
天台县大雷山夏蜡梅群落学特征分析
2021-04-21 01:18张忠钊谢文远张培林
浙江农林大学学报 2021年2期
张忠钊,谢文远,张培林
(1. 天台县自然资源和规划局,浙江 天台 317200;2. 浙江省森林资源监测中心,浙江 杭州 310020)
夏蜡梅Sinocalycanthus chinensis是1963年发表的新种[1],1964年又被提升为新属[2]。该种是中国特有的第三纪古老孑遗植物,目前仅在浙江、安徽两省有少量分布,数量稀少,在分子上有保守的叶绿体基因组,在分类地位上与美国蜡梅属Calycanthus又有着很深的渊源,对阐明东亚—北美植物区系历史的发展和联系很有意义[3]。近年来,大量研究人员对它的种群分布、生理生化以及园林、药用价值等方面作了深入研究[4-9],但对种群结构和群落种间联结的研究未见报道。天台县境内的夏蜡梅则是1959年阙良寿在大雷山首次发现(阙良寿28677,浙江省自然博物院植物标本馆)。天台大雷山是夏蜡梅的第四纪冰期的避难所之一(另一个是临安大明山),与临安不共享一个单倍型(h3),且多样性较低,表明天台县夏蜡梅的种群是比较孤立的,是一个重要的种群分化中心[3]。本研究尝试通过调查天台县大雷山的夏蜡梅种群结构和群落学特征,研究群落组成树种之间的相关性,反映该种群数量动态,揭示群落的结构和功能,评价该种群与生境间的适合度,并预测群落的发展动态,为境内夏蜡梅种群的保护和发展提供科学依据[10-12]。
1 研究地区概况
大雷山位于天台县南面,属仙霞岭山脉,最高峰海拔1 229.4 m,是浙东南第2高峰,也是永安溪与始丰溪分水岭。区域内年平均气温为16.9 ℃,极端最高气温41.7 ℃,极端最低气温-9.1 ℃,≥10 ℃活动积温2 858.0~5 157.0 ℃,全年无霜期243.0 d,年平均降水量1 411.7 mm,相对湿度78%,土壤pH 4.7~5.4[8,13-14]。
2 研究方法
2.1 样地设置与调查
