孔导度(Gs)、水分利用效率(WUE)和叶绿素含量,光系统Ⅱ的光量子捕获效率(Fv'/Fm')、实际光化学效率(PhiPSⅡ)、光化学效率(qL)和PSⅡ(光系统Ⅱ)非环式光合电子传输速率(ETR)均会下降;而耐旱型基因型中,仅光合速率和叶绿素含量,Fv'/Fm'和qL值有所降低。淹水胁迫下,油菜叶片叶绿素a、叶绿素b、叶绿素a/b和类胡萝卜素含量均会降低,光合作用受限[27],但影响程度因淹水时间、淹水强度和基因型等表现不同[39]。在淹水早期,由于气孔限制,光合速率急剧下降,但水分消退后,植株表现出较高的恢复能力;若淹水时间过长,PSⅡ受损,光合作用则受到不可逆损伤[40]。Wan等[41]发现油菜终花期淹水后叶片中Rubisco活性下降,且降幅随淹水时长不断增加,同时叶片Pn、Gs、胞间CO2浓度(Ci)和蒸腾速率(Tr)等均显著降低。
1.3 干旱胁迫对油菜根系生长的影响及油菜响应机制
根系生长发育及其在土壤中的时空分布不仅决定了作物对水分和养分的吸收[42],而且直接影响到地上冠层的建立、同化物的分布和最终产量[43-44]。前人研究表明,较大和较深的根系有利于水分吸收[45];然而,近年研究表明适宜大小和较好生理功能的根系更有利于养分的吸收[46],根系过大导致冗余,使得干物质分布不平衡[47],也会造成不必要的养分消耗。
1)根系形态。根系是油菜水分及养分吸收的主要器官,与油菜的抗旱性关系密切。油菜根系表型指标主要有:根系体积、根系平均直径、主根长、侧根长、根总长度、根尖数[48]。
干旱使油菜根质量及侧根数目显著降低,且随干旱强度增加,根长递减[49],耐旱性品种降幅低于敏感型品种[50]。根系伸长是作物响应干旱的关键行为,可帮助植物从深层土壤中获得水分,从而缓解干旱带来的伤害[51],这是干旱胁迫下根冠比增加的主要原因。前人研究发现,油菜苗期适度控水胁迫会引起侧根数减少,但其平均侧根长度增加[52-53],更能适应不良水分环境。在培育抗旱品种时,根系对土壤水分的响应可作为关键筛选指标[54-55]。通常耐旱品种根系较敏感型品种更为发达,具有较大的根系生物量、总根长、总根系表面积。
根是第一个面临氧张力降低的器官,在淹水条件下,与地上部相比,经历了显著的表型变异和更大损伤[56]。连续淹水后油菜苗期根系生物量、根表面积、根体积、主根长和总根长、侧根数量显著减少,平均根直径显著增加[57]。5叶期淹水21 d后,油菜幼苗根生物量降至对照的50%[58]。Darwent[59]比较了2个不同抗性的油菜群体,发现耐淹群体比敏感群体在主根上形成更多的侧根。