在天台县夏蜡梅分布区域内,选取有代表性的区域进行样地调查。样地面积为20 m×20 m,均匀分成16个5 m×5 m的样方调查乔木层,后在每个样方的右下角划出2 m×2 m的小样方调查灌木层和草本层。乔木层(胸径≥1 cm)进行每木调查,记录种名、高度、胸径、冠幅等;灌木层和草本层记录种名、高度、株数及盖度,并调查样地内的所有层间植物,同时记录样地环境资料(表1)。测量样地内全部夏蜡梅的树高、胸径、冠幅、年龄、分枝数。

表1 大雷山夏蜡梅群落样地调查特征Table 1 Survey characteristics of S. chinensis community plot in Dalei Mountain
2.2 数据处理
2.2.1 重要值计算[15]乔木层重要值(VI)=(相对密度+相对显著度+相对频度)/3×100%。灌木层、草本层种的重要值(VI)=(相对密度+相对盖度+相对频度)/3×100%。

2.2.3 种群结构计算 根据夏蜡梅1个生长季内只形成1次生长高峰,且9月中旬之后就不再生长的特性[18],本研究采用数节法计算夏蜡梅的种群结构。
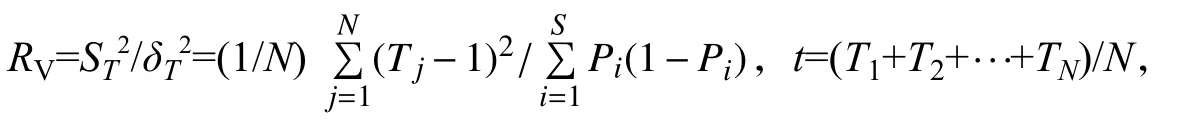
共同出现百分率:PC=a/(a+b+c)。PC的值域为[0,1],PC越接近于1,表明2个物种间正联结越紧密;若PC为0,表明该种对间无关联。联结系数:CA=2(ad-bc)/[(a+b)(b+d)+(a+c)(c+d)]。CA用来说明种间联结程度,其值域为[-1,1]。CA为0,说明2个物种间完全独立;CA越接近于1,说明2个物种间正联结越强;CA越接近于-1,说明2个物种间负连接越强。PC和CA计算公式中:a为种A和种B同时出现的样方数,b为只有种A出现的样方数,c为只有种B出现的样方数,d为种A和种B都不出现的样方数。
Pearson积矩相关系数和Spearman相关系数是反映2个物种种间协变线性关系的重要指标,可用来定量分析2个物种间的线性关系,其计算参照文献[21]。
3 结果与分析
3.1 群落结构
根据重要值计算结果,参照《中国植被》的群落命名原则[23],夏蜡梅分布区的植被类型有落叶阔叶林(Q1化香树林、Q2山胡椒林、Q3灯台树林),针叶林(Q4杉木林),竹林(Q5毛竹林)3个类型。乔木层优势种是化香树、山胡椒、灯台树、杉木、毛竹;灌木层优势种是中国绣球Hydrangea chinensis、夏蜡梅、悬铃木叶苎麻Boehmeria tricuspis;草本层优势种是金星蕨Parathelypteris glanduligera、辽宁堇菜Viola rossii、透茎冷水花Pilea pumila、虎杖Reynoutria japonica(相关重要值详见表2)。这与金则新等[9]调查时认为夏蜡梅主要分布在针阔混交林的结论略有不同,可能是夏蜡梅适应性广,适宜生境中植被类型多样导致。
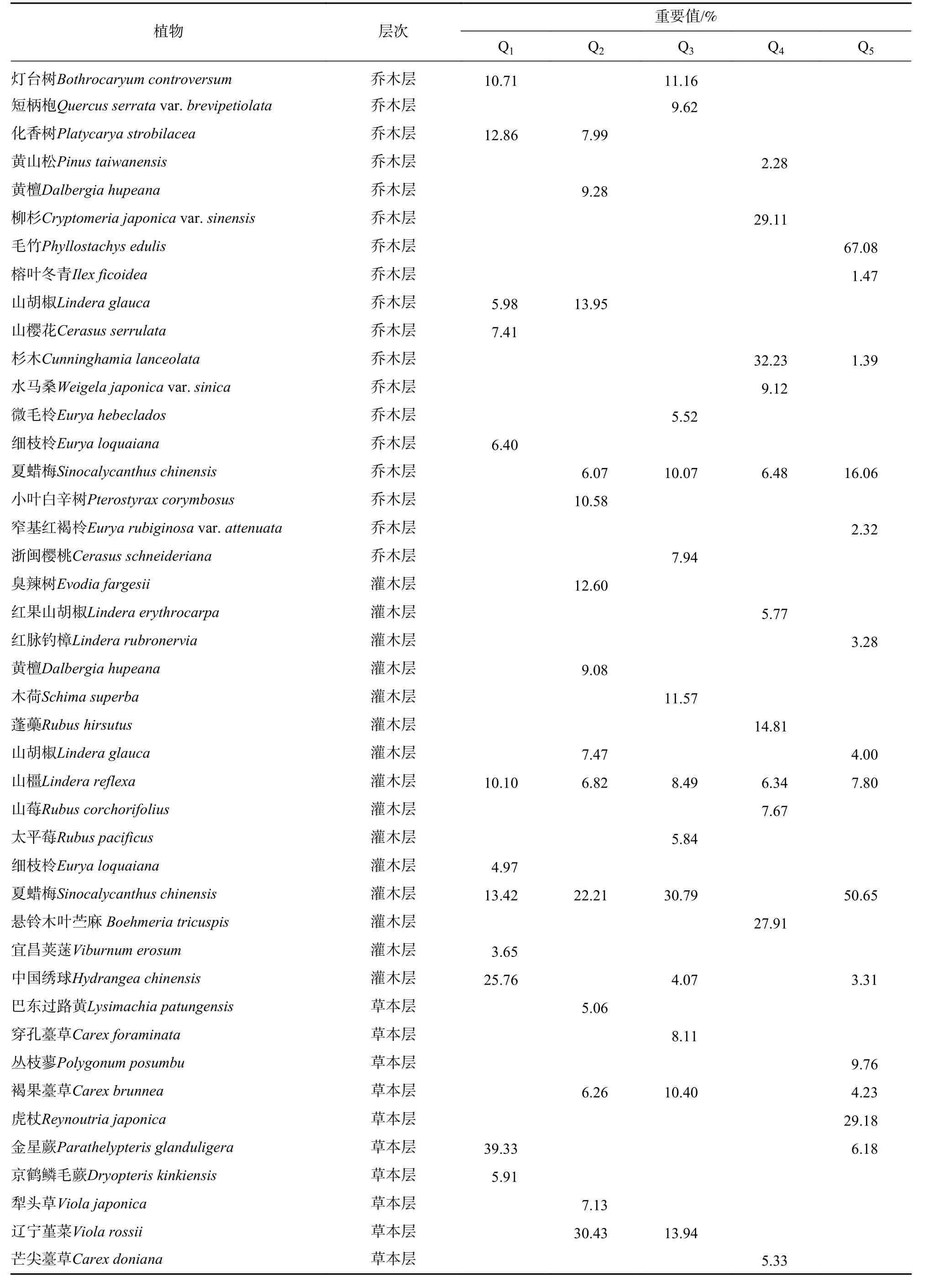
表2 大雷山夏蜡梅群落各层重要值前5的物种及重要值一览表Table 2 List of species and importance values of the top 5 important values of each layer of S. chinensis community in Dalei Mountain

表2 (续)Table 2 Continued
从重要值看,在乔木层,夏蜡梅处在伴生种位置,而在灌木层夏蜡梅常处在优势种或常见种位置,尤其是在毛竹林下,夏蜡梅重要值为50.65%,达到最高。
3.2 物种多样性
通过上述3个多样性指数来分析样地物种多样性。计算(表3~5)表明:乔木层物种多样性指数从大到小依次为阔叶林、针叶林、毛竹林,除均匀度指数外,其余指数阔叶林约是毛竹林的2倍,差异明显,说明阔叶林物种丰富、多样性明显、均匀度高。灌木层物种丰富度指数从大到小依次为阔叶林、毛竹林、针叶林,物种多样性和均匀度指数为阔叶林≈针叶林>毛竹林,说明灌木层阔叶林物种多样性指数最高,毛竹林物种丰富度较高,但均匀度低。草本层物种多样性指数除均匀度指数外,均为针叶林>阔叶林≈毛竹林,说明针叶林物种丰富度高,但均匀度低。