2)根系结构。根系结构和解剖表型对作物适应干旱有重要意义。根系解剖表型与根系的径向和轴向水分运输密切相关,从而影响作物水分吸收和分配的效率[60]。木质部导管性状(数量、直径和面积)影响轴向导水率,而皮层性状和细胞亚硝化层的存在可能影响径向导水率。较大的木质部导管和较粗的根是作物耐旱的特征[61]。Yin等[62]通过对油菜根系冰冻切片组织化学染色发现,与对照相比,干旱胁迫下生长的油菜根中木质部显著加厚,染色更深。同时根中的导管孔径和导管数量均明显增加。此外,根皮层通气组织(RCA)的形成对作物抗旱也有重要意义,RCA可以显著降低根部代谢成本,促进根部生长并促进植株从干燥土壤中吸水,从而提高抗旱性。Zhu等[63]研究发现干旱胁迫使玉米的RCA从54%提高到77%,且高RCA基因型籽粒产量比低RCA基因型高78%~143%,油菜中暂未见报道。
植物在淹水诱导的缺氧条件下,根系会通过细胞程序性死亡将活的皮层细胞转化为空气空间,也就是RCA。RCA的形成具有改善缺氧条件下氧气向根部输送的功能[64-65],减少植株的呼吸需求,增加内部孔隙度,从而缓解缺氧对植株的伤害。研究发现植物形成通气组织的能力越强,抗淹能力越强[66-67]。已有研究表明,油菜与其他作物在耐淹性方面存在差异,在研究作物缺氧的适应机制中观察其根系结构时发现,甘蓝型油菜无法形成RCA,而小麦具有20%~22%的通气组织作为对淹水的响应,在淹水期间光合作用得以维持。大麦不定根含有19%的气孔,而油菜根系不形成气孔[68-69]。Voesenek等[68]研究了2个甘蓝型油菜品种根系对缺氧的响应,发现甘蓝型油菜根系孔隙度低至3%~5%,是耐淹性低的典型表现。在淹水油菜的根系中,中柱氧浓度低,皮层氧浓度稍高。尽管根皮层组织中存在连续的胞间空气空间,但缺乏通气组织,导致根系内部氧扩散速率低,限制了根在缺氧条件下的生长,这可能是甘蓝型油菜对淹水敏感的重要原因之一。
3)根系功能。干旱条件下,作物会通过根系功能物质的改变维持生存,主要表现为渗透调节物质的含量变化及抗氧化机制的启动。研究发现,在轻度和中度干旱胁迫下,植物根系可溶性蛋白、脯氨酸和可溶性糖含量增加。随着胁迫程度的增加,根系MDA含量和质膜透性逐渐升高。同时,根系POD、SOD和AsA在低中度胁迫下加深,重度胁迫下降低。而复水后根系抗氧化防御系统和渗透调节物质又迅速恢复到对照水平[70]。干旱增加了2个基因型油菜根系中MDA和过氧化氢(H2O2)含量。为防止细胞进一步氧化受损,油菜根系抗氧化酶受干旱刺激后迅速上升,POD、SOD和CAT均有不同程度的增加[38]。

2 水分胁迫对油菜产量和品质的影响
干旱和淹水是限制油菜产量和品质的关键非生物胁迫因子,其影响程度因选用的试验品种、胁迫时期、胁迫强度等的不同存在较大差异。总体而言,在水分胁迫下,油菜产量降低,品质变差(图1)。