表3 夏蜡梅群落 5个样地乔木层物种多样性指数Table 3 Species diversity index of trees in 5 plots of S. chinensis community
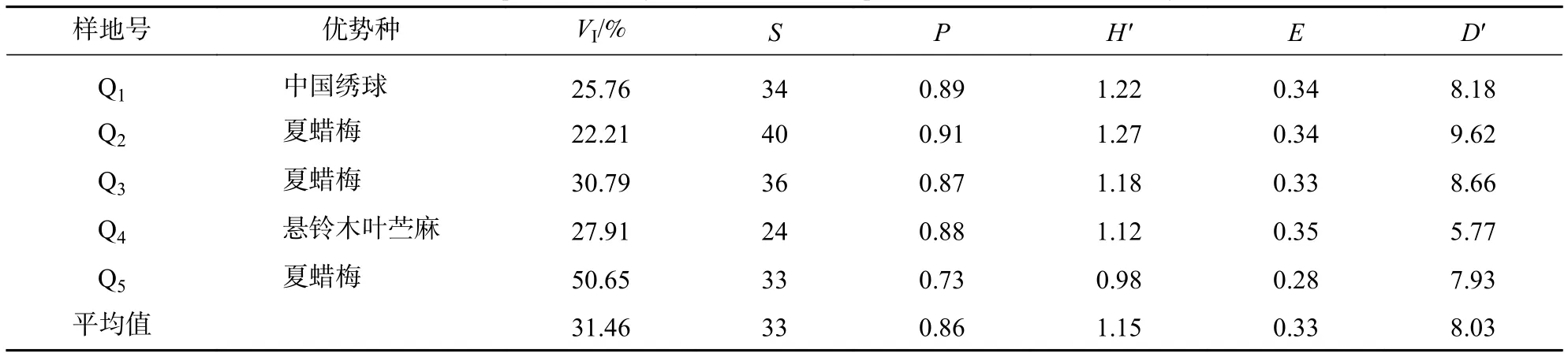
表4 夏蜡梅群落 5个样地灌木层物种多样性指数Table 4 Species diversity index of shrubs in 5 plots of S. chinensis community

表5 夏蜡梅群落 5个样地草本层物种多样性指数Table 5 Species diversity index of herbs in 5 plots of S. schinensis community
夏蜡梅在群落内的优势度与均匀度指数呈负相关,表明夏蜡梅的种间竞争较弱,是一种集群分布的物种。
3.3 种群结构
图1显示:5个样地中共有夏蜡梅453株,个体中节数最少的为1节,最多的为46节,平均16节,株数最多为1节,共177株,占39.07%。按10节为1级,共分5个级别,编制种群结构图,可见第Ⅰ级(1~10节)的个体比率最高,占56.51%,整体呈增长型中的“金字塔”型,Ⅰ级个体比较丰富,这与金则新等[9]的结论一致。说明天台大雷山的生境适宜夏蜡梅种群的生长与更新,同时也说明数节法在分析夏蜡梅的种群内部结构是可行的。
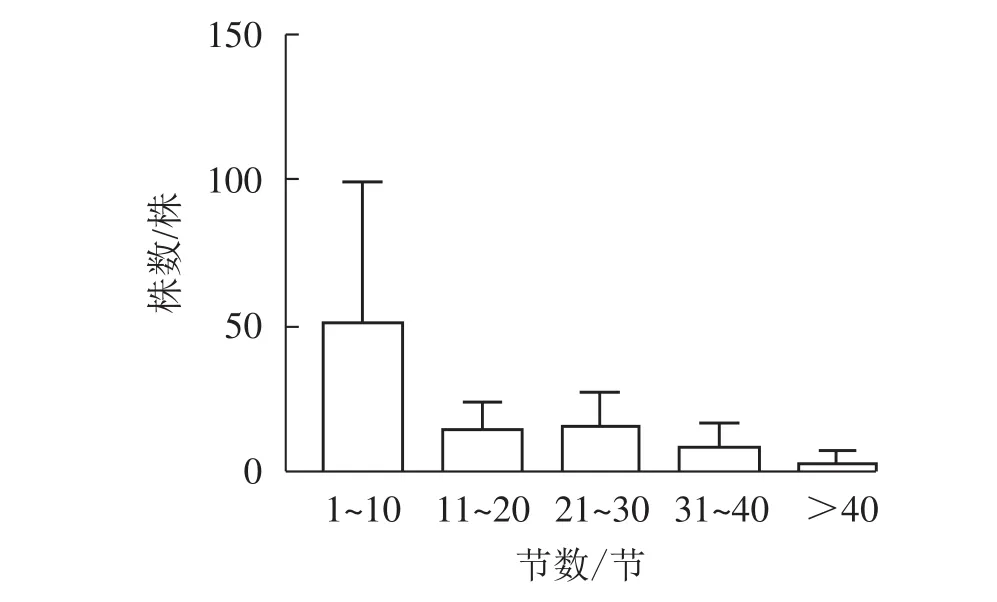
图1 天台县夏蜡梅种群结构Figure 1 Populations structure of S. chinensis in Tiantai County
由图2显示:不同的样地中,Q1、Q2、Q3都是Ⅰ级个体较多,种群结构呈“金字塔”型,属增长型;在Q4中,Ⅰ级个体极少,种群结构呈“壶”型,属衰退型;Q5中,种群结构呈“钟”型,属稳定型。
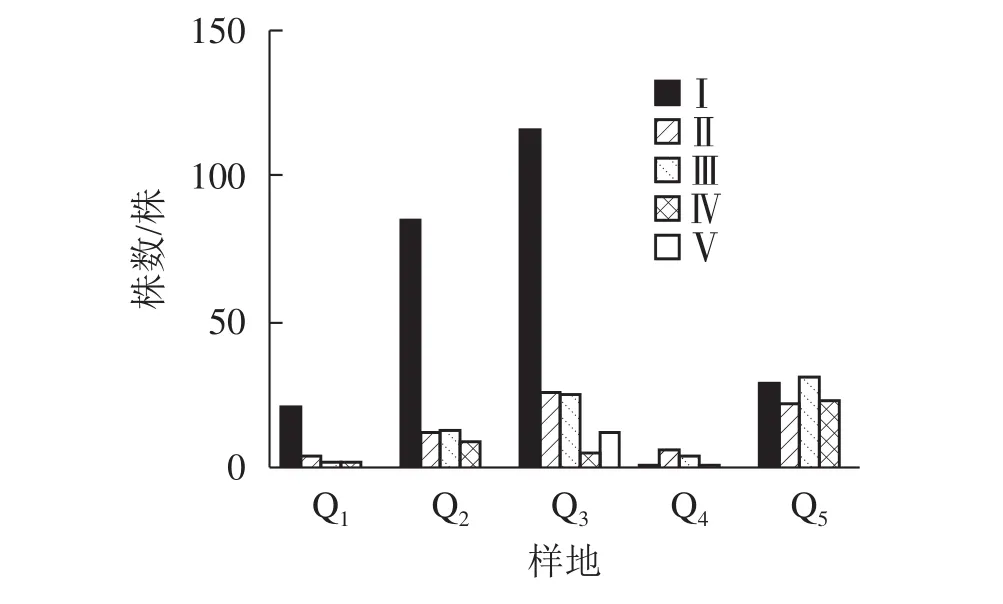
图2 天台县不同样地中夏蜡梅种群结构Figure 2 Populations structure of S. chinensis in Tiantai County
3.4 种间关联
根据夏蜡梅群落中乔木层和灌木层重要值大于5%的物种分别在5个样地中出现(1)和不出现(0)矩阵,计算出方差比率RV和统计量W。
乔木层中共选取了18个物种,RV=1.634>1,表明种间呈净的正关联;灌木层中共选取了12个物种,RV=1.250>1,表明种间呈净的正关联。由于N=5,乔木层和灌木层的统计量W分别为8.171和6.250。查表得:χ20.50,5=4.351,χ20.10,5=9.236,乔木层和灌木层的W值均落入χ20.50,5和χ20.10,5之间,即RV偏离1不显著。因此,乔木层18个物种和灌木层12个物种均在整体上表现出不显著的净的正关联。
通过共同出现百分率检测、Pearson相关系数检验、Spearman秩相关系数检验等方法分析种间联结和种间相关性,结果(图3~10)显示:乔木层和灌木层中夏蜡梅与大部分物种间均无关联,仅在乔木层中,夏蜡梅与红果山胡椒呈极显著负相关,与化香树和浙闽樱桃呈显著负相关;在灌木层中夏蜡梅与山莓呈显著负相关,与山橿间的联结性较强。
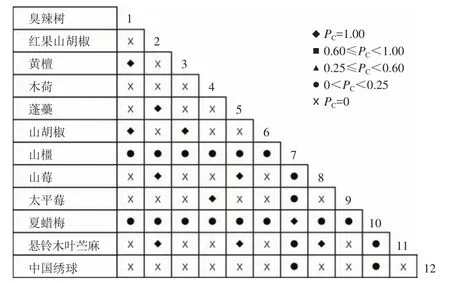
图3 灌木层共同出现百分率Figure 3 Percentage co-occurrence in shrubs layer-PC

图4 乔木层共同出现百分率Figure 4 Percentage co-occurrence in tree layer-PC