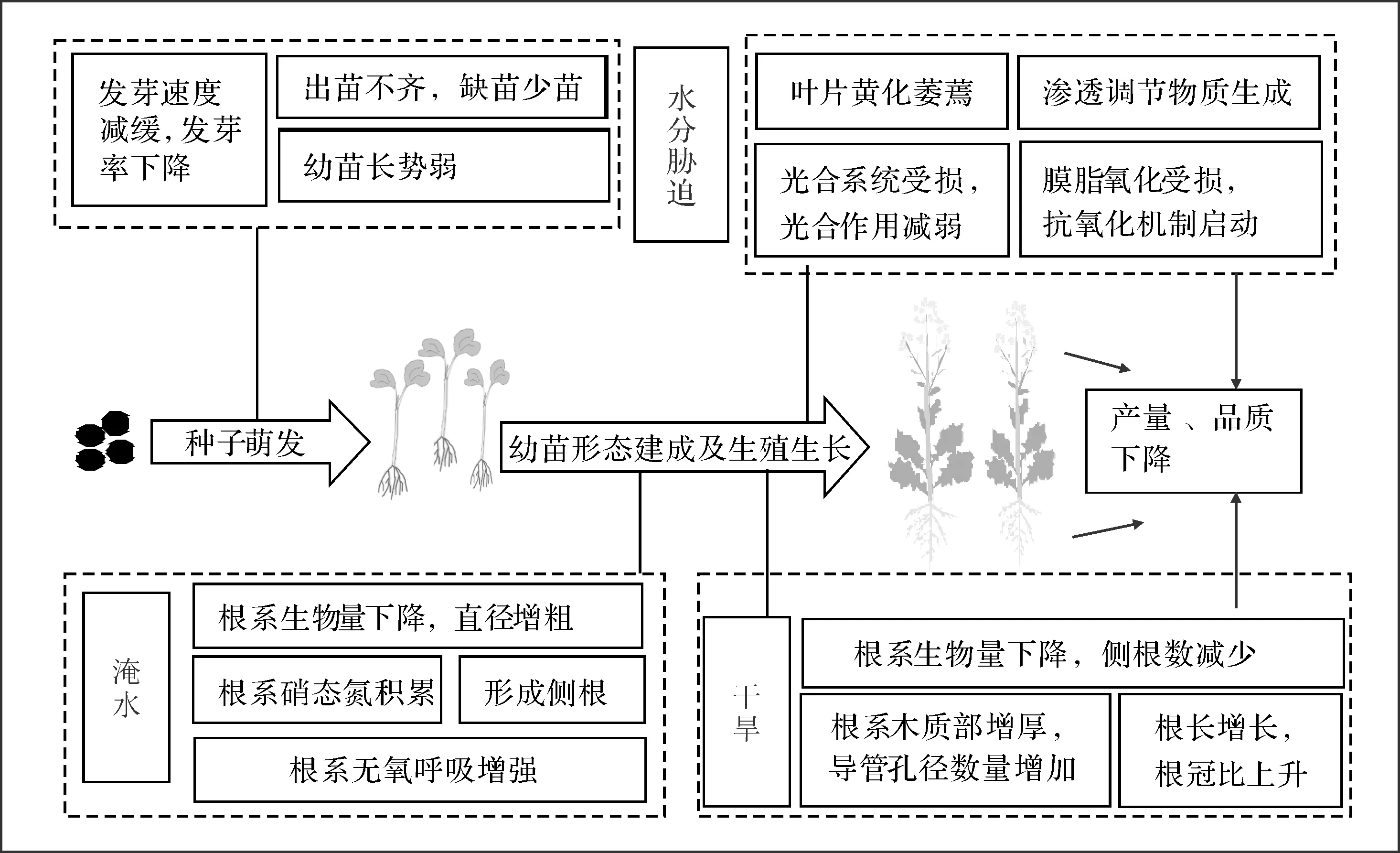
图1 水分胁迫对油菜生长的影响及响应机制Fig.1 Effect of water stress on rapeseed(Brassica napus L.) growth and its response mechanism
2.1 产 量
油菜产量和产量构成因素受水分影响较大,薹期到成熟期不灌溉籽粒产量显著降低[75-76],而花期和角果发育期干旱对油菜产量影响最大[77-78],花期干旱显著延长了开花时间,减少了单株角果数,减产幅度在32%~48%[29,79],干旱主要通过减少单株角果数来影响产量[80]。淹水对油菜产量的影响程度因生育期和品种的敏感程度不同存在差异:淹水后,油菜产量下降17.0%~42.4%,严重水分胁迫则会引起植株死亡而绝产[8, 81]。不同生育期中以萌发期最为敏感,减产幅度最大;其次为开花期、苗期、角果期和成熟期[26]。Li等[82]对15份甘蓝型油菜材料进行了大田苗期淹水处理,淹水后有效分枝数、单株角果数、每角粒数、单株产量均显著下降,其中有效分枝数比对照减少31.81%~78.02%,耐淹基因型产量降幅低于敏感型品种。
2.2 籽粒品质
作物籽粒品质是由淀粉、蛋白质和脂肪等物质的比例决定的,受水分条件等环境因素的影响[83]。成熟的油菜籽粒中含有30%~50%的脂肪、21%~30%的蛋白质,脂肪和蛋白质的生成会随着水分变化发生相应改变[84-85],变化程度因生育期而异。开花期进行干旱处理后籽粒含油量降低6%,而成熟期干旱对含油量的影响更为突出,此阶段干旱处理比对照含油量低10%以上。籽粒蛋白变化与含油量规律相反。同时,种子硫苷含量在水分亏缺下增加了60%[86]。苗期淹水后,油菜籽粒品质变化规律与干旱一致,即含油量降低、蛋白质含量上升[85]。另有研究表明淹水不仅会降低油菜籽粒含油量,还能影响其脂肪酸组成。Xu等[87]研究发现淹水会降低油菜籽粒中亚麻酸(C18∶3)和亚油酸(C18∶2)含量,同时,淹水通过增加芥酸(C22∶1)和硫代葡萄糖苷含量来影响油质。
3 提高油菜耐水分胁迫能力的措施
3.1 加强品种选育,选育优质抗逆品种
油菜生产区域的物候环境具有多样性,不同品种之间的耐水分胁迫能力也存在着较大差异。与其他表型相似的材料相比,耐旱油菜材料具有更有效的分子保护机制,如活性氧清除、调控源/库比和发育时间。耐渍型强的油菜幼苗,硝酸还原酶(NaR)和亚硝酸盐还原酶(NiR)将硝酸盐还原为NO,随后非共生血红蛋白Hb将NO转化为NO2,提供了维持淹水根系生物能的机制[88],这些研究为油菜耐淹抗旱育种提供了借鉴。