图5 灌木层联结系数Figure 5 Associations coefficient in shrub layer
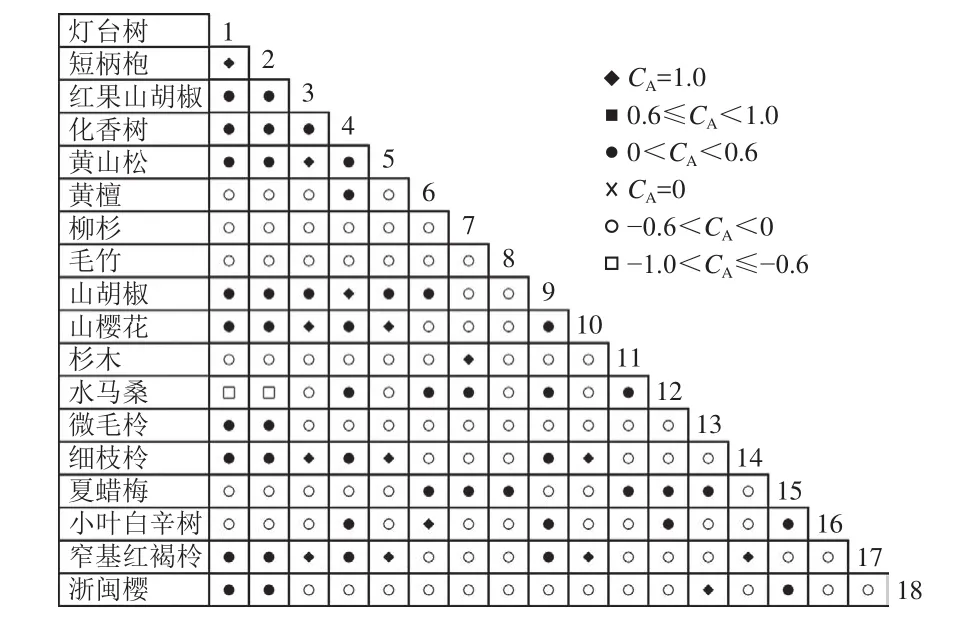
图6 乔木层联结系数Figure 6 Associations coefficient in tree layer
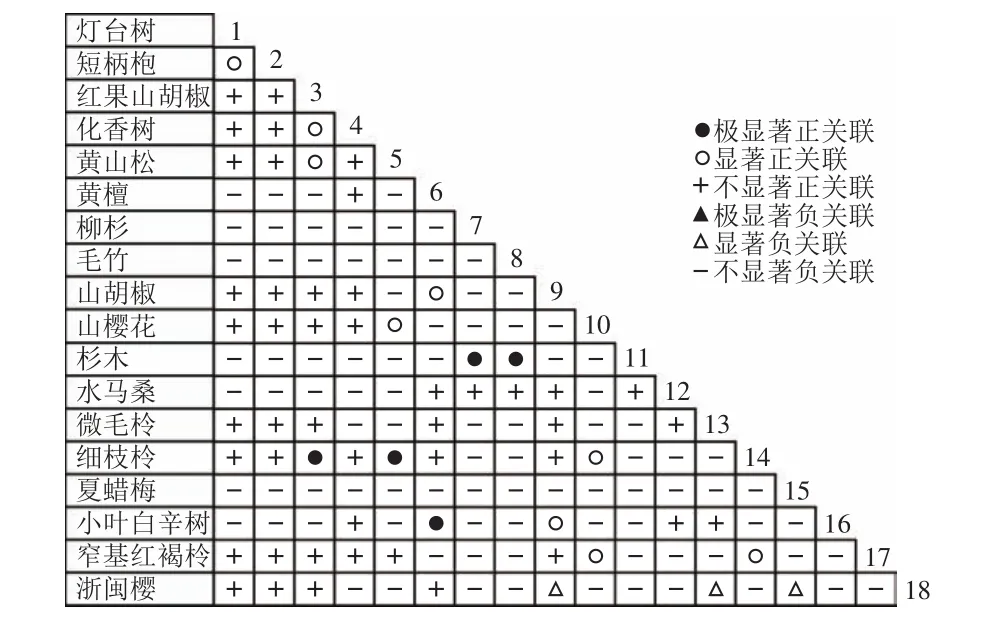
图7 乔木层Pearson系数Figure 7 Pearson coefficient in tree layer

图8 灌木层Pearson系数Figure 8 Pearson coefficient in shrub layer
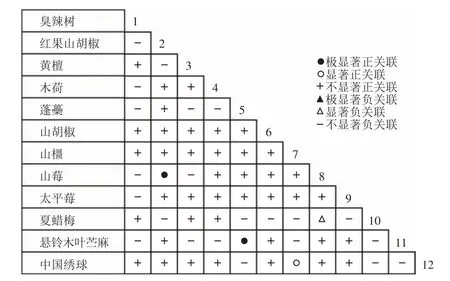
图9 灌木层Spearman系数Figure 9 Spearman coefficient in shrub layer
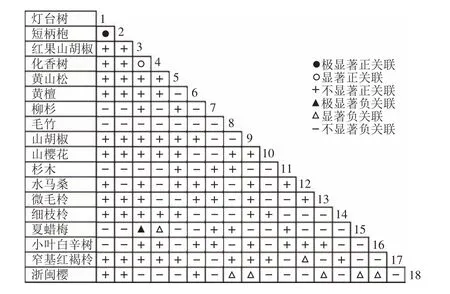
图10 乔木层Spearman系数Figure 10 Spearman coefficient in tree layer
4 结论与讨论
4.1 夏蜡梅所处群落结构分析
本研究表明:夏蜡梅所处的群落类型多样,有落叶阔叶林、针叶林、毛竹林,乔木层优势种有化香树、山胡椒、灯台树、杉木、毛竹;灌木层优势种有中国绣球、夏蜡梅、悬铃木叶苎麻;草本层优势种有金星蕨、辽宁堇菜、透茎冷水花等。夏蜡梅种群主要集中分布在落叶阔叶林和毛竹林中。生物多样性指数最高的是灯台树林,最低的是毛竹林;均匀度指数最高的是杉木林,最低的是毛竹林。
4.2 夏蜡梅种群结构分析
夏蜡梅的种群结构是增长型中的“金字塔”型,幼林个体丰富,与早期“天台县夏蜡梅产地由于植被破坏严重,夏蜡梅都呈散生状”的描述相比[6],种群得到了很好的保护。
除柳杉疏林外,落叶林、毛竹林下的夏蜡梅种群数量都比较多,特别是毛竹林下种群数量达到最高。这可能是夏蜡梅是喜荫植物,午间强光、高温会导致其叶肉细胞活性降低,进而引起光合能力的下降,直至幼苗、幼树死亡[24]。落叶林、毛竹林的林冠能为其遮光,减弱强烈的光照,降低土壤表明温度,减少土壤水分蒸发,从而促进幼苗的萌发和存活。这也为今后野外保护、繁育夏蜡梅提供了很好的指示。
4.3 种对种间联结关系研究
3种检验结果表现出一致性,正负关联比均大于1,说明各树种联系紧密,群落的结构和功能趋于完善。夏蜡梅与大部分树种无相关关系,说明夏蜡梅在群落中可能处于一个相对独立的地位。群落中与夏蜡梅呈负相关关系的主要是红果山胡椒、山莓、浙闽樱桃、化香树等树种,说明这些种与夏蜡梅有竞争关系。建议在后期夏蜡梅野外保育过程中,对与夏蜡梅有竞争关系的物种,采取适当人为择伐、抚育,营造良好的生长环境。
4.4 人为干扰对夏蜡梅种群的影响
人为干扰对夏蜡梅种群的影响不尽相同。在柳杉林等人工经营林,由于砍伐强度大,露出大量林窗,生境遭到破坏,夏蜡梅种群结构趋于衰退;在毛竹林中,虽然人工干预强烈,但对夏蜡梅的生境破坏不强,使得种群结构趋于稳定;在化香树林、山胡椒林等落叶阔叶林中,是先有人为干扰,后干扰逐渐减弱,群落进入自然演替,夏蜡梅种群则开始恢复,表现出种群增长的模型。
5 致谢
浙江农林大学赵宏波审阅全文并提出修改意见;浙江省森林资源监测中心钟建平,浙江中医药大学林王敏、董荧荧、金晓青、王志栋等参加野外调查;杭州师范大学陈伟杰帮助内业计算。在此一并致谢!
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
热带农业科学(2019年8期)2019-09-19
绿色科技(2017年15期)2017-09-01
绿色科技(2017年9期)2017-06-01
农家科技下旬刊(2017年2期)2017-03-27
中国篆刻·书画教育(2016年2期)2016-03-07
中国火炬(2009年7期)2009-07-24
中国新闻周刊(2004年30期)2004-08-23