3.2 合理排灌
油菜产区一般秋冬降雨偏少,土壤干旱,不利于播种出苗和培育壮苗。而长江中下游地区雨水偏多,常超过油菜正常需水量,加之该产区主要采用水旱轮作尤其是稻茬免耕的种植模式,地下水位高,土壤黏重,易发生淹水胁迫危害,导致油菜产量下降。因此,控制土壤中适宜的含水量对油菜生长尤为重要,合理灌排是保证其高产稳产的重要措施[89]。宋丰萍等[79]通过试验明确了油菜苗期适宜地下水位为 30~90 cm, 蕾薹期、花期为 30~60 cm,角果发育成熟期为30~90 cm,就全生育期而言,地下水位控制在 30~60 cm 时能满足油菜生长发育及产量形成需求。另外有研究证明,连续地下淹水对油菜产量影响最敏感的生长期是花期和角果期,做好油菜田春季排水管理对油菜生产具有重要意义[90]。
3.3 种子引发
种子引发也称播种前处理,即控制种子缓慢吸收水分使其停留在吸水的第二阶段,让种子进行预发芽的生理生化代谢和修复[91]。近期研究表明,种子引发可以提高油菜种子的萌发能力与抗逆性,与未启动的种子相比,经褪黑素引发后的油菜种子萌发率和出苗率显著提高[92]。盐引发可以提高种子萌发速率,同时增强幼苗抗逆性,王建成等[93]发现在20%和25%的水分条件下,Ca(NO3)2启动处理显著提高了浙油758的发芽势和发芽率。
3.4 施用生长调节剂
水分胁迫能够通过干扰植物激素的内源水平和信号传导途径,显著影响植株的生长,最终降低产量。而外源施用不同的植物生长调节剂可以调节有关生理生化过程,从而减轻水分胁迫对作物的不利影响。应对水分胁迫常见的植物生长调节剂有水杨酸、ABA、油菜素甾醇、乙烯利、烯效唑S-3377、多效唑PP333等[94-96]。Leul等[58]发现施用烯效唑可显著提高油菜幼苗在淹水胁迫下的叶片光合速率、生物量,增加分枝和有效角果数。多效唑(PBZ)处理也是提高油菜幼苗耐淹能力的有效措施[97]。5-氨基乙酰丙酸(ALA)是一种潜在的植物生长调节剂,可有效对抗各种非生物胁迫引起的伤害。植物对ALA积累可以提高抗逆性[98]。
3.5 合理施肥
叶面施氮可显著提高淹水胁迫环境下油菜的产量[99]。硝酸还原酶是氮代谢中的第一个酶,也是硝酸盐同化中的限速酶[100]。硝酸盐是油菜吸收和储存的主要氮素形式,高硝酸盐含量有利于油菜生长[101]。研究发现,与对照相比,经施氮处理后,受淹油菜品种GH01籽粒含油量略有增加,ZS 9籽粒含油量显著增加[27]。叶面喷施钾肥同样也能减轻淹水对油菜的不利影响,并延缓冬季叶片衰老。Wan等[41]在开花期对油菜进行了淹水胁迫,随后进行叶面喷钾,发现施钾增加了Rubisco在淹水胁迫下的活化,从而显著减轻了淹水对植株光合作用的抑制。
4 研究展望
油菜是食用植物油主要来源之一,优质高产高效油菜生产对保障食用植物油安全供给具有重要作用。受全球气候变暖影响,目前农业生产中水分逆境频发,严重影响了油菜产量和品质的提升。较多学者对油菜水分逆境进行了研究,近年的研究热点主要集中于三个方面:(1)油菜各生育期抗性评价与耐性种质筛选[102-105];(2)水分逆境下油菜地上部光合生理、渗透调节、酶活变化等有关生理响应机制[57,106];(3)油菜耐水分胁迫栽培调控措施。当前以种子引发[91-92]、应用植物生长调节剂[94-95]及施肥[99]3种方式可操作性最强。
目前,上述研究虽已取得了一定的进展,但仍存在一些问题。植株耐旱(淹)能力是一个综合的性状,前人总结的评价体系多以发芽指标与农艺性状为主,而这些指标常因处理时期与胁迫程度的不同而存在差异,从而很难得到一套统一的标准。为全面而准确地评价品种的耐水分胁迫能力,遗传分析、分子标记及基因表达等方法的应用十分必要。Khanzada等[107]以油菜为材料,利用全基因组关联研究(GWAS)鉴定出314个与干旱指数密切相关的标记,分布在A和C基因组的所有19条染色体上,这些标记可以用来鉴定油菜品种的抗旱性,而具体的应用还需进一步研究证实。另外,油菜对水分胁迫的响应是地上部与地下部综合作用的结果。根系形态结构是植物发育水平的重要体现,与生理功能关系密切[108]。根系木质化程度、输导组织、表皮附属结构等会影响植物耐水分胁迫能力[109],导管直径大小、通气组织(RCA)会影响根系生理机能以及对水分的吸收和运筹能力[110]。但从现有的研究来看,与其他作物相比,油菜在该领域的研究显得相对薄弱,需加强研究。
油菜干旱、淹水环境的根-冠响应是一个复杂的生物学过程,涉及到分子、器官、个体、群体的复杂效应,多条信号通路交叉调控。随着微生物-植物互作、多组学、纳米技术等领域不断取得突破性进展,将这些手段应用在油菜响应水分逆境胁迫机制及辅助栽培调控技术的研究上,将大大加快油菜逆境响应机制的解析进度,并有望实现更为精准高效的抗逆栽培调控。
猜你喜欢
今日农业(2021年21期)2021-11-26
今日农业(2021年14期)2021-10-14
四川劳动保障(2021年3期)2021-06-09
今日农业(2020年20期)2020-12-15
今日农业(2020年17期)2020-12-15
世界农药(2019年4期)2019-12-30
娃娃乐园·综合智能(2018年3期)2018-03-22
现代园艺(2017年13期)2018-01-19
数学小灵通(1-2年级)(2017年10期)2017-11-08
上海农业学报(2016年2期)2016-10-27
